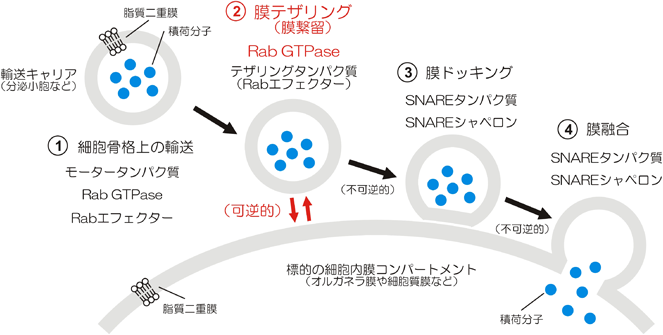
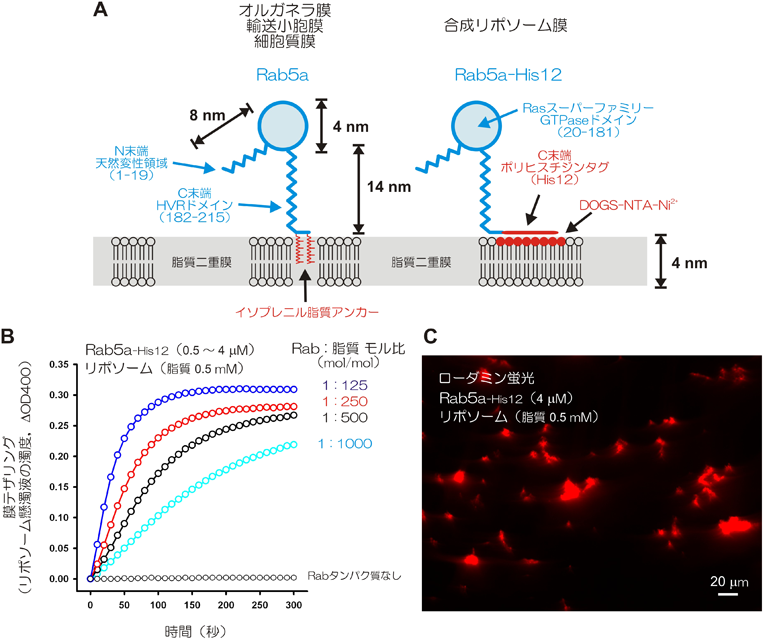
ヒトRabファミリー低分子量GTPaseが駆動する細胞内膜テザリング反応の再構成Reconstituted membrane tethering driven by human Rab-family small GTPases
大阪大学蛋白質研究所Institute for Protein Research, Osaka University ◇ 〒565–0871 大阪府吹田市山田丘3–2 ◇ 3–2 Yamadaoka, Suita, Osaka 565–0871, Japan
発行日:2018年8月25日Published: August 25, 2018