日本酒酵母の染色体異数性が生化学的代謝に及ぼす影響の解明と異数性に着目した育種技術の開発Metabolic impact of chromosomal aneuploidy in sake yeast
佐賀大学農学部Saga University ◇ 〒840–8502 佐賀市本庄町1 ◇ 1 Honjo-cho, Saga city, Saga 840–8502, Japan
発行日:2018年8月25日Published: August 25, 2018
© 2018 公益社団法人日本生化学会© 2018 The Japanese Biochemical Society
自然界の出芽酵母Saccharomyces cerevisiaeは二倍体として通常は生息している.すなわち,栄養条件の良好な環境では16本の染色体を2本ずつ持って増殖している.しかし,危機的な環境になると全滅を防ぎ一部の子孫だけでも生き残らせるべく,遺伝的多様性を増すために胞子形成して各染色体をシャッフリングし,多様な遺伝子を持った一倍体(16本の染色体を1本ずつの状態)を生み出す生物である.一方,日本酒酵母をはじめとする醸造酵母はもともと野生のS. cerevisiaeであったものが,栄養の豊富な醸造環境で植え継がれてきたものである.醸造環境で長年植え継がれてきた日本酒酵母は,危機的な環境を長年経験していないため,危機に対処するという意義のある胞子形成能力を遺伝的に失っており,一倍体にならずにずっと二倍体の状態だけで出芽・分裂を行うようになっている1).さらに近年のゲノム解析で,人類に家畜化されてきた多くの醸造酵母では染色体の異数性,すなわち染色体を完全に二倍体なら2本ずつ,四倍体なら4本ずつではなく余分な染色体を持つ現象が高頻度で観察されることが明らかになりつつあった2).しかしそのような異数染色体の醸造における意義は明らかになってこなかった.
これまで日本酒酵母の育種は一塩基多型(SNP)レベルの点突然変異3)や染色体の非相互転座4)によってもたらされるものと考えられてきたが,異数染色体が醸造特性に与える影響を明らかにすれば,日本酒酵母で異数染色体に着目した育種が実現できるのではないかと考え,研究を開始した.
日本酒の低アルコール製造技術はまだ発展途上の技術である.日本酒は現在の製造技術では,多くの場合でアルコール度数が最低でも17%以上になるまで醸造を行う.この技術が確立したのは糖源の追加投入を可能にする,三段仕込みに関する最古の記述のみられる多聞院日記が完成した室町時代後期以降であると考えられるが,保存貯蔵技術のなかった当時はできた日本酒を腐らせないためにこのような技術を用いる必然性があった.保存貯蔵技術の発達した現代においては,国民福祉,消費者の健康志向もあり,低アルコール製造技術が求められているといえる.しかし大規模産業化に遺伝的に適応し数百年経った日本酒酵母は,高アルコール製造技術に最適化されており5),低アルコール日本酒を造るとオフフレーバーであるジアセチルが生成するという問題があった.
酒類醸造時のジアセチルの生成機構についてはこれまで多くのことが明らかになっている6).酒類の醸造において,グルコースからピルビン酸までは解糖系でできるため,酵母の活性の高い時期にはピルビン酸が酵母細胞内から自然拡散により漏出し,発酵液のピルビン酸濃度が高くなる.ピルビン酸濃度が高い状態で上槽,すなわち発酵液を酵母から切り離してしまうと,ピルビン酸やそこからできるα-アセト乳酸が発酵液に残存し,オフフレーバーであるジアセチルになってしまう.しかし醸造期間を長くすることによりピルビン酸は酵母に再吸収されて発酵液からなくなっていき,上記のような問題は起きにくくなる.逆に醸造の途中で上槽するような低アルコール日本酒の場合,ピルビン酸濃度が高いまま酵母と切り離すことになり,ジアセチルが発生しやすいという問題があった.したがって,醸造の途中でもピルビン酸濃度が低く推移する酵母が求められており,ピルビン酸アナログ物質耐性などの手法で育種が行われていた7).
