S1PシャペロンとしてのApoMの機能ApoM as a S1P chaperone
群馬大学未来先端研究機構Gunma University Initiative for Advanced Research ◇ 〒371–8511 群馬県前橋市昭和町3–39–22 ◇ Showa-machi 3–39–22, Maebashi, Gunma 371–8511
発行日:2018年10月25日Published: October 25, 2018
スフィンゴシン1-リン酸(S1P)は,免疫系や心血管系などにおいて多彩な生理作用を発揮するリゾリン脂質性の生理活性脂質であり,その生理作用は,主に細胞膜に存在する五つのGタンパク質共役型受容体S1P1~5を介して発揮される.比較的高濃度のS1Pが常に血液中およびリンパ球液中を循環しており,循環S1Pの約60%が高密度リポタンパク質(HDL)に,残りの約35%がアルブミンに結合している.S1Pは,HDLが持つ抗炎症作用や血管保護作用などを担う主要因子の一つであり,HDL上のS1Pの生理活性は,S1P特異的な結合タンパク質であるアポリポタンパク質M(ApoM)によって調節されることが,最近の研究により明らかになってきた.本稿では,S1PシャペロンとしてのApoM同定の経緯や,ApoMの解析を通して明らかになってきたアルブミンとの機能面での違い,またさまざまな疾患との関わりなどについて概説する.
© 2018 公益社団法人日本生化学会© 2018 The Japanese Biochemical Society
スフィンゴシン1-リン酸(sphingosine 1-phosphate:S1P)は,スフィンゴシン骨格を持つリゾリン脂質性の生理活性脂質であり,免疫系や心血管系などにおいて多彩な生理作用を有している.S1Pの生理作用は,主に細胞膜に存在する五つのGタンパク質共役型受容体S1P1~5を介して発揮される.
プロスタグランジンやロイコトリエンなどの生理活性脂質が,局所で一過性に産生され,作用発揮後に速やかに代謝されることが多いのに対して,比較的高濃度のS1Pが,常に血液中およびリンパ球液中を循環している.循環S1Pの主要な供給細胞は,赤血球と血管内皮細胞であり,S1Pはそれぞれ特異的なS1Pトランスポーターによって細胞外に運搬された後,約60%が高密度リポタンパク質(HDL)に,残りの約35%がアルブミンに結合して循環にのる(詳細は西,川原の稿を参照).HDLは,末梢組織から余剰コレステロールの引き抜きを行うだけでなく,抗炎症作用,抗酸化作用,抗アポトーシス作用などさまざまな働きを持つ多機能粒子であり,その機能の一部は,HDLに含まれているS1Pによって発揮されていると考えられてきた.しかしながら,HDL上でS1Pがどのように保持されているのかは不明であったため,HDL上のS1Pの生理作用を正確に評価することは困難であった.
筆者らの研究グループは,HDL上のアポリポタンパク質M(apolipoprotein M:ApoM)が,特異的なS1Pキャリアタンパク質として機能することを同定した.ApoMは約5%のHDLにのみ見いだされるマイナーアポリポタンパク質であるが,S1PはApoMを持つHDLにのみ集積している.本稿では,S1PキャリアとしてのApoM同定の経緯や,ApoMの解析を通して明らかになってきたアルブミンとの機能面での違い,またさまざまな疾患との関わりなどについて概説する.
ApoMは,1999年にDahlbäckらのグループによって同定されたアポリポタンパク質である1).肝臓と腎臓にmRNAの発現が認められる.ヒトApoMは,188残基のアミノ酸からなり,N結合型グリコシル化部位を一つ持つ.SDS-PAGE上の分子量は約26 kDaである1).大部分のApoMはHDLに結合しており,残りはLDLやVLDL上に見いだされる2).ヒト血漿中のApoMの濃度は0.9 µmol/L前後であり,全HDL粒子中の約5%がApoMを持つと算定される2, 3).
ApoMは,アミノ末端に21残基からなる疎水性の分泌シグナルペプチドを持つが,ペプチダーゼによる認識配列を欠くため,切断を受けることなく分泌される.この疎水性のシグナルペプチドが,HDLへのアンカーの役割を担っていると考えられる.シグナルペプチドを欠く変異体ApoMは,HDL上に保持されることなく,腎臓で速やかに除去されることがわかっている4).
ApoMは,立体構造的にレチノール結合タンパク質やアポリポタンパク質D,プロスタグランジンD合成酵素などと同じリポカリンファミリーに属し,8本の逆平行のβストランドからなるβバレル構造を持つ(図1).このβバレルのポケットの中に,疎水性の分子を結合して運搬することが可能である.ApoMは,in vitroでミリスチン酸,パルミチン酸,ステアリン酸などの脂肪酸やレチノイン酸などとも結合することが報告されているが5, 6),生体内で意味のある結合なのかどうかは不明である.
