レスベラトロール研究の進展Progress in the study on resveratrol
奈良女子大学研究院生活環境科学系食物栄養学領域Department of Food Science and Nutrition, Nara Women’s University ◇ 〒630–8506 奈良市北魚屋西町 ◇ Kita-uoya Nishi-machi, Nara 630–8506, Japan
発行日:2018年8月25日Published: August 25, 2018
© 2018 公益社団法人日本生化学会© 2018 The Japanese Biochemical Society
医学の進歩と食生活の改善によって,日本は世界有数の長寿国となっている.一方で,医療費の増大,ライフスタイルの変化による生活習慣病罹患者の増加などが問題になっている.このような社会的背景から,毎日の食生活を通して健康維持に努めることは,健康長寿社会の実現のために重要である.食品機能成分の生活習慣病予防効果が注目され,さまざまな効果が報告されている.しかしながら,その効果の分子作用機構は必ずしも明らかではない.その理由の一つに,食品機能成分が薬剤に比べて作用が弱いことが考えられる.このことは副作用が少ないという長所となる一方で,効果が現れるまでに長い時間を必要とし,科学的検証を困難にしている.我々は,食品機能成分が薬剤と同じ標的タンパク質に作用して効果を示し,薬剤よりは弱いものの長期間摂取することで効果を発揮すると考えて研究を進めている.本稿では,さまざまな生理作用を有するレスベラトロール(3,5,4′-trihydroxystilbene)について紹介する.
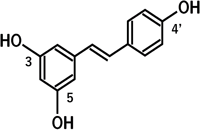
レスベラトロール(図1)は,ブドウの果皮や赤ワイン,ピーナッツ等に含まれる抗菌性物質で,1940年に高岡道夫博士(北海道帝国大学)がバイケイソウの根から分離精製,構造決定し,レスベラトロールと命名した日本発の物質である.その後1963年には,生薬の虎杖根(イタドリの根)から分離・精製されている.
レスベラトロールは,哺乳類においてSirtuinファミリーのSIRT1を活性化し寿命を延長することが報告され1),世界的に注目を集めるようになった.SirtuinはNAD+依存性ヒストン脱アセチル化酵素活性を有し,酵母,線虫,ショウジョウバエからヒトまで広く分布している.酵母から最初に同定されたSir2は,酵母の寿命制御に関わることが示されている.ヒトSirtuinには7種類のサブタイプ(SIRT1~7)が存在し,SIRT1とSir2は高い相同性をもつ.一方で,摂取カロリーの制限と抗老化作用・寿命延長との関係が注目されている.SIRT1はカロリー制限によって活性化され,ヒストンの脱アセチル化によりエネルギー代謝に関わる遺伝子の発現を調節し,細胞内のエネルギー恒常性を維持している.レスベラトロールはSITR1を活性化することによってカロリー制限の効果を模倣し,寿命延長に関わると考えられている.
さらにSIRT1活性化は,抗肥満やインスリン抵抗性の改善などのレスベラトロールのさまざまな効果に対して関与すると考えられているが,レスベラトロールが直接SIRT1を活性化するかは議論がなされており,SIRT1以外の分子作用機構が寄与する可能性が考えられる.また新しい標的として,レスベラトロールによるcAMP依存性ホスホジエステラーゼ(PDE)活性阻害が報告されている2).
我々は,レスベラトロールがある種の培養がん細胞において,誘導型シクロオキシゲナーゼ(COX-2)の酵素活性と発現の両方を抑制することを明らかにした3).さらに,レスベラトロールは細胞選択的にCOX-2発現を抑制すること,この細胞選択的発現調節に核内受容体peroxisome proliferator-activated receptor(PPAR)γ活性化が関与することを報告した4).COXは,プロスタグランジン(PG)産生の律速酵素であり,アラキドン酸を基質としてPGH2を生成する反応を触媒する.PGH2からは,合成酵素の違いによって作用の異なるプロスタノイドが産生され,選択的な受容体を介して効果を発揮する(図2).また,プロスタノイドの一部は,PPARを介して作用すると考えられている.アスピリンをはじめとした非ステロイド性抗炎症剤は,COX活性を阻害することによって抗炎症作用を持つ.COXには,酵素化学的に同定されたハウスキーピング型のCOX-1と分子生物学的な方法で同定された誘導型のCOX-2の2種類のアイソザイムが存在する.COX-2は炎症性刺激により誘導され,抗炎症性ステロイドにより抑制されることから,炎症との関与が明らかになっているが,炎症以外にも発がん,生活習慣病にも関与することがわかってきている5).
