1. はじめに——タンパク質接着を担うトランスグルタミナーゼとは
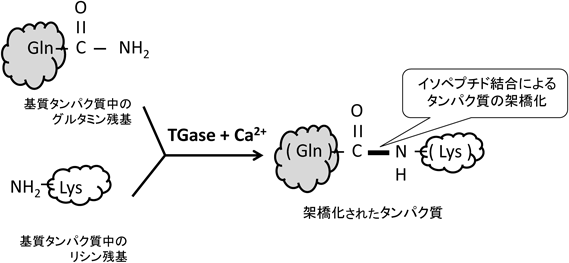
生物界にはタンパク質どうしが接着する酵素反応が存在するのをご存知だろうか.表題にも「接着」と記したがわかりよいための言い換えで,通常はタンパク質「架橋化」酵素[トランスグルタミナーゼ(transglutaminase)]による反応である.「接着」の意味合いとしては,タンパク質どうしが,共有結合レベルで不可逆的な架橋をすることである.つまりそれまでの構造が一変し,元に戻らない結合が完成される(図1).この酵素反応は微生物から植物,動物まで幅広く存在し,できあがったタンパク質は新たな機能が付与されたり,あるいは溶解性が下がったりなど,翻訳後修飾の一つとして構造に大きな変化をもたらす1, 2).酵素反応自体は今から60年前に見つかっており,生体内での生理的役割について多くの研究がなされてきた.一方,微生物(放線菌)由来の酵素標品は食品産業等でも応用されるなど,幅広くいろいろな領域に顔をのぞかせる.
高等動物では組織分布や役割を異にする8種類のアイソザイム(Factor XIII, TG1~TG7)が存在し,血液凝固,皮膚表皮の形成,細胞外マトリクスの強化などに必須である.詳細を述べると,Factor XIII(血液凝固第十三因子:以下FXIII)は,血液凝固のカスケードの最終産物としてフィブリンを架橋重合して安定な不溶物を形成させる.表皮の形成においては,TG1とTG3が主要な酵素として表皮のバリア機能の発揮に必須である.ここでの接着反応は表皮細胞内の構造タンパク質を架橋重合し,表皮分化の最終段階に細胞膜を裏打ちする高分子複合体を形成させ,これが強固なバリア機能を発揮する3).TG2は幅広くいろいろな組織に発現して,細胞死制御や細胞外マトリクスの強化,転写制御などさまざまに関与する.また架橋反応以外の機能(GTPase活性,フィブロネクチンとインテグリン結合の強化など)も有する.TG4は精巣に発現し,げっ歯類では交尾栓形成に必要とされる.TG5, TG6, TG7は表皮で発現される遺伝子群として見いだされたが,機能の詳細はまだ明らかでない1).
このようにトランスグルタミナーゼは必要な状況に応じて,できあがったタンパク質を修飾して機能変換を担っている.酵素反応はカルシウム依存性であるため,カルシウムが一つの制御因子となっている.しかしこの制御が崩れたり,異常な発現レベルになったりする場合には疾患を生体にもたらす.実際にFXIIIやTG1が活性を失った場合,血液凝固や皮膚形成において重篤な疾患が生じることが報告されている4, 5).また,TG2が小腸で過剰に働くと,コムギグルテン由来のペプチドを修飾して抗原となり,セーリアック病という自己免疫疾患を起こす6).これは欧米諸国では深刻な問題となっており,頻度も人口の1%と比較的高く,重要な疾患とされている.そのため,近年は阻害剤や活性制御剤の開発を目指した研究が競争的に行われている.それぞれのトランスグルタミナーゼの遺伝子欠損マウスは作り出されているものの,組織特異的な欠損動物,またスクリーニングを進めるうえでハイスループットな探索用の動物を用いた研究などはまだ途上である.
そのような状況の中で,我々は小動物としてのメダカ(ニホンメダカOryzias latipes)を利用できないかとの研究をこの数年行ってきた.メダカを対象にしたのは,所属する名古屋大学ではかねてよりメダカを対象にしたさまざまな研究が行われてきたこと,バイオリソースとしてメダカを対象にした研究が行われている基礎生物学研究所(岡崎市)と連携したなどの理由による.メダカは体長が成魚では約2 cmであり,繁殖が早くコスト面も含めて飼育が容易である.ゼブラフィッシュに比すとまだ国際的な知名度は低いかもしれないが,ゲノム配列やcDNAデータベースもほぼ整っていて,発生や行動の研究から,疾患モデル研究にまで広く用いられている7–9).
