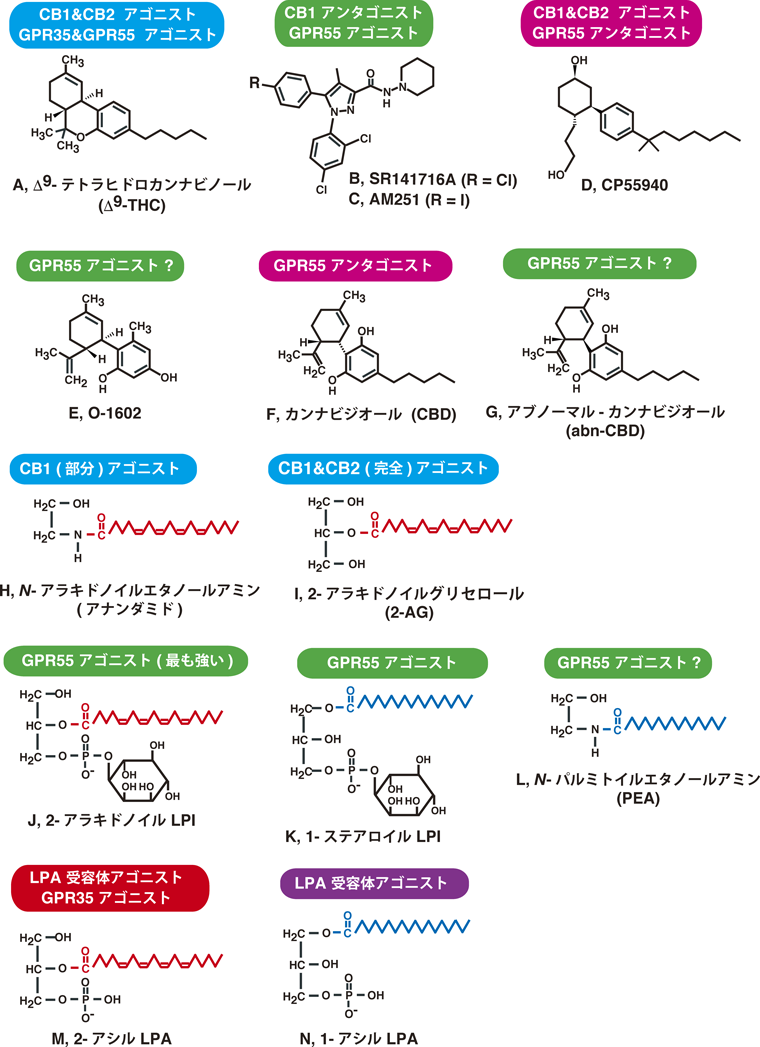
2. カンナビノイドCB1, CB2受容体と内在性リガンド
マリファナ(大麻)は多くの国で禁止薬物に指定されており,マリファナおよび類似薬物の乱用は社会的な問題になっている.マリファナを摂取すると,時間感覚・空間感覚の混乱,多幸感,記憶の障害,痛覚の低下,幻覚など多彩な精神神経反応がみられる.マリファナの作用の多くはΔ9-テトラヒドロカンナビノール(Δ9-THC)に代表されるカンナビノイド(植物由来なのでphytocannabinoidとも呼ばれる)と総称される化合物によるものである.Δ9-THCの精神作用は,主に脳などの中枢神経系に多く発現しているカンナビノイド受容体1(CB1受容体)を介する1, 2).CB1受容体は,Gタンパク質(Gi/Go)共役型の受容体で,特に黒質,線条体,淡蒼球,小脳の分子層,海馬,大脳皮質などで,多く発現している.
一方,マリファナ常習者は感染症やがんになりやすいことが知られており,マリファナの摂取は,免疫系を抑制することが考えられている.この作用は免疫組織に発現しているカンナビノイド受容体2(CB2受容体)を介する3).CB2受容体は脾臓や扁桃腺など,免疫系の組織や細胞に多く発現していることとも符合する.CB2受容体はCB1受容体と近縁の受容体で,Gi/Goに共役する.
CB1およびCB2受容体の研究の過程で,さまざまな合成カンナビノイドが合成された(図1,表1).それらの薬理学的な研究は,内在性リガンドの探索の際にも大きく貢献をしている.オピオイド受容体の場合と同様に,CB1受容体およびCB2受容体に対する内在性のアゴニスト(endocannabinoid)が存在することが想定され,その検索が行われた.Devane, Mechoulamらは,合成カンナビノイドのCB1受容体への結合を競合阻害する活性を指標に,ブタの脳から,N-アラキドノイルエタノールアミン(アナンダミド)を単離した4)(図1, H).我々のグループはアナンダミドと構造が類似する2-アラキドノイルグリセロール(2-AG)がCB1受容体のリガンドであることを発見した5)(図1, I).Mechoulamのグループもほぼ同時に2-AGを報告している6).我々は2-AGがCB1受容体を介して,細胞内カルシウムイオン濃度の上昇(Caレスポンス)を引き起こすことを見いだし,2-AGがCB1受容体のアゴニストであることを証明した7).