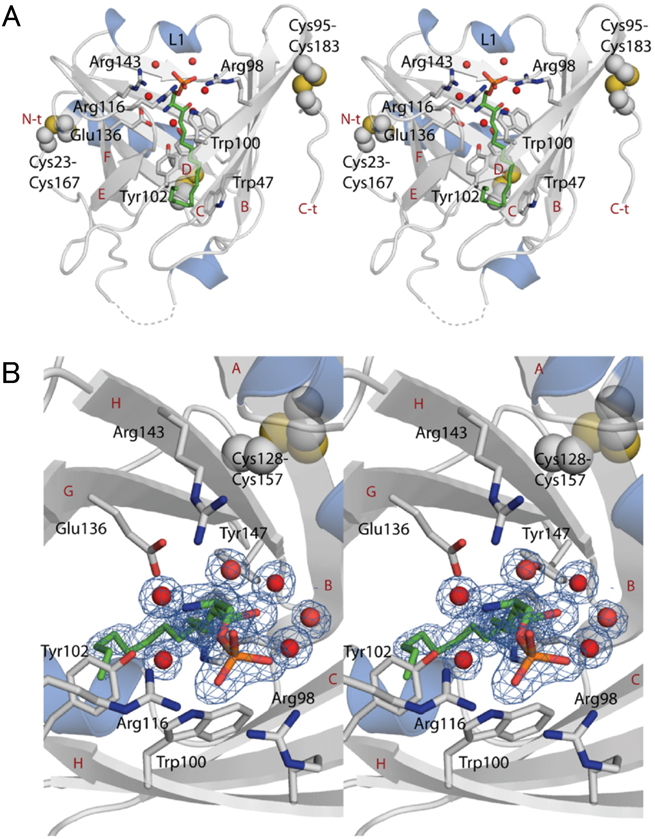
S1Pが結合した状態で解かれたApoMの結晶構造(解像度1.7 Å).S1Pの炭素鎖は緑色で,また酸素,リン,窒素原子はそれぞれ赤,橙,青で示されている.赤丸は水分子を示す.N-t, N末端,C-t, C末端,A-Hはβバレルを構成する八つのβストランドである.また分子内ジスルフィド結合を形成する3組6個のCys残基が示されている.細胞膜に平行な方向から見た側面図(A)および細胞外の方向から見たβバレル内部の拡大図(B)を示す.S1Pのリン酸基はArg98, Trp100, Arg116のアミノ酸側鎖と,またアミノ基はTyr102, Glu136, Arg143のアミノ酸側鎖と水素結合を形成している(文献7より引用).
1999年の同定以降,ApoMの機能解析は,ルンド大学(スウェーデン)のDahlbäckらと,コペンハーゲン大学(デンマーク)のNielsenらのグループが中心となって推進していた.ApoMの結晶構造を解析した結果,大腸菌で発現させたリコンビナントのApoMには,調製の過程でパルミチン酸やステアリン酸などが結合することが明らかとなった.炭素数16または18の骨格を持つ疎水性分子を中心にスクリーニングした結果,S1PがApoMの有力なリガンド候補として浮かび上がったため6),S1P研究の第一人者である,Timothy Hla博士(当時コーネル大学,米国)との共同研究が開始された.筆者がHla研究室に留学してまもなくのころであり,幸運なことにプロジェクトに加わることになった.
Nielsenらは,すでにApoMの欠損マウスとApoM発現量が異なる2系統のトランスジェニックマウスを所持していたため,はじめにこれらのマウス血漿中のS1P濃度の測定を行った.野生型マウス血漿では,S1P濃度が500 nM程度であるのに対し,ApoM欠損マウスでは約40%の200 nM程度に減少した(図2A)7).一方,ApoMを通常の2倍あるいは10倍ほどの濃度で発現するトランスジェニックマウスは,S1P濃度が野生型のそれぞれ160%あるいは350%に上昇し,血漿S1P濃度がApoMの発現量に応じて増減することがわかった.次に,抗ApoMモノクローナル抗体を用いたアフィニティカラムにより分画された,ApoMポジティブのHDL画分とApoMネガティブのHDL画分中のS1P濃度を測定すると,ApoMポジティブのHDLは,全体の5%程度にすぎないにもかかわらず,S1PはApoMポジティブのHDLにのみ検出された(図2B).これらの結果から,S1PはHDL中の脂質層に保持されているのではなく,ApoMに特異的に結合した状態でHDL上に保持されていると考えられる.