2. メダカのトランスグルタミナーゼ相同遺伝子(オルソログ)
そこでまず,メダカのトランスグルタミナーゼはどのようなものか,遺伝子検索から始めた.先述したように哺乳類のトランスグルタミナーゼは,ゲノム上も8個のアイソザイムが存在する.メダカについては,先述の基礎生物学研究所NBRP(National BioResource Project)によるcDNAデータベースが整い,存在が調べられ,cDNAも入手できる(ウェブサイト:https://shigen.nig.ac.jp/medaka/).
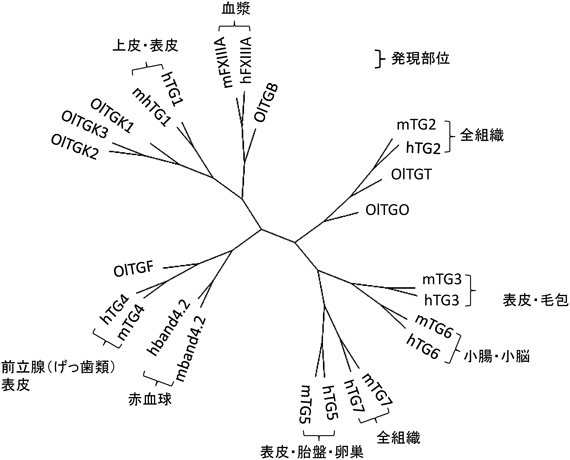
そこで,マウスの主要なアイソザイム(TG1, TG2, TG3, FXIII)のアミノ酸配列を使って相同な遺伝子をくまなく検索した.得られた配列を基に,推定アミノ酸配列をこれまでの哺乳類のものと比較したところ,いずれの遺伝子も酵素活性発現部位など機能に必要な領域はきちんと保存されており,推定分子量もほぼ同じであった.系統樹を作成して検討したところメダカには明確な7種のオルソログ(相同遺伝子)が存在した(図2).NBRPより分与されたcDNAを基に,全長であることを確認した後に,すべての組換えタンパク質を得ることができた.また,並行してゲノム編集によって遺伝子変異個体も獲得した.以下,それぞれのアイソザイムごとに特徴を述べる.
1)血液凝固に関するトランスグルタミナーゼのオルソログ
FXIIIは翻訳された直後の前駆体の段階では活性を持たない.血液凝固は「傷害」イベントで初発の反応からの何段階もの酵素反応を経て,トロンビンという鍵となる酵素が,FXIII前駆体を限定分解して活性化状態にする.これは突発的に凝固が起こらないための巧妙な生体の仕組みであり,メダカFXIIIの場合でも,ヒトのトロンビンと反応させると同様の活性化を再現できる.メダカでの他の血液凝固関連酵素はまだあまり明らかではないが,おそらく同様の血液凝固反応が生じていると推測される.またヒトFXIIIは骨・軟骨の形成にも関わることが報告されているが,不確実という反論もあった10).しかし我々はメダカのFXIII(OlTGB)の生化学的な解析や欠損変異体作製を行った結果,血小板に相当する細胞に加えて,脊椎骨をはじめいくつかの骨組織での発現を認めた.さらに,変異体での血液凝固は不完全で,本来の基質であるフィブリンの接着重合化が生じないことに加え,骨にも異常があったこと(脊椎での棘の形成)を明らかにしている11).また,FXIII前駆体を限定分解するトロンビンについても明確なオルソログがメダカに存在することを見いだしたため,発現解析や変異体作製を行って,より重症の血液凝固不全メダカモデルの確立も進めている.
2)皮膚表皮に関わるトランスグルタミナーゼのオルソログ
ヒトでは皮膚表皮に関わる2種類の主要な酵素が,細胞内タンパク質を接着して重合化させ,不溶性で安定した高分子の塊を細胞膜直下に固定する.メダカなどの魚類では,表皮組織は存在するがうろこがあるために両者がバリア機能を果たしているようである.表皮内にヒトと同様な基質や産物がどの程度あるかは不明であるが,メダカにもヒト表皮に特異的なトランスグルタミナーゼのオルソログが存在した.