注目すべきことに,アナンダミドはCB1受容体に対して高い親和性を示すものの,アゴニストとして活性が弱く,部分アゴニストとして作用することがわかった8).また,我々は,通常の組織にアナンダミドはごく少量しか存在しないこと,虚血など病的な状態のときに大量に産生されることを見いだした.その生合成経路も,通常ほとんど存在しないリン脂質のグリセロールsn-1位のアラキドン酸が利用されるなど合理的なものでないことを示した9–11).この生合成経路は他の脂肪酸を持つN-アシルエタノールアミンを産生することが主の経路で,アナンダミドは副生成物である.一方,2-AGは,CB1およびCB2受容体の完全アゴニストとして作用する.生合成経路も,PI代謝回転が亢進した際に生じる,アラキドン酸含有ジアシルグリセロールがリパーゼで代謝されて生じることが想定される.これらのことから,カンナビノイドCB1, CB2受容体の本来の生理的なリガンドは,2-AGであることが強く示唆される.
中枢神経系において,CB1受容体はシナプス前終末に発現しており,その活性化は神経伝達物質の放出を抑制的に制御していると考えられている12–14)(図2, A).実際に,シナプス前膜から放出された神経伝達物質によって,シナプス後膜のPI代謝回転が亢進する.その際に生じた2-AGがシナプス間隙を輸送され,シナプス前膜のCB1受容体に作用することで,神経伝達物質の放出を抑制する.すなわち,2-AGは逆行性の神経抑制因子(retrograde negative messenger)として働くことが想定されている.また,Δ9-THCもアナンダミドと同様,CB1受容体の部分アゴニストであり,マリファナによって多幸感,記憶の障害,幻覚など多彩な精神神経反応が現れるのは,生理的なリガンドである2-AGの作用を撹乱することによることが考えられる.
CB2受容体の役割も,内在性アゴニスト2-AGの発見により明確となった.CB2受容体が炎症や免疫に促進的に,あるいは抑制的に作用するのかに関しては多くの議論があった.マリファナの摂取により感染症やがんが増えることから,CB2受容体は免疫系を抑制することが考えられていた.しかし,我々は,CB2受容体のアゴニストの2-AGが,マクロファージ様細胞の遊走や接着,サイトカインの産生に関与することなどから,CB2受容体とそのリガンド2-AGは炎症・免疫に促進的に働くことを示した15–18).CB1と同様にCB2受容体の場合においても,Δ9-THCは部分アゴニストまたはアンタゴニストとして2-AGの作用を抑えることで,免疫抑制作用がみられると考えられる(図2B).
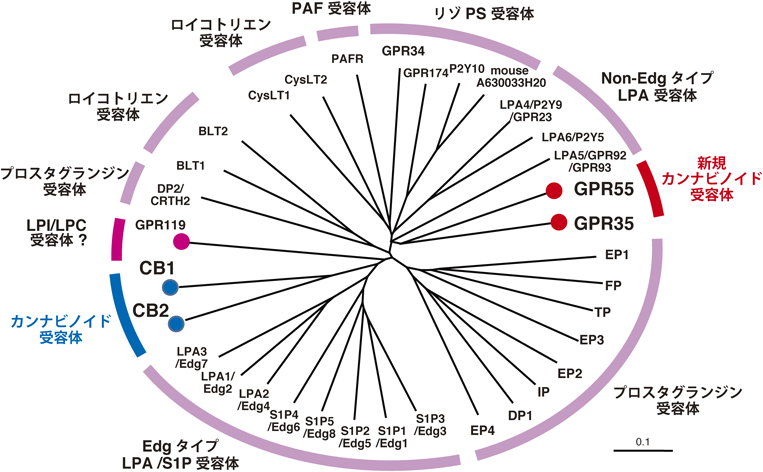
内在性リガンドの発見やノックアウトマウスの開発によって,マリファナのCB1, CB2受容体を介する多彩な薬理作用の解析は大きく進展した.しかし,カンナビノイド化合物の血圧低下作用が,CB1受容体とCB2受容体だけでは説明できないことが指摘され,新たな受容体の存在が想定された.2000年代の半ばに,製薬会社の特許の形で,それまでリガンドが不明であったGタンパク質共役型受容体のGPR55(G-protein coupled receptor 55)が新規のカンナビノイド受容体である可能性が報告された19, 20).もともとGPR55は脳に多く発現するGタンパク質共役型受容体としてクローニングされた分子21)で,CB1およびCB2受容体に相同性は低く,non-EDGタイプのLPA受容体と近縁であった(図3).その後,脳だけでなく,脾臓,腸管などに発現していることも確かめられた.ヒトの場合,319個のアミノ酸からなっており,G12/G13と共役していると考えられている(表1).ヒトGPR55遺伝子はイントロンを含まない遺伝子で,染色体2q37に位置している.