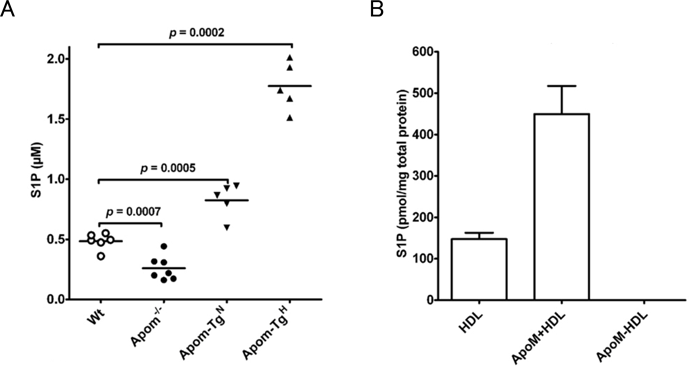
(A)野生型マウス(Wt),ApoM欠損マウス(Apom−/−),ApoMトランスジェニックマウス2系統(Apom-TgN,Apom-TgH; ApoMの発現量が野生型のそれぞれ2倍および10程度)の血漿中のS1P濃度.各点は各個体の値,横棒は平均値を表す.(B)ヒト血漿から調製したHDLを抗ApoMモノクローナル抗体を用いたアフィニティカラムによりApoMポジティブのHDL画分(ApoM+HDL)とApoMネガティブのHDL画分(ApoM−HDL)に分画し,それぞれのS1P濃度を測定した.S1PはApoMポジティブのHDL画分にのみ検出される(文献7より引用).
図1は,S1Pを結合させた状態で解かれたApoMの結晶構造である7).S1Pは,炭化水素鎖をβバレルの底部に向けた方向でポケットに入り込み,S1Pのリン酸基はArg98, Trp100, Arg116のアミノ酸側鎖と,またアミノ基はTyr102, Glu136, Arg143のアミノ酸側鎖と,水素結合を形成していることが読み取れる.生体内でApoMがS1P以外の脂溶性分子とも結合する可能性は否定できないが,結晶構造からApoMとS1Pの結合特異性が非常に高いことが示唆される.S1Pのリン酸基と相互作用するArg98, Trp100, Arg116の三つのアミノ酸をすべてAlaに変換した変異体は,S1Pの結合活性を失う8).
大腸菌で発現させたリコンビナントのApoMには,調製の過程でミリスチン酸などの脂肪酸が結合するので,この脂肪酸とS1Pとの置き換わりを,ApoMが持つ自家蛍光量の変化により測定すると,S1PのIC50は約0.9 µMと算定される6).測定手法により前後するが,血漿中のS1P濃度は数百nMから1 µM程度と算定されることから,0.9 µMというIC50は妥当な数値だと思われる.もう一つのS1P運搬タンパク質であるとされるアルブミンに関しては,S1Pとの結合親和性に関する情報がないが,さまざまな脂溶性因子を結合させることができるアルブミンの性質や,血漿中でのApoMとアルブミンの存在量比を考慮に入れた場合,S1Pに対する特異性や親和性はApoMに比べると低いと考えられる.
アルブミンに結合させた標識S1Pを,マウスに静注した実験結果によると,血中でのS1Pの半減期は約15分と非常に短い9).これは,血管内皮細胞表面に発現するlipid phosphate phosphataseなどにより分解を受けるためであると考えられる.一方,HDLに保持されているS1Pは,これよりずっと長い半減期を示す10).ApoMは,アルブミンに比べるとより高い親和性でS1Pをポケットの中に保持することで,S1Pを分解から保護し,S1P受容体までエスコートして適切なシグナル伝達を媒介すると考えられることから,S1Pシャペロンと表現されるようになった.
ApoMに結合したS1P(ApoM-S1P)は,アルブミンに結合したS1P(Alb-S1P)と同様に,S1P受容体の活性化能を保持しており,血管内皮細胞やCHO細胞などの培養細胞を用いたアッセイ系では,S1P1~3の各受容体を活性化し,細胞内シグナル伝達を惹起する7, 8).しかしながら,ApoM-S1PとAlb-S1Pとでは,機能的にさまざまな差異があることが明らかになってきた.なお,実験的にHDL(に保持されているApoM-S1P)を用いている場合と,HDL粒子に結合していないApoMを用いている場合とがあるが,これまでのところ両者に明確な機能的差異は見いだされていないので,本稿ではどちらもApoM-S1Pと表現した.
Alb-S1Pをリガンドとして用いたS1P1受容体の細胞培養系の解析では,S1P1受容体はGαiに選択的に共役し,実験的にはフォルスコリンによる細胞内cAMPの上昇を,S1Pとの共刺激により抑制することが示されていたが,ApoM-S1PではcAMPの産生抑制は観察されない11).また,Alb-S1Pは刺激後にS1P1受容体の内在化を強力に惹起するが,ApoM-S1P刺激では内在化の程度が非常に弱い.一方,Gαi経路の下流にあると考えられるERK/Aktの活性化や細胞の遊走活性では,両者にはほとんど差が観察されない.
血管内皮細胞において,S1P-S1P1受容体シグナルは,接着結合の強化により血管のバリア機能を上昇させる12, 13).単層の血管内皮細胞を用いて経内皮電気抵抗値(trans-endothelial electric resistance:TEER)の変化を測定した実験では,Alb-S1PによるTEERの上昇は30分ほどでピークに達した後,4時間程度で基底レベルに戻るのに対し,ApoM-S1Pでは同じく30分ほどでピークに達した後,20時間以上にわたって持続的にTEERを上昇させる14).この結果は,ApoM-S1Pが血管内皮細胞のバリア機能をより持続的に維持できることを示唆している.