この技術課題を解決するために,我々は,ミトコンドリア輸送を増強することを意図した低ピルビン酸日本酒酵母の新たな育種手法を考案した.すなわち,遺伝子組換え技術を使わずにピルビン酸のミトコンドリアへの輸送を増強するため,ピルビン酸のミトコンドリアへの輸送の阻害剤ethyl α-transcyanocinnamateへの耐性株(TCR株)を日本酒酵母より取得した8).ethyl α-transcyanocinnamateが存在すると,酵母はピルビン酸をミトコンドリアに取り込むことができず,死滅すると考えられ,ethyl α-transcyanocinnamateに耐性を示す株は,ピルビン酸のミトコンドリアへの輸送活性が増強されている遺伝子を持っていると考えられる.実際にethyl α-transcyanocinnamateを入れた培地ではほとんどの日本酒酵母は死滅し,わずかな割合のコロニーだけが生育することができた.これらのコロニーを拾い上げ,TCR株として日本酒醸造試験に供しピルビン酸濃度を測定した.その中でもTCR7株が,研究室レベルでは低いピルビン酸濃度を示した9).TCR株は実生産レベルでも親株よりピルビン酸の生産性が低かった10).こうして育種した日本酒酵母は実際の製造現場で使われ,権威あるコンクールで受賞するなど,市場での高い評価を得ている.
TCR7株が本当にピルビン酸のミトコンドリアへの輸送活性が高いのかを調べるため,そのミトコンドリア呼吸活性を調べた.その結果,ピルビン酸を基質としたミトコンドリアの呼吸活性は,TCR7株の方が高いことがわかった.TCR7株の代謝を調べるため,13C代謝フラックス解析を行ったところ,確かにTCR7株はミトコンドリア内の代謝フラックスが高いことがわかった11).以上の解析から,TCR7株はミトコンドリア代謝活性が高いためにピルビン酸が低いと考えられた.
以上の研究結果から,TCR7株はミトコンドリア代謝活性が上がっていることがわかった.しかしその遺伝学的基盤はわからないままであった.事実,近年同定されたピルビン酸のミトコンドリアへの輸送遺伝子の発現も変わっていなかった.そこで,このTCR7株のピルビン酸低生産性の遺伝学的基盤を探るため,清酒酵母協会7号(K7-4)のゲノム情報をレファレンスにTCR7株の次世代シークエンサーを使ったゲノム解析を行ったところ,TCR7株は染色体11番と14番が通常の2本から3本になっていることがわかった(図1).
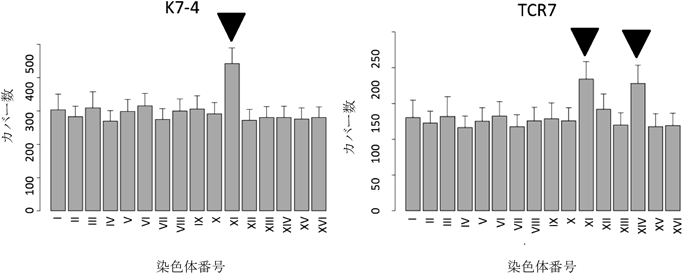
日本酒酵母(K7-4)とそのピルビン酸低生産株(TCR7)のゲノムをIllumina Miseqで解析した.染色体ごとのリード数を表し,エラーバーは標準偏差を表す.
このことから,染色体11番と14番の2本から3本への増加がピルビン酸の低減をもたらしているのではないかとの仮説を立てた.しかし,TCR7株は親株とはSNPをはじめとした多くの遺伝子変異があり,ピルビン酸の減少は遺伝子変異によってもたらされた可能性もあり,染色体が増えた株のピルビン酸が減少しているからといって単純に異数染色体が増加したためであると推定することはできない.そこで,「異数染色体の増加がピルビン酸を低減させている」という仮説を立証するため,染色体数がばらついた多数の日本酒酵母を分離するという戦略を立てた.日本酒酵母は長年,醸造環境で植え継がれてきたため,胞子形成に異常があり,一倍体を取得しても異数染色体を持つ一倍体が多発する.このような染色体数のばらついた多数の日本酒酵母を分離すれば,個別の遺伝子変異はピルビン酸を高くするものも低くするものもあるだろうから,平均すれば個別の遺伝子変異の影響はキャンセルされて「ない」ものと扱うことができる.すなわち,染色体数がばらついた多くの日本酒酵母一倍体を取得してそのピルビン酸濃度を解析すれば,個別の遺伝子変異はないものとして,純粋に異数染色体の増加のピルビン酸への影響だけを知ることができる.この考え方を用いて,日本酒酵母から多数の一倍体株を取得した.これらを使って,異数染色体の増加がピルビン酸にどのような影響を与えるかを調査した.その結果,異数染色体を持った一倍体日本酒酵母において,異数染色体の増加数はピルビン酸の濃度と統計的に有意に逆相関の関係にあった(図2).