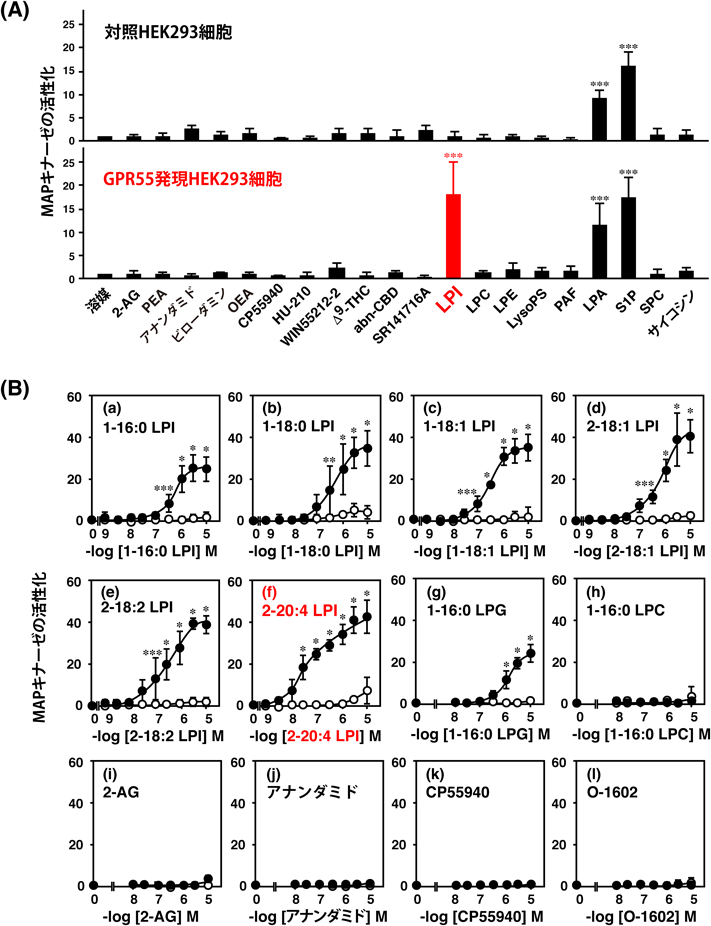
我々は,GPR55をHEK293細胞に発現させ,CaレスポンスやMAPキナーゼの活性化を指標に,カンナビノイド化合物の活性を検討した.Δ9-THCは高濃度(5 μM)でGPR55を介した反応を示し,GPR55がマリファナに反応する受容体であることを確認した.しかし,CB1およびCB2受容体のアゴニストとして開発された合成カンナビノイドのCP55940やWIN55212-2にはGPR55に対するアゴニスト活性は認められなかった22)(図4).
GPR55と天然および合成カンナビノイドとの反応性は相反する報告が存在し,結論が得られていない.マリファナの主要成分の一つのカンナビジオール(cannabidiol:CBD)は,Δ9-THCのような精神作用がないとされるが,GPR55のアンタゴニストとして作用するという報告がなされている23).また,カンナビジオールの立体異性体のアブノーマルカンナビジオール(abnormal cannabidiol:abn-CBD)やその構造類縁物のO-1602にGPR55アゴニストとしての活性があるという報告24)がある.しかし,我々を含めて否定的な結果25, 26)もあり,さらなる検討が必要である.
非常に興味深いことに,CB1受容体のアンタゴニスト(実際にはインバースアゴニスト)のAM251やSR141716Aに,GPR55のアゴニストとしての活性が認められると報告された26–28)(図1,表1).よって,実験動物に投与した際に観察されたAM251やSR141716Aのさまざまな影響の中には,CB1受容体アンタゴニストとしての作用によるものだけでなく,GPR55アゴニストとしての作用によるものも含まれている可能性がある.CB1およびCB2受容体とGPR55で,アゴニストとアンタゴニストと相反する作用を示すことは,非常に不思議なことである.後述するが,これらの受容体の内在性リガンドの構造が似ている[CB1受容体は2-AG, GPR55はアラキドン酸をグリセロールsn-2位に持つLPI(2-アラキドノイルLPI)]ことと関連するかもしれない.
我々はGPR55と近縁の受容体のGPR35が高濃度のΔ9-THCに反応することを見いだしている29)(図3).詳細は割愛するが,GPR35の内在性アゴニストは2-アシル型LPAであることがわかった(図1,表1).マリファナに反応する受容体がCB1およびCB2以外に複数あることは興味深い.
4. GPR55アゴニストとしてのリゾホスファチジルイノシトール
オーファン受容体のリガンドを同定すること(deorphaning)は,受容体の機能を知るために必須である.GPCR創薬を進める製薬会社もアゴニストおよびアンタゴニストをいちはやく検討できることから,オーファン受容体のリガンド探索に参入してきている.GPR55に関しても,製薬企業も含めさまざまなグループが内在性リガンドの同定に参入した.当初は論文ではなく,特許の書類として結果が公表されたので,混沌とした時期があった.