これらのApoMとアルブミンの機能面での違いは,単純に共役しているGαタンパク質の違いだけでは説明がつかない.前述のとおり,ApoMに結合しているS1Pは,アルブミンの場合に比べて分解が遅いことから,より持続的に徐放性のシグナルを伝達できるのかもしれない.また,ApoM自身やHDL上の別の分子が,S1P受容体や他の受容体を介して,S1P受容体の活性化を調節している可能性も考えれる.非特異的な脂質キャリアとしての性質が強いアルブミンに対して,ApoMのS1P特異性は高く,生体内ではApoM-S1PがS1Pシャペロンとして,S1Pの運搬やシグナル伝達において主要な役割を果たしていると思われる.S1P受容体のシグナル伝達は,これまでAlb-S1Pを用いて大部分の解析が行われてきたが,ApoM-S1Pを用いて再評価する必要があると考えられる.
複数の疫学的な研究から,HDLコレステロールレベルと冠動脈疾患の罹患率が,逆相関することが明らかとなっている.末梢組織からコレステロールを引き抜いて肝臓に送り返す,HDLの逆輸送系の働きが重要であると考えられるが,これ以外にもHDLは抗酸化作用や血管内皮細胞に対する抗炎症作用など多面的な働きを持つ.したがって,循環HDL量を増加させることにより,冠動脈疾患の予防につなげようという試みが数多くなされてきたが,期待されるような成果は収められておらず,近年では,HDLの量よりも質の方が重要であると考えられるようになってきた15).しかしながら,HDLは種々の構成要素を持つヘテロな集団であり,HDL上の抗炎症作用の本体は明らかではなかった.S1Pは一つの候補であると考えられていたが,それを実験的に証明するのは困難であった.
S1PシャペロンとしてのApoMの同定により,HDLをS1Pを持つApoM+HDLと,S1Pを持たないApoM−HDLに分画することが可能となったため,これらのHDL画分が血管内皮細胞に対して発揮する抗炎症作用を検討した.炎症性サイトカインで刺激された血管内皮細胞では,ApoM+HDLは濃度依存的にNF-κB経路の活性化やICAM-1/VCAM-1(白血球接着因子;血管の炎症性マーカー)の発現上昇などを抑制したが,ApoM−HDLやAlb-S1Pでは,このような炎症反応の抑制効果がほとんど観察されなかった11).また,血管内皮細胞特異的に,S1P1受容体の欠損を誘導したマウスにおいては,大動脈の血管内皮細胞でICAM-1/VCAM-1の発現が上昇し,高脂肪食負荷による動脈硬化巣の形成が亢進することが観察された11).
これらの結果から,S1P-S1P1受容体シグナル伝達系は,血管の炎症抑制に寄与しており,ApoMに結合してHDL上に保持されているS1Pが,HDLの抗炎症作用を媒介する主要な因子の一つであることが強く示唆された.興味深いことに,Sattlerらは,冠動脈疾患を持つ患者群でHDL中のS1P含量が低下していることを報告している16).HDL中のS1P含量と冠動脈疾患罹患率の相関や,S1P含量低下の原因などは不明であり,今後の研究が待たれる.
ApoM欠損マウスでは,血漿中のHDL濃度が約20%減少し,逆にApoMトランスジェニックマウスでは約20%上昇する17).また,ApoM欠損によりサイズの大きい未成熟HDL粒子の増加や,HDL粒子へのコレステロールの取り込み不全が起こるという報告もなされている18).ApoMはHDL上に保持されているマイナーアポリポタンパク質の一つであるが,HDLの生成や成熟の過程に何かしらの影響を持つものと考えられる.
ApoM遺伝子改変マウスとLDL受容体欠損マウスをかけ合わせ,高脂肪食の負荷を与えたマウス動脈硬化モデルにおいては,ApoM欠損マウスで動脈硬化巣の増加が観察されているが19),血管内皮細胞特異的にS1P1受容体を欠損させた場合に比べると効果は限定的である11).ApoM欠損マウスでは約40%のS1PがAlb-S1Pとして残存しており,ApoM-S1Pの機能を代償するものと考えられる.また,ApoMトランスジェニックマウスを用いた動脈硬化モデルでは,動脈硬化巣の減少が観察されるが,やはり効果は限定的である19).血漿中には常にS1P1受容体を活性化するのに十分な量のS1Pが含まれているため,ApoMの過剰発現によりS1Pの循環量を増加させても,血管の慢性炎症を抑制する効果は低いのかもしれない.
