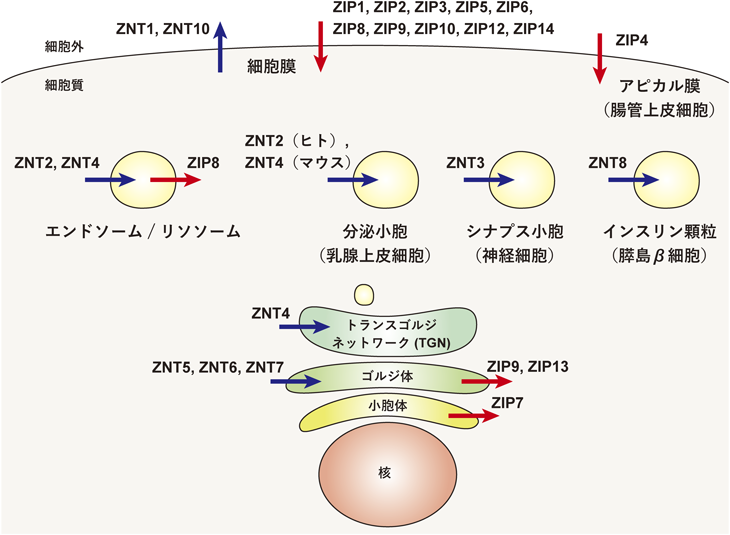
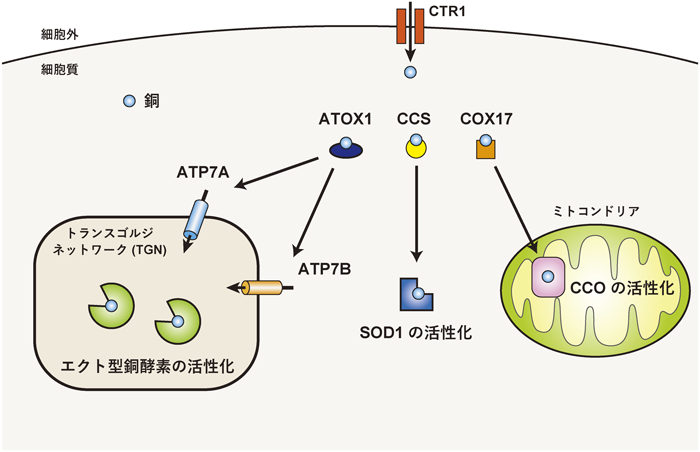
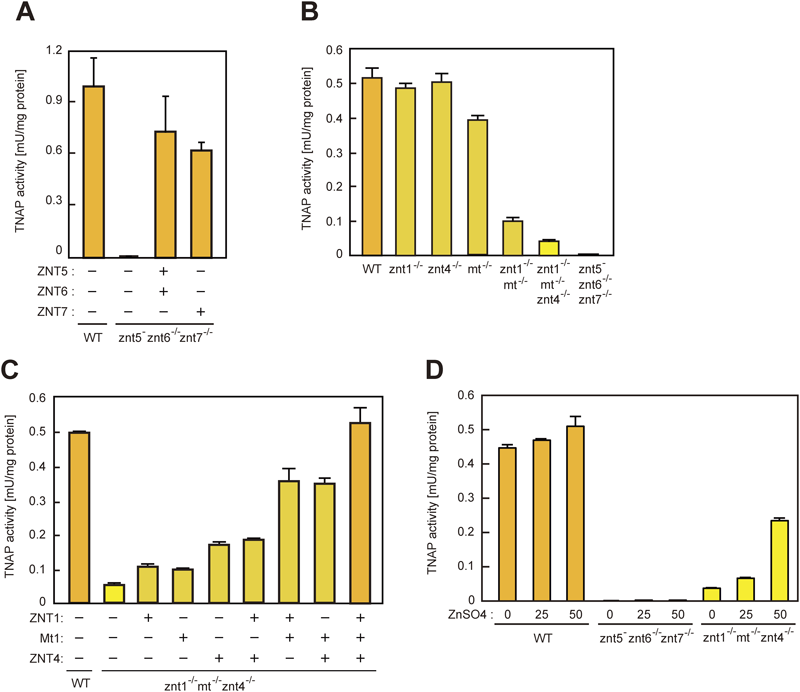
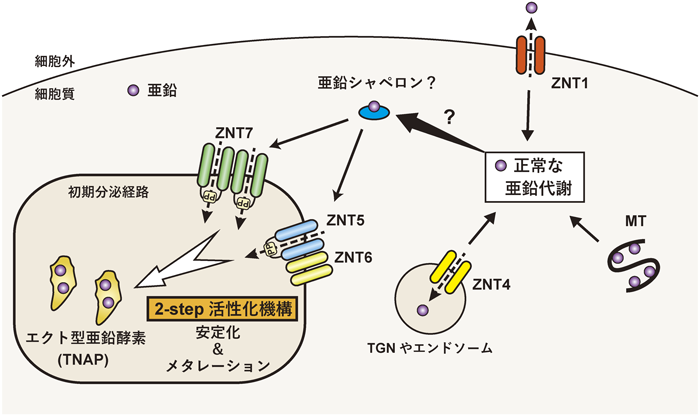
エクト型亜鉛酵素の亜鉛獲得による活性化機構Activation mechanism of zinc-requiring ectoenzymes by zinc acquisition
京都大学大学院生命科学研究科Graduate School of Biostudies, Kyoto University ◇ 〒606–8502 京都市左京区北白川追分町 ◇ Kitashirakawa-Oiwake-cho, Sakyo-ku, Kyoto 606–8502, Japan
発行日:2018年6月25日Published: June 25, 2018