我々は,GPR55を発現させたHEK293細胞を用いて,ERK(p42/p44 MAPキナーゼ)のリン酸化と,Caレスポンスを指標に,内在性リガンドのスクリーニングを行った22).前述のように,我々はCB1とCB2受容体の内在性リガンドの2-AGを同定した経緯があったので,当初,GPR55の内在性リガンドも2-AGである可能性が高いと考えた.安定的にGPR55を発現しているHEK293細胞を用い,さまざまな実験系で2-AGの影響を調べたが,残念ながらこの仮説を支持する結果は得られなかった.また,アナンダミドを含む各種N-アシルエタノールアミンなど,さまざまなリガンド候補物質で細胞を刺激したが,GPR55発現HEK293細胞は反応しなかった.
GPR55がnon-EDGタイプのLPA受容体と近縁であることから(図2),LPAなどリゾリン脂質に反応する受容体である可能性を考え,各種,リゾリン脂質を試してみた22).GPR55発現HEK293細胞は,LPAとスフィンゴシン1-リン酸(S1P)に反応を示したが,この反応はHEK293細胞に存在する内在性のLPA受容体,S1P受容体によるもので,GPR55に対するものではなかった.
最後に,古くから生理活性が報告されているにもかかわらず,これまで受容体が同定されていなかったLPIを試してみた.その結果,GPR55を発現させたHEK293細胞で,LPIがERKの活性化を引き起こすことを見いだした.空ベクターを導入した細胞ではLPIによるERKの活性化が観察されなかったことより,GPR55の内在性アゴニストはLPIである可能性が高いことがわかった.
上述した会社の特許情報に,2-AGやアナンダミドをはじめとするさまざまな内在性カンナビノイドがGPR55のリガンドである可能性が言及されていたので,慎重にGPR55に対するLPIの活性の有無を検討した22, 25).我々はGPR55を発現させたHEK293細胞にLPIを作用させ,細胞内Caイオン濃度が速やかに一過的に上昇すること(Caレスポンス),空ベクターを導入した細胞ではLPIによるCaレスポンスはみられないことを見いだした.また,LPIによるGPR55発現細胞のCaレスポンスは,細胞をLPIで前処理することにより消失し,脱感作現象がみられること,GPR55に対するsiRNAをトランスフェクトするとLPIの作用が強く抑制されることから,LPIの作用は特異的な受容体すなわちGPR55を介したものであることが証明された.また,GPR55を発現させたHEK293細胞の膜画分に,LPI依存的なGTPγS結合活性が存在することより,GPR55はLPIの作用により,Gタンパク質を活性化することも示された.
LPIがGPR55を発現させた細胞においてCaレスポンスやERKの活性化を引き起こすことは,我々の報告のあと,いくつかのグループによって確認されている27, 30, 31).また,LPIがGPR55のアゴニストとして作用することは,β-アレスチンの解析28)や,受容体の内在化の解析によっても確認された27, 31).
LPIはさまざまな臓器に存在することが知られていたため,GPR55の内在性アゴニストであることが強く示唆される.そこで次に動物の体内に存在するLPIの詳細な分析を行った25).ラットの脳に含まれているLPIについて調べると,37.5 nmol/g組織のLPIが含まれていることがわかった.分子種としてはステアリン酸(18 : 0)を持つものが最も多く(50.5%),次いでアラキドン酸(20 : 4)を持つ分子種であった(22.1%)(表2).
表2 ラット脳に存在するLPIとその脂肪酸組成| 脂肪酸部分 | nmol/g (%) |
|---|
| 16 : 0 | 4.8 (12.8) |
| 16 : 1 | 0.5 (1.3) |
| 18 : 0 | 18.9 (50.5) |
| 18 : 1 | 5.0 (13.3) |
| 18 : 2 | trace |
| 20 : 4 | 8.3 (22.1) |
| 22 : 6 | trace |
| 計 | 37.5 (100.0) |
我々はLPIの生物活性を調べる際に,当初は市販品のダイズ由来のLPIを使用していた(ダイズPIをホスホリパーゼA2で処理したもの).これはsn-1位にパルミチン酸(16 : 0)とステアリン酸(18 : 0)を持つものの混合物である.我々は,sn-1位またはsn-2位に各種脂肪酸を持つLPIを調製し,GPR55に対する構造-活性相関を検討した25)(図4).興味深いことに,グリセロール骨格のsn-2位にアラキドン酸を持つLPI(EC50=30 nM)は,sn-1位にステアリン酸を持つLPI(EC50=450 nM)より,はるかに強い受容体活性化能を示した(10~20倍).2-アラキドノイルLPI(図1,表2)は,ラット脳に均一に存在し,その含量(8.29 nmol/g)は組織の80%を水と仮定すると10 μM程度と計算され,受容体を活性化する濃度より高い濃度で存在していた.このことからも2-アラキドノイルLPIはGPR55の生理的なリガンドとして働くと考えられる.
構造活性相関の検討の過程で,2-アラキドノイルLPIがホスホリパーゼC(PLC)やホスホリパーゼDで分解を受けて生じる2-AGや2-アラキドノイルLPAはGPR55のアゴニスト活性がないことが明らかになった.活性は2-アラキドノイルLPI自身によるもので,分解産物によるものではないことが確認された.