S1P-S1P1受容体シグナル伝達系は,血管内皮細胞のバリア機能の維持に重要な役割を果たしており12, 13),ApoM欠損マウスではこのバリア機能が低下している.エバンスブルー色素や蛍光デキストランを尾静脈より注入すると,野生型では血管外にほとんど漏出しないのに対して,ApoM欠損マウスでは,末梢や肺などの毛細血管で漏出が観察される7, 20).したがって,ApoM欠損マウスでは,血管のバリア機能が低下し,血漿成分の漏出が起こりやすくなっていると考えられるが,通常の飼育条件下では目立った異常は観察されない.一方,カラギーナン(carrageenan,含硫黄多糖類)の足底部注入による足浮腫炎症モデルにおいては,初期相(~6時間)および遅発相(1日~)において,ともにApoM欠損マウスで浮腫の増大が観察され,この傾向はカラギーナン注入1週間後にまで遷延していた20).また,これらの浮腫の増大はアデノウイルスによるApoMの過剰発現や,S1P1受容体のアゴニスト投与により回復した.血管透過性の亢進は,炎症惹起の重要なステップの一つであり,ApoM-S1PによるS1P1受容体の活性化が血管透過性の制御に重要であることがわかる.
肝臓は高い再生能力を持つ臓器であるが,肝組織が再生する上で,類洞をはじめとする血管系の再形成が必要不可欠である.肝臓の部分切除による肝再生モデルにおいて,ApoM欠損マウスでは肝再生が遅延し,死亡率が大幅に上昇することが観察される21).ApoM欠損マウスでは,肝組織の再生の過程において類洞の形成不全や血栓の形成,肝組織の線維化などが観察され,これらはS1P1受容体のアゴニスト投与やApoMトランスジェニックマウスで軽減する.ApoM-S1PによるS1P1受容体の活性化は,類洞内皮細胞の増殖や血管網形成を促進するだけでなく,メカニズムの詳細は不明であるが,肝細胞の線維化を抑制する効果を持つことが示唆される.
ApoM欠損マウスにおいては,循環リンパ球数の増加が観察される22).これはApoM-S1PがS1P1受容体の活性化を介してリンパ球系共通前駆細胞の発生分化を抑制するためであることが示されている.一方,Alb-S1Pではこのような効果は認められない.実験的自己免疫性脳脊髄炎モデル(EAEモデル)において,ApoM欠損マウスは野生型より重篤な症状を示すことから22),ApoM-S1Pはリンパ球の発生分化だけでなく,獲得免疫反応においてもリンパ球の動態を制御していることが示唆される.
S1P-S1P受容体シグナル伝達系は,上述のように血管のバリア機能や炎症制御において重要な役割を果たすので,虚血性組織障害23)や高血圧症24),敗血症25, 26)などさまざまな疾患モデルにおいて,S1P-S1P受容体シグナル伝達系の増強が,有益な効果をもたらすことが報告されている.また,糖尿病患者のHDLではS1P含量が減少すること27)や,ApoM-S1Pがインスリン分泌を促すこと28)など,糖代謝との関連も報告されている.したがって,循環S1P濃度を上昇させることは,さまざまな疾患において有効な治療戦略となる可能性がある.
ApoMトランスジェニックマウスにおいて血漿のS1P濃度が上昇することから7),循環ApoM量を増加させることで血中S1P濃度を上昇させることが可能であると考えられる.しかしながら,HDLに結合していないApoMは,腎臓で速やかに除去されるため半減期が短い4).そこで,我々はApoMのC末端側に,免疫グロブリンのFc領域を融合させたタンパク質を作製した(ApoM-Fc).その結果,ApoM-Fcの血中半減期は約90時間と十分に安定であり(図3A),100 µgのApoM-Fcを腹腔投与することで,循環S1P量が約30%増加することが認められた(図3B)8).一方,S1Pのリン酸基と相互作用するArg98, Trp100, Arg116の三つのアミノ酸をAlaに変換したApoM変異体(ApoM-Fc-TM)は,S1Pを保持することができず,腹腔投与によるS1P濃度の上昇が観察されなかった.ApoM-Fcに結合したS1Pは,S1P受容体を活性化する能力を保持しており,ApoM-Fcの投与により,マウス高血圧モデルにおける血圧上昇(図3C)や,虚血再灌流実験における脳や心臓の組織障害(図3D)が,著明に低減することが観察された8).これらの効果は,血管内皮細胞のS1P1受容体活性化によるNO産生の増加,および血管透過性制御による炎症反応の軽減により,それぞれもたらされていると予測される.