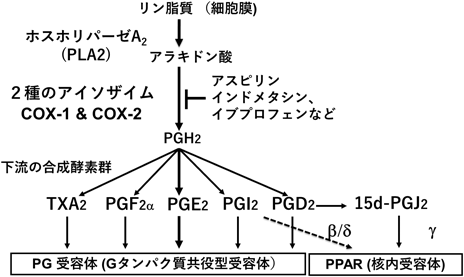
リン脂質の2位にはアラキドン酸が配位しており,これをPLA2が切り出す.アラキドン酸からCOXの触媒により生成するPGH2からは,多彩な生理作用を持つプロスタノイドが産生される.たとえばプロスタサイクリン(PGI2)とトロンボキサンA2(TXA2)は,血管の拡張と収縮,血小板凝集の抑制と促進といった相反する活性を持ち,そのバランスによって血管のホメオスタシスを維持する.
PPARは核内受容体スーパーファミリーに属するリガンド依存性転写因子で,3つのサブタイプα, β/δ, γが存在している.いずれも脂質代謝,糖代謝,細胞増殖や分化に関与している.αは主に肝臓に発現し脂肪燃焼に,β/δは筋肉などさまざまな組織に発現して脂肪燃焼や運動機能改善に,γは白色脂肪組織やマクロファージに発現してインスリン感受性に関与している.αの合成リガンドであるフェノフィブラートは高脂血症改善薬,γの合成リガンドであるチアゾリジン誘導体はインスリン抵抗性改善薬として各々処方されている6).また,多価不飽和脂肪酸をはじめとした脂肪酸や,アラキドン酸由来エイコサノイドがPPARの内因性リガンドとして作用することが明らかになっている.
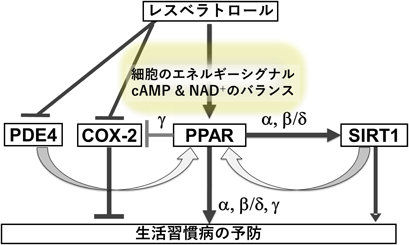
我々はマクロファージ系の細胞で,PPARγを介してCOX-2発現がフィードバック制御されることを報告した4).この制御は,PDG2の代謝産物である15d-PGJ2がPPARγのリガンドとして作用し,それがNF-κB等を介してCOX-2の発現を抑制することによる.一方で,PPARγの発現が低い血管内皮細胞ではこのようなフィードバック制御は認められず,細胞特異性があることがわかった.血管内皮細胞ではPPARγ発現ベクターを導入することでCOX-2の発現抑制効果が観察されたことから,COX-2発現抑制とPPAR活性化は相互に作用する関係にあると考えられた.そして両方の効果を有する単一の成分として,我々はレスベラトロールを最初に見いだした(図3).同様の効果をもつ成分として,植物精油成分カルバクロール,シトラール,シトロネロール,ゲラニオールを見いだしている7–9).また,ビールホップ成分フムロンやパセリ成分クリシン等においても同様の効果が報告されており,COX-2とPPARを指標にして,レスベラトロールと類似の効果を有するフィトケミカルを探索できると考えている.
これらの知見は,植物二次代謝物生合成の視点から考察すると興味深い.レスベラトロールは,植物が細菌感染など環境からの刺激に対する防御として誘導されるスチルベン合成酵素(STS)によって作られる.STSを持つ植物はあまり多くはないが,STSはケルセチンやカテキンなどの生合成に関与するIII型カルコン合成酵素スーパーファミリーに属している.さらに,このファミリーには脂肪酸合成酵素サブユニットも含まれており,アラキドン酸やエイコサペンタエン酸(EPA)の生合成に関わる.これら脂肪酸は,COXの基質であり,かつPPARの内因性リガンドとしてヒトに効果をもたらす.