興味深いことに,リゾホスファチジルグリセロール(LPG)には,弱いながらGPR55発現細胞のCaレスポンスを惹起させる活性が認められた25)(図4).この事実は,GPR55のリゾリン脂質の極性頭部,特に糖部分の認識がそれほど厳密でないことを意味する.グリセロールがイノシトール環の半分の炭素原子を三つ持つ構造であることを考えると興味深い.理研の上口,平林らのグループは,リゾホスファチジルグルコースがGPR55のリガンドであることを報告した32)(本巻,上口の稿を参照).GPR55のリガンド認識を考えると反応すること自体は不思議ではない.今後,2-アラキドノイルLPIとリゾホスファチジルグルコースのどちらが生理的なアゴニストかを検証しなければならない.いずれにしても,GPR55がLPIに対する受容体であることは,疑いのない事実である.LPAやS1Pの受容体の命名にならって「GPR55/LPI1」としてもよいかもしれない.
1)免疫系への関与
GPR55は,当初,脳に多く発現する受容体として認識されていたが,脾臓,胸腺,精巣,小腸にも,多く発現している33, 34).この事実は,GPR55とその内在性リガンドであるLPIが,神経組織だけでなく免疫組織や消化管でも何らかの重要な役割を演じていることを強く示唆するものである.
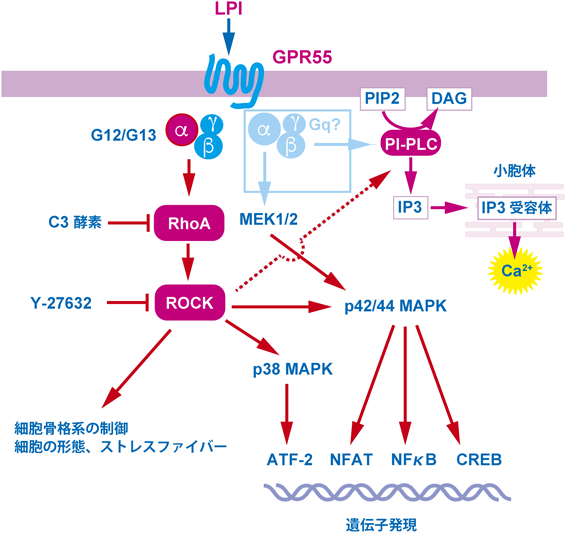
我々は,ヒトBリンパ芽球であるIM-9細胞がGPR55を発現していること,LPIで刺激することによりp38 MAPキナーゼやATF-2(activating transcription factor 2)の活性化を引き起こすことなどを見いだした33).GPR55を発現させたHEK293細胞も,内在性GPR55を発現しているIM-9細胞と同様に,LPI刺激によって,p38 MAPキナーゼやATF-2のリン酸化を引き起こした.我々は予備的に,GPR55を発現させたHEK293細胞が,LPIを作用させることにより,細胞の形態変化(rounding)やストレスファイバーの形成を起こすことを見いだしていたので,GPR55のシグナル伝達系が,三量体Gタンパク質のG12/G13や低分子量Gタンパク質のRhoAを介するかどうかを調べた.LPIによるp38 MAPキナーゼやATF-2の活性化は,RhoAを不活性化するボツリヌスC3酵素や,RhoAの下流で働くプロテインキナーゼのROCK(Rhoキナーゼ)の阻害剤Y-27632の前処理により消失した.これらの結果は,LPI-GPR55を起点として,G12/G13-RhoA- ROCK-p38 MAPキナーゼを介して,ATF-2を活性化することが考えられた(図5).ATF-2はサイトカインの発現に関する転写因子として知られているので,免疫細胞においてサイトカイン産生など重要な働きをしていることが考えられる.G12/G13-RhoA-ROCK系が関与することは,他のグループによっても確認されている31).先に述べたように,我々はLPI-GPR55の刺激が,細胞内Caの一過的な上昇を引き起こすことを報告したが22, 25),加えて持続的なCaオシレーションが起こることが示されている31).興味深いことに,持続的なCaオシレーションは初めにLPI-GPR55の刺激があれば十分で,Caオシレーションが起こっている際にLPIが存在しなくても起こることが示された.Caオシレーションは,G13αやRhoAのドミナントネガティブ変異体の発現やROCKの阻害剤の処理により消失した.また,NFAT(nuclear factor of activated T cells)を活性化することも報告された.NFATはT細胞のIL-2産生に関与するので,LPI-GPR55は免疫細胞において重要な働きをしていることが確認された.さらに,CREB(cAMP response element binding protein)やNFκB(nuclear factorκB)など他の転写因子の活性化26)などが報告されている.
一方,GPR55ノックアウトマウスでは,アジュバントで惹起した炎症局所の機械的な痛覚過敏が完全に消失することが示され,炎症性の痛覚過敏にGPR55が関与することが示された.また,GPR55ノックアウトマウスで,炎症局所のIL-4, IL-10,インターフェロンγ(IFN-γ),GM-CSFなどのレベルが上昇していることが示され,GPR55とそのリガンドLPIは,サイトカインの産生調節を介して炎症に関与することが考えられる35, 36).