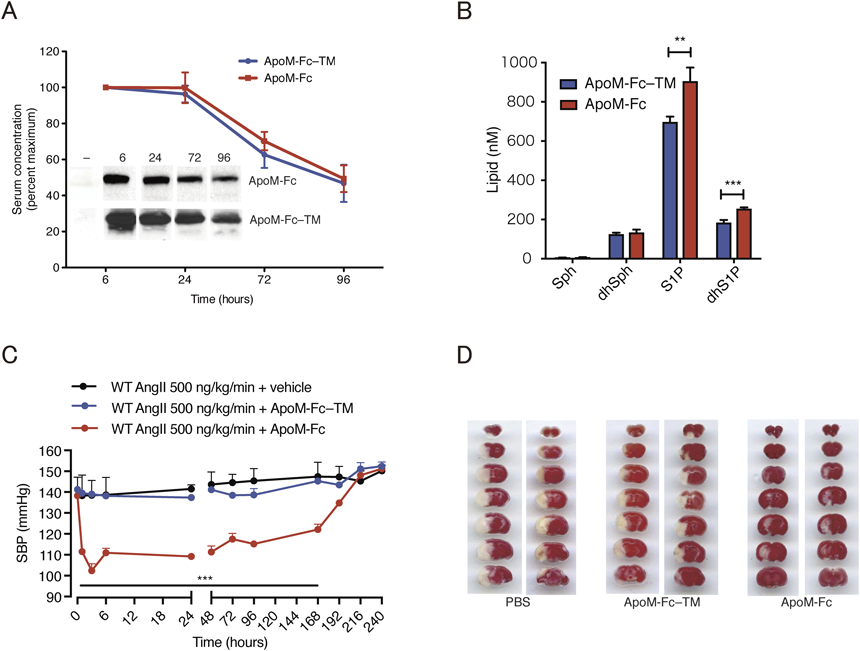
(A)免疫グロブリンのFc領域を融合させたApoM(ApoM-Fc)の血中半減期は約90時間と安定である.ApoM-Fc-TMはArg98, Trp100, Arg116の変異によりS1Pに結合できないApoM変異体コントロール.血中半減期は両者で違いが観察されない.(B) ApoM-Fc(100 µg)を腹腔投与24時間後のマウス血漿中では,S1PおよびジヒドロS1P(dhS1P)の濃度が約30%増加した.(C)アンジオテンシンIIの持続投与によるマウス高血圧モデルにおいて,ApoM-Fc(100 µg)の腹腔投与により収縮期血圧(SBP)の低下が観察されるが,ApoM-Fc-TMではそのような効果が認められない.(D)中大脳動脈結紮によるマウス脳梗塞モデル(MCAOモデル)において,ApoM-Fc(100 µg)の腹腔投与により虚血再灌流後の組織障害(白色部位)が軽減される(文献8より引用).
S1P受容体をターゲットとする小分子化合物は,臓器選択性が低く,免疫系や心血管系などさまざまな組織の受容体に働いてしまうが,ApoM-Fcでは循環リンパ球数の変化などは観察されず,血管内皮細胞のS1P1受容体シグナルを選択的に増強しているようである.ApoMトランスジェニックマウスを用いた動脈硬化モデルでは,ApoM発現量の増加による動脈硬化抑制の効果がそれほど高くないため19),慢性の炎症性疾患に対する効果は見込めない可能性もあるが,ApoM-Fcなどにより一過性に循環S1P濃度を上昇させ,S1P受容体シグナルを増強することが,高血圧や虚血性組織障害に対して有用な治療戦略となるかもしれない.
以上述べてきたように,ApoMはS1Pに特異的に結合してHDL上に保持し,S1P受容体までエスコートして適切なシグナル伝達を媒介する,S1Pシャペロンとして働く.ApoM-S1Pを保持するHDLは全体の約5%にすぎないが,血管内皮細胞に対する抗炎症作用や,バリア機能の増強などにおいて,有益な効果を担っており,いわゆる“質の高い” HDLを構成する因子の一つとして考えることができる.また,循環ApoM量を一過的に上昇させて,S1P受容体シグナルを増強することが,高血圧や虚血性組織障害に対して有用な治療戦略となる可能性も示された.これらの知見が,今後,さまざまな疾患の治療法・予防法の開発の礎となれば望外の喜びである.
S1PシャペロンとしてのApoMが同定されたことにより,リンパ球系共通前駆細胞の発生分化制御や糖代謝との関わりなど,アルブミンをS1Pキャリアとして用いたこれまでの研究では見いだされていなかった知見が,次々と明らかになってきている.ApoM-S1PとAlb-S1Pとで機能的差異が生まれる分子機序や,ApoMに結合したS1PがいかにS1P受容体に受け渡されるのかなど,解明すべき課題も多いが,ApoMにエスコートされたS1Pによる生体機能の制御機構がますます明らかになっていくことと期待される.
本稿は,筆者が2009年から2015年まで所属していたTimothy Hla研究室(米国コーネル大学,現ハーバード大学)において行った研究と,帰国後の共同研究の成果を中心に執筆しました.Hla教授と研究室の全メンバー,特にSylvain Galvani博士,Steven Swendeman博士,ラボマネージャーのCatherine Liu氏に深く感謝いたします.また,ApoMの共同研究を行ったDahlbäck博士,Nielsen博士,およびChristoffersen博士に深く感謝いたします.