前述したように,COX-2遺伝子の発現調節機構の解析から,レスベラトロールによる細胞選択的なCOX-2の発現抑制にPPARγが関与することを明らかにした4).さらにレスベラトロールは,培養細胞系でPPARα,β/δ,γを選択的に活性化すること10, 11),脳卒中モデルマウスにおいてPPARα活性化を介し脳梗塞を抑制し,脳保護作用を持つことを明らかにした11).また,レスベラトロールを摂取したマウスの肝臓で,SIRT1がPPARα依存的に発現誘導されることも見いだしている.一方,SIRT1を活性化するとPPARαが活性化することが報告されており,両者は相互を活性化する関係にあると考えられる(図3).レスベラトロールによるSIRT1活性化はアロステリックな制御を受けることが報告されているが2),活性化濃度を考慮すると,我々はPPARがレスベラトロールの最初の標的であると考えている.
レスベラトロールによるPPARα活性化の分子作用機構を明らかにするため,種々のポリフェノールの化学構造とPPAR活性化を比較検討したところ,レスベラトロールの4′位の水酸基が活性化に関与すると考えられた12).さらにPPARα結合ドメインのX線構造解析データを基にした結合様式の予測から,4′位の水酸基がPPARαのTyr-314残基と水素結合し,レスベラトロールは直接PPARαを活性化し,効果を発揮する可能性を明らかにした12).
さらに,レスベラトロールによるPPARα活性化がPDE阻害やアデニル酸シクラーゼ活性化など細胞内のcAMPを増加させた条件で増強されることを見いだした12).注目すべきことに,cAMPだけではPPARα活性化は検出されなかった.この結果から,我々は以下のような機構を現在考えている.レスベラトロールがPPARを活性化すると,脂質代謝が活性化する.これによってβ酸化-酸化的リン酸化-電子伝達系によって細胞内ATPの増加とcAMPの減少が生じる.その結果,PPAR活性化が抑制されるように制御される.しかし,レスベラトロールはPDE阻害活性も同時に有しているため,PPARを持続的に活性化する.このようなフィードフォワードPPAR活性化が,レスベラトロールの持続的な摂取による生活習慣病予防に寄与する分子機構と考えている.
レスベラトロールは,心血管系疾患のリスク軽減に関わる分子として注目されてきた.レスベラトロールの血管に対する作用として,血管拡張作用,血小板凝集作用,生体防御作用などに関わる一酸化窒素(NO)の増加や血管内皮型NO合成酵素(eNOS)の発現誘導が報告されている.我々は,生理的条件により近い濃度のレスベラトロールで,正常ヒト臍帯静脈由来血管内皮細胞を処理した場合,eNOS遺伝子の発現が誘導されること,SIRT1が誘導されることを見いだした13).さらに生体の恒常性維持に関わるオートファジー関連遺伝子,活性酸素消去や抗炎症作用に関する遺伝子の発現が誘導されことを見いだし13),これらの遺伝子群の発現変動がレスベラトロールの効果に関与している可能性を明らかにした.オートファジーの活性化にPPARα活性化やcAMPが関与することが報告されており,レスベラトロールのcAMPを介するPPAR活性化にも関連していると予想される.
また近年,レスベラトールの作用にmicroRNA(miRNA)の発現調節が関与することが注目されている.ヒトマクロファージ様細胞における抗炎症性miR-663の発現誘導を介した炎症性miR-155の発現抑制や,乳がん細胞における腫瘍抑制性miR-16, miR-141, miR-143, miR-200cの発現誘導などが報告されている14).PPARsに関連するmiRNAも複数報告されている15).現在はまだ明らかにされていないが,ㇾスベラトロールによるPPAR活性化にもmiRNAが関与する可能性も考えられる.
我々のPPARαノックアウトマウスを用いた実験において,レスベラトロールによる効果には,系統による差,すなわち遺伝背景による差があることがわかった.また,栄養条件によってもその効果は異なっていた.これらの結果は,遺伝要因と食事などの環境要因が,食品機能成分のヒトへの応用を考えるときに非常に重要であることを意味している.ゲノムワイドな研究が進み,医療の分野ではゲノム情報に基づいたオーダーメイド医療が確立されつつある.医療費の削減を考えると,治療よりも予防への寄与が期待できる食品機能成分の分野において,ゲノム情報の利用を進めるべきであると考えている.ゲノム情報の視点と栄養など環境要因の視点を入れて初めて,食品機能成分のヒトへの応用が可能になると考えられる.
1) Sinclai, D.A. & Guarente, L. (2014) Small-molecule allosteric activators of sirtuins. Annu. Rev. Pharmacol. Toxicol., 54, 363–380.
2) Park, S.J., Ahmad, F., Philip, A., Baar, K., Williams, T., Luo, H., Ke, H., Rehmann, H., Taussig, R., Brown, A.L., et al. (2012) Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell, 148, 421–433.