Pietrらは,ミクログリアにGPR55が発現していること,LPIがGPR55を介してERKのリン酸化を引き起こすことなどを報告した36).GPR55やCB2受容体の発現が,リポ多糖処理により抑制されるのに対して,IFN-γにより上昇されることが示された.GPR55とCB2受容体が脳内炎症・免疫にも関与することが考えられる.
図5にLPI-GPR55を起点としたシグナル伝達系をまとめた.
2)神経系への関与
GPR55は脳に多く発現する受容体として発見されたのにもかかわらず,GPR55とそのリガンドLPIの脳や神経系における役割はまだ十分に明らかとなっていない.末梢神経やモデル細胞で,神経系における役割を類推することにとどまっていたが,しかし,最近になって,GPR55が記憶や不安,行動など高次作用に影響を与えることが報告された.
LPIが後根神経節(DRG)のニューロンにおいて細胞内Caイオン濃度の上昇を引き起こすこと,Δ9-THCがGPR55およびKチャネルであるKCNQ2, KCNQ3を発現させたHEK293細胞において,M電流を抑制することなどが報告されている30).GPR55はDRGに比較的高いレベルで発現しており,痛覚の調節に関与することが指摘されている35).また,LPIをラットの坐骨神経領域にマイクロインジェクションすると,坐骨神経が脱分極を起こし,ホットプレート試験における侵害受容の閾値を低下させた37).この作用はGPR55のアンタゴニストで消失することより,LPI-GPR55の遮断が,鎮痛作用につながることを示している.
GPR55は,脊髄の感覚性軸索の神経ガイダンスに重要な受容体であることが報告された32).すでに,GPR55が神経細胞のモデルとしてよく使われるPC12細胞の成長円錐(growth cone)に存在すること,LPIがPC12細胞の神経突起を退縮させることが報告されていた38)が,脊髄の感覚性軸索の場合,GPR55の刺激によって,神経突起の退縮は起こらず,軸索の先端が反発されるという負の化学遊走が観察されている.非常に興味深いことに,この場合のGPR55のアゴニストはLPIではなく,リゾホスファチジルグルコースであるということである.先に示したように我々は,GPR55の糖部分のリガンド認識は高くないことを示しているので,リゾホスファチジルグルコースがGPR55を刺激することは十分あると考えられる.同じくGPR55を刺激するLPIにも同様なガイダンス効果がみられるが,この効果はGPR55のノックアウトマウスでも観察される.この結果はGPR55以外のLPI受容体の存在を意味しているかもしれない.
また,GPR55が網膜の成長円錐の形態,神経節細胞の軸索の伸長を促進し,さらに軸索のガイダンスに関与することが報告された39).網膜神経の軸索は,GPR55のアゴニストのLPIおよびO-1602に対する化学誘引効果が観察され,アゴニスト処理によって成長円錐の大きさや糸状仮足の数が増加した.一方,アンタゴニストのカンナビジオールには逆の作用があり,軸索の化学反発がみられた.GPR55による神経軸索の誘引または反発という相反する作用は,神経細胞の種類によるものかもしれない.
GPR55の脳の高次機能における役割が報告され,GPR55が海馬CA3-CA1のシナプスの伝達を増強させること,すなわち短期増強に関与する可能性が示唆された40).この作用はノックアウトマウスやアンタゴニストのカンナビジオールによってなくなることからGPR55に依存することが示された.GPR55は海馬において記憶の一端に関与する可能性がある.一方,GPR55が記憶の障害に関与する可能性が報告された41).GPR55は,認識や記憶,感情など重要な働きをする海馬に発現している.特に腹側海馬は中脳辺縁系のドパミン神経系と相互作用して文脈記憶に関与する.パルミトイルエタノールアミン(PEA)によってGPR55を刺激すると,中脳辺縁系のドパミン神経系を刺激した.PEA-GPR55刺激は,オピオイドの報酬記憶の形成には影響を与えなかったが,社会的相互作用および認識記憶,空間位置記憶,および文脈独立性の連想恐怖記憶形成を強く混乱させた.これらのことはGPR55選択的アンタゴニストのCID16020046の処理で消失した.この報告ではGPR55刺激が記憶などを障害する可能性が示唆されているが,アゴニストとしてPEAを使っていることなど,精査する必要がある.
内在性カンナビノイドが線条体のGABA神経を調節することが報告された42).PEAはGPR55を介して,抑制性のGABA神経を刺激し,抑制性シナプス後電流(sIPSC)の周波数(頻度)を増加させた.これは(PEAはGPR55を介して)シナプス後膜で2-AGの産生を引き起こし,2-AGはシナプス前膜のCB1受容体を介して,GABAの放出を阻害した.先に述べたように,2-AGは興奮性神経の負の逆行性メッセンジャーとして,興奮性神経の活動を抑制することに働くが(図2A),PEAはGPR55による抑制性神経の抑制を介して,間接的に興奮神経のファインチューニングをしていることが示された.この報告ではGPR55に作用する内在性カンナビノイドとしてPEAを想定しているが,LPIの場合はどうなるか,今後の研究が待たれる.