1) Xu, N. & Dahlbäck, B. (1999) A novel human apolipoprotein (apoM). J. Biol. Chem., 274, 31286–31290.
2) Christoffersen, C., Nielsen, L.B., Axler, O., Andersson, A., Johnsen, A.H., & Dahlbäck, B. (2006) Isolation and characterization of human apolipoprotein M-containing lipoproteins. J. Lipid Res., 47, 1833–1843.
3) Axler, O., Ahnström, J., & Dahlbäck, B. (2007) An ELISA for apolipoprotein M reveals a strong correlation to total cholesterol in human plasma. J. Lipid Res., 48, 1772–1780.
4) Christoffersen, C., Ahnström, J., Axler, O., Christensen, E.I., Dahlbäck, B., & Nielsen, L.B. (2008) The signal peptide anchors apolipoprotein M in plasma lipoproteins and prevents rapid clearance of apolipoprotein M from plasma. J. Biol. Chem., 283, 18765–18772.
5) Ahnström, J., Faber, K., Axler, O., & Dahlbäck, B. (2007) Hydrophobic ligand binding properties of the human lipocalin apolipoprotein M. J. Lipid Res., 48, 1754–1762.
6) Sevvana, M., Ahnström, J., Egerer-Sieber, C., Lange, H.A., Dahlbäck, B., & Muller, Y.A. (2009) Serendipitous fatty acid binding reveals the structural determinants for ligand recognition in apolipoprotein M. J. Mol. Biol., 393, 920–936.
7) Christoffersen, C., Obinata, H., Kumaraswamy, S.B., Galvani, S., Ahnström, J., Sevvana, M., Egerer-Sieber, C., Muller, Y.A., Hla, T., Nielsen, L.B., et al. (2011) Endothelium-protective sphingosine-1-phosphate provided by HDL-associated apolipoprotein M. Proc. Natl. Acad. Sci. USA, 108, 9613–9618.
8) Swendeman, S.L., Xiong, Y., Cantalupo, A., Yuan, H., Burg, N., Hisano, Y., Cartier, A., Liu, C.H., Engelbrecht, E., Blaho, V., et al. (2017) An engineered S1P chaperone attenuates hypertension and ischemic injury. Sci. Signal., 10, eaal2722.
9) Venkataraman, K., Lee, Y.-M., Michaud, J., Thangada, S., Ai, Y., Bonkovsky, H.L., Parikh, N.S., Habrukowich, C., & Hla, T. (2008) Vascular endothelium as a contributor of plasma sphingosine 1-phosphate. Circ. Res., 102, 669–676.
10) Kimura, T., Sato, K., Kuwabara, A., Tomura, H., Ishiwara, M., Kobayashi, I., Ui, M., & Okajima, F. (2001) Sphingosine 1-phosphate may be a major component of plasma lipoproteins responsible for the cytoprotective actions in human umbilical vein endothelial cells. J. Biol. Chem., 276, 31780–31785.
11) Galvani, S., Sanson, M., Blaho, V.A., Swendeman, S.L., Obinata, H., Conger, H., Dahlbäck, B., Kono, M., Proia, R.L., Smith, J.D., et al. (2015) HDL-bound sphingosine 1-phosphate acts as a biased agonist for the endothelial cell receptor S1P1 to limit vascular inflammation. Sci. Signal., 8, ra79.
12) Lee, M.J., Thangada, S., Claffey, K.P., Ancellin, N., Liu, C.H., Kluk, M., Volpi, M., Sha’afi, R.I., & Hla, T. (1999) Vascular endothelial cell adherens junction assembly and morphogenesis induced by sphingosine-1-phosphate. Cell, 99, 301–312.
13) Garcia, J.G., Liu, F., Verin, A.D., Birukova, A., Dechert, M.A., Gerthoffer, W.T., Bamberg, J.R., & English, D. (2001) Sphingosine 1-phosphate promotes endothelial cell barrier integrity by Edg-dependent cytoskeletal rearrangement. J. Clin. Invest., 108, 689–701.
14) Wilkerson, B.A., Grass, G.D., Wing, S.B., Argraves, W.S., & Argraves, K.M. (2012) Sphingosine 1-phosphate (S1P) carrier-dependent regulation of endothelial barrier: high density lipoprotein (HDL)-S1P prolongs endothelial barrier enhancement as compared with albumin-S1P via effects on levels, trafficking, and signaling of S1P1. J. Biol. Chem., 287, 44645–44653.
15) Rader, D.J. & Tall, A.R. (2012) Bench to bedside:Is it time to revise the HDL cholesterol hypothesis? Nat. Med., 18, 1344–1346.
16) Sattler, K., Gräler, M., Keul, P., Weske, S., Reimann, C.-M., Jindrová, H., Kleinbongard, P., Sabbadini, R., Bröcker-Preuss, M., Erbel, R., et al. (2015) Defects of high-density lipoproteins in coronary artery disease caused by low sphingosine-1-phosphate content:correction by sphingosine-1-phosphate-loading. J. Am. Coll. Cardiol., 66, 1470–1485.