哺乳類では3種の酵素(TG1, TG3およびTG5)があるが,意外なことにメダカにはTG1のみに相同な酵素が3種類存在した.このうち2種(OlTGK1, OlTGK2)の存在量が多かったため,それぞれの組換えタンパク質を作製し,酵素活性のあることを確認した.表皮に特異的に存在するかどうか,抗体を作製して調べたところ,表皮以外にも椎間板や腺組織にも存在した12).
さらにこの遺伝子欠損体を複数種得ることができたが,表皮がうまく形成されないことが予測された.実はマウスの場合,TG1欠損はバリア機能を有する表皮が完成されず誕生後数日で死に至る.ところがメダカは成魚まで異常なく生育し,形態にも特別な変化はみられなかった.そこで,仔魚と呼ばれる,孵化後数日で泳ぎ始めた段階のメダカについて,塩水(150 mM NaCl)を含む水中で生育させたところ,野生型と比べて生存時間が短い,という結果になった.これはきちんとした表皮ができずに浸透圧変化等に耐えられないためと推測しているが,成魚の場合はウロコがあるためにその効果が表れにくいのではと考えている.今後は,表皮がどのような変化をしているのか調べる予定である.また,水質検査など環境モニタリング等へ応用ができないかとも考えている13).
3)遊泳行動解析の違いを示す組織型トランスグルタミナーゼオルソログの変異個体
残る主要な酵素,哺乳類のTG2オルソログについても同様に解析を進めている.ヒトのTG2は幅広い組織に分布するともにその役割は多彩である.先に述べた酵素活性に加えて,GTPase活性,フィブロネクチンとインテグリンの結合補助のように,酵素活性とはまったく無関係な機能も併せ持つ不可思議な多機能タンパク質でもある.その一方でノックアウトしたマウスは,マクロファージの貪食能や傷害修復に異常がみられる等の報告はあるが外面的には普通の状態であった14).
そこで,メダカのオルソログ(OlTGT)の生化学的解析をしたうえで15),遺伝子変異個体を作製して解析した.その結果,一見して何の変化もなく,マウス個体や線維芽細胞株で報告のある傷害修復遅延も明確でなかった.
しかしその後,飼育する水槽の上に手をかざした際に,やや反射行動が遅いという事実が発覚した.またちょうどそのころ,基礎生物学研究所にメダカの遊泳行動を追跡する装置が導入されていた.そこでこれを利用して,5分間の遊泳距離をこの変異型と野生型とで比較した.ゲノム編集技術の詳細を省くが,我々は自らCRISPR/Cas9で変異体を確立し,また共同研究者の木下政人博士(京大農)にTALEN法によっても変異型を取得してもらっていた.2種類の変異型を用いて,5分間の遊泳距離を野生型と比較した.その結果,変異型の方が6~7割の程度ではあるが,再現性をもって遊泳距離が短く,特にこれはオスについて顕著な結果であった(投稿準備中).現在,その理由を解明すべく,脳神経でのOlTGTの発現パターンを詳細に解析するとともに,神経細胞のどの細胞種や領域にOlTGTが発現して関わるのかの解明を目指している.
4)まとめ
我々はこのようにメダカのトランスグルタミナーゼファミリーについてその性状と変異体の挙動を明らかにしてきた(表1).意外な発現分布などもあったが,変異体が示した血液凝固や表皮形成での表現型はヒトの機能からおおむね予想できるものであった.このことから,関連機能での薬剤スクリーニングのモデルとしても活用ができると考えている.この点で,メダカの場合,変異体(個体数)を取得しやすいのは利点である.また,行動異常(OlTGT変異)や骨形成の異常(OlTGB変異)などから,新たなトランスグルタミナーゼの生理機能を見いだされる可能性もある.
