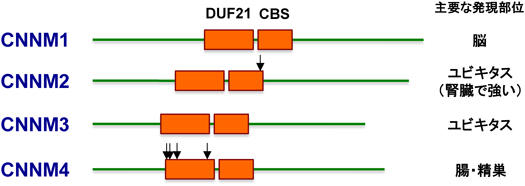
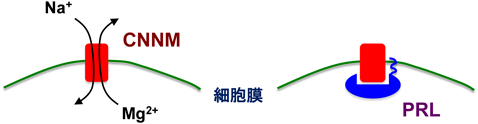
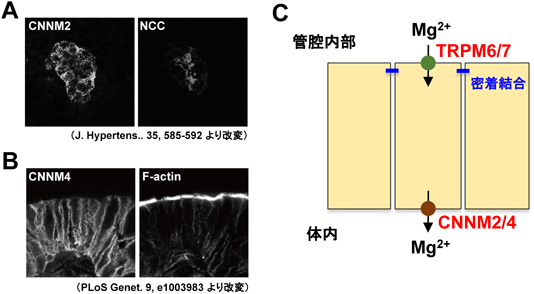
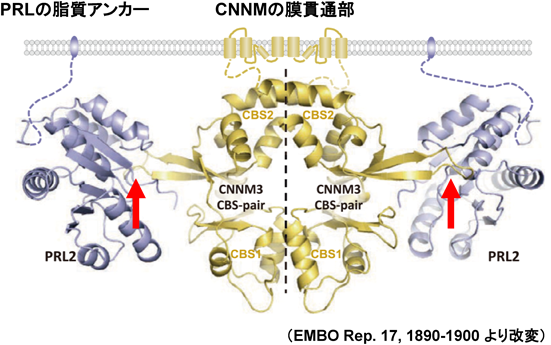
Mg2+トランスポーターCNNMの分子機能と医学生物学的重要性Molecular functions of CNNM Mg2+ transporters and their biomedical importance
大阪大学・微生物病研究所Research Institute for Microbial Diseases, Osaka University ◇ 大阪府吹田市山田丘3–1 ◇ 3–1 Yamadaoka, Suita, Osaka
発行日:2018年8月25日Published: August 25, 2018