3) Subbaramaiah, K., Chung, W.J., Michaluart, P., Telang, N., Tanabe, T., Inoue, H., Jang, M., Pezzuto, J.M., & Dannenberg, A.J. (1998) Resveratrol inhibits cyclooxygenase-2 transcription and activity in phorbol ester-treated human mammary epithelial cells. J. Biol. Chem., 273, 21875–21882.
4) Inoue, H., Tanabe, T., & Umesono, K. (2000) Feedback control of COX-2 expression through PPARγ. J. Biol. Chem., 275, 28028–28032.
5) Grosser, T., Fries, S., & FitzGerald, G.A. (2006) Biological basis for the cardiovascular consequences of COX-2 inhibition: therapeutic challenges and opportunities. J. Clin. Invest., 116, 4–15.
6) Michalik, L., Auwerx, J., Berger, J.P., Chatterjee, V.K., Glass, C.K., Gonzalez, F.J., Grimaldi, P.A., Kadowaki, T., Lazar, M.A., O’Rahilly, S., et al. (2006) International Union of Pharmacology. LXI. Peroxisome proliferator-activated receptors. Pharmacol. Rev., 58, 726–741.
7) Hotta, M., Nakata, R., Katsukawa, M., Hori, K., Takahashi, S., & Inoue, H. (2010) Carvacrol, a component of thyme oil, activates PPARalpha and gamma and suppresses COX-2 expression. J. Lipid Res., 51, 132–139.
8) Katsukawa, M., Nakata, R., Takizawa, Y., Hori, K., Takahashi, S., & Inoue, H. (2010) Citral, a component of lemongrass oil, activates PPARα and γ and suppresses COX-2 expression. Biochim. Biophys. Acta, 1801, 1214–1220.
9) Katsukawa, M., Nakata, R., Koeji, S., Hori, K., Takahashi, S., & Inoue, H. (2011) Citronellol and Geraniol, Components of Rose Oil, Activate PPAR α and γ and Suppress Cyclooxygenase-2 Expression. Biosci. Biotechnol. Biochem., 75, 1010–1012.
10) Inoue, H., Jiang, X.F., Katayama, T., Osada, S., Umedono, K., & Namura, S. (2003) Brain protection by resveratrol and fenofibrate against stroke requires peroxisome proliferator-activated receptor α in mice. Neurosci. Lett., 352, 203–206.
11) Tsukamoto, T., Nakata, R., Tamura, E., Kosuge, Y., Kariya, A., Katsukawa, M., Mishima, S., Ito, T., Iinuma, M., Akao, Y., et al. (2010) Vaticanol C, a resveratrol tetramer, activates PPARα and PPARβ/δ in vitro and in vivo. Nutr. Metab. (Lond.), 7, 46.
12) Takizawa, Y., Nakata, R., Fukuhara, K., Yamashita, H., Kubodera, H., & Inoue, H. (2015) The 4′-hydroxyl group of resveratrol is functionally important for direct activation of PPARα. PLoS One, 10, e0120865.
13) Takizawa, Y., Kosuge, Y., Awaji, H., Tamura, E., Takai, A., Yanai, T., Yamamoto, R., Kokame, K., Miyata, T., Nakata, R., et al. (2013) Upregulation of eNOS, SIRT1 and autophagy-related genes by repeated treatment with resveratrol in human umbilical vein endothelial cells. Br. J. Nutr., 110, 2150–2155.
14) Hagiwara, K., Kosaka, N., Yoshioka, Y., Takahashi, R.U., Takeshita, F., & Ochiya, T. (2012) Stilbene derivatives promote Ago2-dependent tumour-suppressive microRNA activity. Sci. Rep., 2, 314.
15) Jiexia, D., Meng, L., Xingyong, W., Xi, J., Shaohua, C., Chaohui, Y., & Youming, L. (2015) Effect of miR-34a in regulating steatosis by targeting PPARα expression in nonalcoholic fatty liver disease. Sci. Rep., 5, 13729.

奈良女子大学研究院生活環境科学系准教授.学術博士.
奈良女子大学研究院生活環境科学系教授.医学博士.

This page was created on 2018-06-26T15:06:06.8+09:00
This page was last modified on 2018-08-17T09:41:57.898+09:00
このサイトは(株)国際文献社によって運用されています。