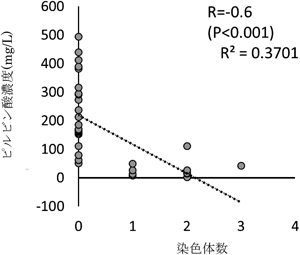
取得した日本酒酵母一倍体で日本酒を仕込みそのピルビン酸濃度を測定した.統計的有意差の計算には無相関検定を用いた.
特に11番染色体にプラスして異数染色体が増えると,ピルビン酸濃度が減少することが明らかとなった(図3).
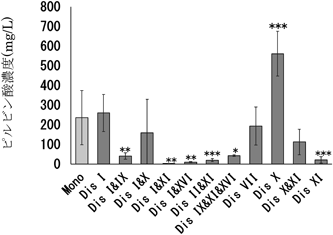
日本酒酵母一倍体のピルビン酸生産性を染色体ごとに集計した.統計的有意差の検定にはDunnet検定を用いた.***: p<0.001, **: p<0.01, *: p<0.05.Monoは異数染色体のない株を,Disは異数染色体があり染色体数が2本になっている株を,その後の数字は増えた染色体の番号を表す.
上記のことから異数染色体を持った日本酒酵母はピルビン酸を低減させる性質を持つことがわかった.以前の研究から,ピルビン酸低減日本酒酵母はミトコンドリア代謝活性が高いことがわかっていた.これらのことから,異数染色体を持った日本酒酵母はミトコンドリア代謝活性が高いためにピルビン酸を低減させていると考えられた.この仮説を実証するため,異数染色体を持った日本酒酵母一倍体株のメタボローム解析,主成分分析を行った.その結果,異数染色体の数とミトコンドリアで起きる代謝物質とが逆相関の関係にあることがわかった.またピルビン酸が低減した11番染色体が増加した株からミトコンドリア代謝活性の下がった呼吸欠損株を取得すると,ピルビン酸濃度が上がってもとに戻ることもわかった.以上のことから,11番染色体の増加が日本酒酵母のミトコンドリア代謝を活性化し,ピルビン酸を減少させていると考えられた(図4)12).
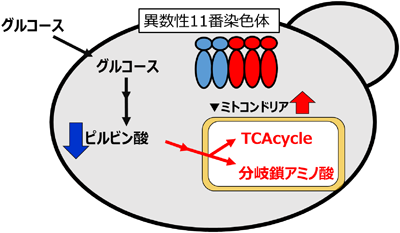
以上の結果は,11番染色体の増加自体がミトコンドリア代謝の活性化,ピルビン酸の減少を引き起こしているのか,11番染色体上の遺伝子の発現増加がこの性質をもたらしているかのどちらかによって引き起こされたと考えられる.この可能性を特定するため,11番染色体上にあるミトコンドリア遺伝子のいくつかをプラスミド上で発現させ,ピルビン酸の増減を調べた.その結果,11番染色体上にあるミトコンドリアシトクロムcペルオキシダーゼをコードする遺伝子CCP1の高発現が,ピルビン酸濃度の減少を引き起こすことを明らかにした.このことから,TCR7株ではCCP1が2本から3本になったためにミトコンドリア活性が増加し,ピルビン酸が減少したと考えられた.
以上の結果は,特定の異数染色体が増えた日本酒酵母を選択すれば,特定の遺伝子の発現を増加させることができ,特定の醸造形質を持った日本酒酵母を育種できることを示している.
これまで日本酒酵母の遺伝子組換え技術が開発されてきたが,社会的アクセプタンスの問題からその普及は遅れていた.一方,異数染色体の倍加はこれまで日本酒酵母の維持の歴史の中で何度も起きてきた現象である(図5)ことから,食経験も豊かであり,普及にも問題はないと考えられる.日本酒酵母の染色体数は不安定であり,容易に染色体数のばらついた株は取得できる.今後,遺伝子組換え技術を使わずに特定の醸造形質を制御できる日本酒酵母の育種として,異数染色体に着目した日本酒酵母の育種は普及していくと考えられる.