表1 マウスとメダカにおける酵素の役割と変異個体の形質| メダカ酵素 | ヒト酵素 | 既知の生理機能(ほ乳類) | 遺伝子変異個体の形質 |
|---|
| マウス | メダカ |
|---|
| OlTGB | Factor XIII(FXIII) | 血液凝固 | 血液凝固阻害,流産 | 血液凝固阻害,骨形成異常 |
| OlTGK1 | TGase 1(TG1) | 表皮形成 | 表皮形成異常(角化形成不全:出生後死亡) | 仔魚の段階で塩水内での生存日数の短縮 |
| OlTGK2 |
| OlTGK3 |
| OlTGT | TGase 2(TG2) | 細胞死制御や転写調節因子の不活性化など | 傷害修復遅延マクロファージの貪食能障害 | 単位時間の遊泳距離の減少 |
| OlTGO | 該当なし | — | 未解析 |
| OlTGF | 該当なし | — | 未解析 |
今後は上述の変異体の活用と,メダカでのトランスグルタミナーゼの機能解析から,ヒトでの新規機能の発見へと結びつけたい.コスト面も含め飼育の容易さと,変異体(数)の取得については多くの利点がメダカにはあり,活用に向けた展開は広がると考えている.
引用文献References
1) Eckert, R.L., Kaartinen, M.T., Nurminskaya, M., Belkin, A.M., Colak, G., Johnso, G.V., & Mehta, K. (2014) Transglutaminase regulation of cell function. Physiol. Rev., 94, 383–417.
2) Hitomi, K., Kojima, S., & Fesüs, L. (eds.). (2015) Transglutaminases, Springer Japan.
3) Candi, E., Schmidt, R., & Melino, G. (2005) The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol., 6, 328–340.
4) Koseki-Kuno, S., Yamakawa, M., Dickneite, G., & Ichinose, A. (2003) Factor XIII A subunit-deficient mice developed severe uterine bleeding events and subsequent spontaneous miscarriages. Blood, 102, 4410–4412.
5) Haneda, T., Imai, Y., Uchiyama, R., Jitsukawa, O., & Yamanishi, K. (2016) Activation of Molecular Signatures for Antimicrobial and Innate Defense Responses in Skin with Transglutaminase 1 Deficiency. PLoS ONE, 11, e0159673.
6) Sollid, L.M. (2002) Coeliac disease. Nat. Rev. Immunol., 2, 647–655.
7) Wittbrodt, J., Shima, A., & Schatle, M. (2002) Medaka—a model organism from the far East. Medaka. Nat. Rev. Genet., 3, 53–64.
8) Kinoshita, M., Murata, K., Naruse, K., & Tanaka, M. (2009) Medaka: Biology, Management, and Experimental Protocols. Ames, Wiley-Blackwell.
9) 若松鷹司(2006)メダカ学全書,大学教育出版.
10) Piecy-Kotb, S.A. & Mousa, A. (2012) Factor XIIIA transglutaminase expression and secretion by osteoblasts is regulated by extracellular matrix collagen and the MAP kinase signaling pathway. J. Cell. Physiol., 227, 2936–2346.
11) Horimizu, R., Ogawa, R., Watanabe, Y., Tatsukawa, H., Kinoshita, M., Hashimoto, H., & Hitomi, K. (2017) Biochemical characterization of medaka (Oryzias latipes) orthologue for mammalian Factor XIII and establishment of gene-edited mutant. FEBS J., 284, 2843–2855.
12) Kikuta, A., Furukawa, E., Ogawa, R., Suganuma, N., Saitoh, M., Nishimaki, T., Katsumura, T., Oota, H., Kawamoto, T., Tatsukawa, H., Hashimoto, H., & Hitomi, K. (2015) Biochemical characterization of medaka (Oryzias latipes) transglutaminases. PLoS ONE, 10, e0144194.
13) Watanabe, F., Furukawa, E., Tatsukawa, H., Taniguchi, Y., Kamei, Y., Hashimoto, H., & Hitomi, K. (2018) Biosci. Biochem. Biotechnol, 82, 1165–1168.
14) Iismaa, S.E., Mearns, B.M., Lorand, L., & Graham, R.M. (2009) Transgltuaminases nad diseases: Lessons from genetically engineered mouse models and inherited disorders. Physiol. Rev., 89, 991–1023.
15) Takada, Y., Watanabe, Y., Okuya, K., Tatsukawa, H., Hashimoto, H., & Hitomi, K. (2017) Biochemical characterization of the medaka (Oryzias latipes) orthologue for mammalian tissue-type transglutaminase (TG2). Biosci. Biochem. Biotechnol., 81, 469–474.