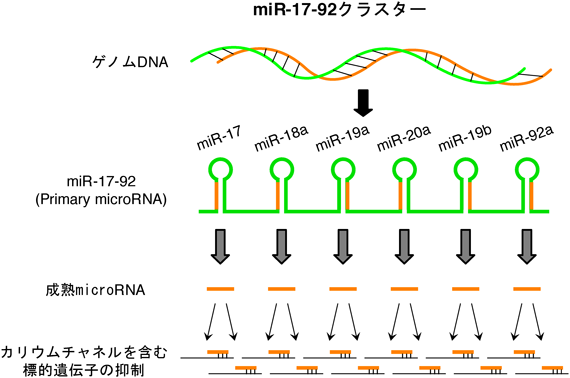
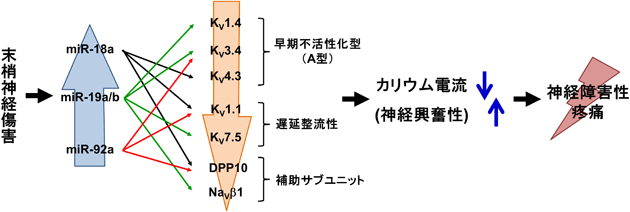
microRNAクラスターmiR-17-92による神経障害性疼痛の制御MicroRNA cluster miR-17-92 modulates the neuropathic pain
日本医科大学・薬理学Department of Pharmacology, Nippon Medical School ◇ 〒113–8602 東京都文京区千駄木1–1–5 ◇ 1–1–5 Sendagi, Bunkyo-ku, Tokyo 113–8602, Japan
発行日:2018年8月25日Published: August 25, 2018