GPR55遺伝子の多型と神経性無食欲症との関連が指摘されている43).Gly195Valのアミノ酸置換を起こすミスセンス多型が神経性無食欲症と関連していることが報告された.興味深いことに,Gly195Val GPR55を発現したCHO細胞は,野生型GPR55を発現させた細胞に比べ,LPI刺激に応答したERKの活性化が低下していた.このことはGPR55の機能低下によって食欲が抑制されることを示す.逆が必ずしも真であるとは限らないが,GPR55は食欲を促進する受容体かもしれない.
GPR55のアンタゴニストのカンナビジオールはてんかん発作や自閉症を低減させることがわかった44).Dravet症候群は乳幼児期に発症する難治性のてんかんで,そのマウスモデルを用いて,カンナビジオールの投与がてんかん発作を低減させることを示した.また,低用量のカンナビジオールは,モデルマウスの自閉症様の社会的障害を改善した.これらの現象は発作伝播の重要な領域である海馬歯状回の抑制性のニューロンの興奮性の回復と関連していた.カンナビノイド化合物の医療応用が考えられているが,その先駆けとなるかもしれない.
GPR55のアゴニストは,パーキンソン病の治療への応用の可能性も考えられている45).GPR55のノックアウトマウスは運動が低下することが知られているので,GPR55が運動に関与する可能性がある.GPR55は線条体に多く発現しているが,パーキンソン病になると,その発現量が下がる.GPR55のアゴニストのabn-CBDは,パーキンソン病誘導剤の1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine and probenecid(MPTPp)の投与で起こる運動低下を予防した.GPR55のアゴニストはパーキンソン病治療薬になる可能性がある.
3)骨代謝への関与
GPR55は破骨細胞に高いレベルで発現しており,骨代謝に関与している可能性が指摘されている24).破骨細胞に発現しているGPR55 mRNAのレベルは,血中の単球のレベルの8倍に達する.LPIや合成カンナビノイドであるO-1602で破骨細胞を刺激すると,ERKのリン酸化やRhoAの活性化,アクチンの重合などが観察される.また,LPIやO-1602は骨髄マクロファージからの破骨細胞の生成を抑制した.これらのLPIやO-1602の効果は,GPR55ノックアウトマウスから得られた破骨細胞や骨髄マクロファージではまったく認められなかった.注目すべきことに,雄のGPR55ノックアウトマウスにおいて,海綿骨の骨量の増加が観察された.これに対し,雌の場合,GPR55ノックアウトマウスと正常マウスの間で骨量に顕著な差は認められなかった.どうして雌雄の間でこのような違いが生じるのか,その理由はわかっていないが,内分泌系の影響等が考えられる.欠損マウスで骨量に変化がみられたことから,GPR55はヒトにおいても何らかの骨疾患に関与している可能性が考えられるが,これを示す知見は今のところ得られていない.
4)がんへの関与
GPR55とがんの増殖の関係が議論されている.GPR55が発見されるよりも以前に,LPIが細胞増殖を促進することが報告されている.Rasでトランスフォームしたラット甲状腺上皮由来細胞FRTL5細胞やKiKi細胞にLPIを作用させると,核へのチミジンの取り込みが亢進し,細胞増殖が誘導されることが見いだされた46, 47).それに先立ち,LPI刺激によってPI代謝亢進と細胞内Caイオン濃度の上昇,アラキドン酸の遊離などが観察されており,これらのシグナル伝達が最終的にDNA合成につながると考えられる.また,K-Rasをトランスフォームした繊維芽細胞がLPIを分泌することも報告され,LPIがオートクリンな作用で,細胞増殖を促進することが示された48).
GPR55がLPIの受容体であることが同定された後も,がんとLPI, GPR55の関連に関してさらに詳細に解析され,LPIがGPR55を介して,細胞増殖につながる鍵酵素であるMAPキナーゼを活性化すること49),siRNAによるGPR55のノックダウンが,卵巣がん細胞OVCAR3や前立腺がん細胞PC-3の細胞増殖を完全に停止させること50)が報告された.各種がん細胞のGPR55の発現解析により,乳がんの増殖性,悪性度などはGPR55の発現と相関することが示された51).同様な結果は,膵臓がんや神経膠腫(グリオーマ)でも確認されている.
がん細胞の遊走,転移とGPR55の関係も調べられている52).高い転移能を持つことで知られるMDA-MB-231乳がん細胞は,転移能が低いMCF-7細胞に比べ,GPR55が30倍も高く発現していた.MDA-MB-231細胞をLPIで処理すると,血清に対する遊走が上昇した.GPR55の発現が低いMCF-7細胞は,血清に対する遊走や浸潤を示さなかったが,MCF-7細胞にGPR55を発現させると,血清に対する遊走・浸潤がみられ,それらはLPIの処理によって増幅した.siRNAによりGPR55をノックダウンすると,遊走・浸潤が完全に阻害された.これらの結果を総合すると,LPIおよびGPR55はがん細胞の遊走・浸潤や転移に関与することが考えられた.