17) Christoffersen, C., Jauhiainen, M., Moser, M., Porse, B., Ehnholm, C., Boesl, M., Dahlbäck, B., & Nielsen, L.B. (2008) Effect of apolipoprotein M on high density lipoprotein metabolism and atherosclerosis in low density lipoprotein receptor knock-out mice. J. Biol. Chem., 283, 1839–1847.
18) Wolfrum, C., Poy, M.N., & Stoffel, M. (2005) Apolipoprotein M is required for prebeta-HDL formation and cholesterol efflux to HDL and protects against atherosclerosis. Nat. Med., 11, 418–422.
19) Christoffersen, C., Pedersen, T.X., Gordts, P.L.S.M., Roebroek, A.J.M., Dahlbäck, B., & Nielsen, L.B. (2010) Opposing effects of apolipoprotein m on catabolism of apolipoprotein B-containing lipoproteins and atherosclerosis. Circ. Res., 106, 1624–1634.
20) Christensen, P.M., Liu, C.H., Swendeman, S.L., Obinata, H., Qvortrup, K., Nielsen, L.B., Hla, T., Di Lorenzo, A., & Christoffersen, C. (2016) Impaired endothelial barrier function in apolipoprotein M-deficient mice is dependent on sphingosine-1-phosphate receptor 1. FASEB J., 30, 2351–2359.
21) Ding, B.-S., Liu, C.H., Sun, Y., Chen, Y., Swendeman, S.L., Jung, B., Chavez, D., Cao, Z., Christoffersen, C., Nielsen, L.B., et al. (2016) HDL activation of endothelial sphingosine-1-phosphate receptor-1 (S1P1) promotes regeneration and suppresses fibrosis in the liver. JCI Insight, 1, e87058.
22) Blaho, V.A., Galvani, S., Engelbrecht, E., Liu, C., Swendeman, S.L., Kono, M., Proia, R.L., Steinman, L., Han, M.H., & Hla, T. (2015) HDL-bound sphingosine-1-phosphate restrains lymphopoiesis and neuroinflammation. Nature, 523, 342–346.
23) Sanchez, T. (2016) Sphingosine-1-phosphate signaling in endothelial disorders. Curr. Atheroscler. Rep., 18, 31.
24) Cantalupo, A., Gargiulo, A., Dautaj, E., Liu, C., Zhang, Y., Hla, T., & Di Lorenzo, A. (2017) S1PR1 (sphingosine-1-phosphate receptor 1) signaling regulates blood flow and pressure. Hypertension, 70, 426–434.
25) Coldewey, S.M., Benetti, E., Collino, M., Pfeilschifter, J., Sponholz, C., Bauer, M., Huwiler, A., & Thiemermann, C. (2016) Elevation of serum sphingosine-1-phosphate attenuates impaired cardiac function in experimental sepsis. Sci. Rep., 6, 27594.
26) Zhu, B., Luo, G.-H., Feng, Y.-H., Yu, M.-M., Zhang, J., Wei, J., Yang, C., Xu, N., & Zhang, X.-Y. (2017) Apolipoprotein M protects against lipopolysaccharide-induced acute lung injury via sphingosine-1-phosphate signaling. Inflammation, 41, 643–653.
27) Brinck, J.W., Thomas, A., Lauer, E., Jornayvaz, F.R., Brulhart Meynet, M.-C., Prost, J.-C., Pataky, Z., Löfgren, P., Hoffstedt, J., Eriksson, M., et al. (2016) Diabetes mellitus is associated with reduced high-density lipoprotein sphingosine-1-phosphate content and impaired high-density lipoprotein cardiac cell protection. Arterioscler. Thromb. Vasc. Biol., 36, 817–824.
28) Kurano, M., Hara, M., Tsuneyama, K., Sakoda, H., Shimizu, T., Tsukamoto, K., Ikeda, H., & Yatomi, Y. (2014) Induction of insulin secretion by apolipoprotein M, a carrier for sphingosine 1-phosphate. Biochim. Biophys. Acta, 1841, 1217–1226.

群馬大学未来先端研究機構准教授.博士(医学).
1996年東京大学教育学部卒,2000年東京大学大学院医学系研究科中途退学,同年群馬大学医学部助手,09年米国Cornell大学博士研究員,12年より同インストラクター,15年より群馬大学大学院医学系研究科准教授を経て,16年より現職.
研究テーマと抱負脂質メディエーターの受容体,特に2009年にTimothy Hla教授のもとに留学してからは血管におけるS1Pの機能について研究を行っている.

This page was created on 2018-08-31T14:26:38.441+09:00
This page was last modified on 2018-10-17T13:39:10.11+09:00
このサイトは(株)国際文献社によって運用されています。