本研究は多くの指導者,共同研究者に支えられて行えたものである.佐賀大学大学院生の門脇真史君(現・三和酒類(株)),田口誠我君(現・霧氷酒造(株)),藤丸裕貴君,山本裕貴君,堀江健太君(現・大関(株)),平田みよさん(現・大関(株)),徳永直也君(現・オエノンホールディングス(株)),佐々木真君(現・(株)フランソア),城間翔大君(現・(株)不二家),国立遺伝学研究所の野口英樹教授,藤山秋佐夫教授,豊田敦教授,独立行政法人酒類総合研究所の赤尾健博士,佐賀大学の穴井豊昭教授,木村佑太研究員,寺沢洋平研究員,佐賀県工業技術センターの柘植圭介特別研究員,澤田和敬副主査,イタリア・バリ大学のAgrimi教授,佐賀県天山酒造の七田謙介社長,後藤潤杜氏,蔵人の皆様,元・佐賀県工業技術センターの小金丸和義先生,大阪大学大学院の福崎英一郎教授に心から感謝申し上げる.
1) Nakazawa, N., Niijima, S., Tanaka, Y., & Ito, T. (2012) Immunosuppressive drug rapamycin restores sporulation competence in industrial yeasts. J. Biosci. Bioeng., 113, 491–495.
2) Gallone, B., Steensels, J., Prahl, T., Soriaga, L., Saels, V., Herrera-Malaver, B., Merlevede, A., Roncoroni, M., Voordeckers, K., Miraglia, L., et al. (2016) Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell, 166, 1397–1410.
3) Kitagaki, H. & Kitamoto, K. (2013) Breeding research on sake yeasts in Japan:history, recent technological advances, and future perspectives. Annu. Rev. Food Sci. Technol., 4, 215–235.
4) Watanabe, D., Wu, H., Noguchi, C., Zhou, Y., Akao, T., & Shimoi, H. (2011) Enhancement of the initial rate of ethanol fermentation due to dysfunction of yeast stress response components Msn2p and/or Msn4p. Appl. Environ. Microbiol., 77, 934–941.
5) Miyashita, K., Sakamoto, K., Kitagaki, H., Iwashita, K., Ito, K., & Shimoi, H. (2004) Cloning and analysis of the AWA1 gene of a nonfoaming mutant of a sake yeast. J. Biosci. Bioeng., 97, 14–18.
6) Inoue, T. & Yamamoto, Y. (1969) Absence of diacetyl in fermenting wort. Arch. Biochem. Biophys., 135, 454–457.
7) 福田和郎,宮本ともこ,籾谷亘慶,大内弘造(1998)アルコール飲料および発酵調味料の製造方法,特開平10-179131.
8) Horie, K., Oba, T., Motomura, S., Isogai, A., Yoshimura, T., Tsuge, K., Koganemaru, K., Kobayashi, G., & Kitagaki, H. (2010) Breeding of a low pyruvate-producing sake yeast by isolation of a mutant resistant to ethyl alpha-transcyanocinnamate, an inhibitor of mitochondrial pyruvate transport. Biosci. Biotechnol. Biochem., 74, 843–847.
9) 平田みよ,元村沙織,佐々木真,堀江健太,大場孝宏,柘植圭介,吉村臣史,小金丸和義,北垣浩志(2011)ピルビン酸のミトコンドリア輸送阻害剤耐性酵母の小規模および中規模パイロットスケールでの低アルコール清酒醸造特性,日本醸造学会誌,106, 323–331.
10) 佐々木真,大場孝宏,末永光,稲橋正明,佐藤真佐恵,鶴田裕美,小林元太,柘植圭介,吉村臣史,小金丸和義,他(2011)吟醸酒製造用清酒酵母からのピルビン酸低生産株の育種と実製造でのピルビン酸およびα-アセト乳酸の低減,生物工学会誌,89, 222–227.
11) Agrimi, G., Mena, M.C., Izumi, K., Pisano, I., Germinario, L., Fukuzaki, H., Palmieri, L., Blank, L.M., & Kitagaki, H. (2014) Improved sake metabolic profile during fermentation due to increased mitochondrial pyruvate dissimilation. FEMS Yeast Res., 14, 249–260.
12) Kadowaki, M., Fujimaru, Y., Taguchi, S., Ferdouse, J., Sawada, K., Kimura, Y., Terasawa, Y., Agrimi, G., Anai, T., Noguchi, H., et al. (2017) Chromosomal aneuploidy improves the brewing characteristics of sake yeast. Appl. Environ. Microbiol., 83, e01620–e17.

This page was created on 2018-06-26T13:38:17.559+09:00
This page was last modified on 2018-08-17T09:38:09.217+09:00
このサイトは(株)国際文献社によって運用されています。