卵巣がんや腹膜がんの患者の血液や腹水の脂質性メディエーターを質量分析計で解析すると,健常人に比べて,LPIが蓄積していることが示された53–56).これらの結果は,LPI-GPR55とがんの進展の関係を示すものであり,LPIががんのバイオマーカーとして,臨床応用できることを示している.
5)肥満,糖尿病への関与
GPR55がLPI受容体と同定される前から,LPIの生理活性は報告されていた.LPIの生理活性は最初,膵臓におけるインスリン分泌を促進する物質として報告された57, 58).膵島をLPIで刺激すると,細胞内Caの上昇を介してインスリンが分泌された.この分泌は生理的なもので,エキソサイトーシスを介していた.
LPI-GPR55と肥満,2型糖尿病の関連が指摘されている59).脂肪細胞のGPR55の発現レベルは,肥満や2型糖尿病の患者で高いことが報告されている.内臓脂肪組織のGPR55の発現は,特に女性において,体重,BMI(body mass index:体格指数),体脂肪率と高い相関性がある.また,循環系のLPI量も,特に女性で,体重,BMI,体脂肪率と相関性があった.ヒトから取り出した内臓脂肪にLPIを作用させると,脂質合成酵素の発現量を増加させたり,細胞内Caを上昇させた.これらはLPI/GPR55と肥満に正の相関があることを示している.
また,GPR55の発現量が,ホルモンの状態により変化することが報告された60).ラットの白色脂肪のGPR55およびCB1受容体の発現量は,絶食後に増加し,レプチン処置により回復した.また,GPR55およびCB1受容体の発現量は妊娠により減少し,加齢によって増加した.興味深いことに,雄ラットの精巣を摘出すると,白色脂肪のGPR55およびCB1受容体の発現量は減少した一方,雌ラットの卵巣を摘出すると,GPR55の発現は増加したが,CB1の発現は低下した.GPR55の発現量がホルモンの状態で変化することは,ホルモンによる代謝変化とGPR55が密接に関係していることを意味し,興味深い.
6)血管系への関与
血管系にCB1, CB2受容体とは異なる未知のカンナビノイド受容体が存在していること,アナンダミドやabn-CBDはこの受容体を介して血圧を低下させることなどが,CB1/CB2ダブルノックアウトマウスに関する研究などから示唆されていた23).GPR55を発現させたHEK293細胞の膜画分は,O-1602やabn-CBDで刺激すると,GTPγSの結合を促進させた(EC50=2 nM).しかし,GPR55のノックアウトマウスの血圧は対照マウスと変わらず,abn-CBDを投与した際にみられる血圧降下も,ノックアウトマウスと対照マウスで変化がなかった.GPR55が血圧降下に関与する可能性は低く,血圧低下に関与する新たなカンナビノイド受容体が存在することが示唆される.
GPR55が血管内皮細胞に発現していることはすでに確認されている.GPR55とその内在性リガンドは,血圧の調節に関わる主要な因子ではないかもしれないが,場合によって影響を与えうるものである可能性がある.GPR55は消化管,特に小腸に多く発現していることが知られている25, 33).GPR55とその内在性リガンドは,小腸における消化・吸収や,腸管免疫等において何らかの重要な役割を担っている可能性があるが,詳しいことはわかっていない.
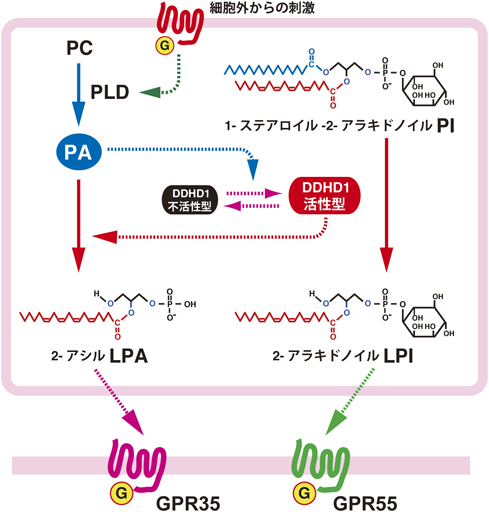
動物組織に存在するPIは,主に1-ステアロイル-2-アラキドノイル分子種からなっている61, 62).先に示したように,脳には2-アラキドノイルLPIや1-ステアロイルLPIが存在しており,これらは1-ステアロイル-2-アラキドノイル分子種が,ホスホリパーゼA1(PLA1)またはホスホリパーゼA2(PLA2)の作用を受けて生じることが考えられる.我々は最も活性が強い2-アラキドノイルLPIの産生酵素としてのPLA1の同定を試みた.PIは多くが細胞内に存在することから,細胞内型PLA1の関与を調べた.
現在まで,細胞内型PLA1は3種類しか知られていない.我々は,Glomsetらが同定したin vitroの反応でホスファチジン酸(PA)をよい基質とするとされたPLA1(PA-preferring PLA1: PA-PLA1)63)を候補としてその性状を検討した64).PA-PLA1は遺伝子名からDDHD1とも呼ばれ,脳をはじめとする各臓器に広く発現していること,酸性リン脂質のPIも基質とする可能性が考えられることなどが候補とした理由の一つである.Glomsetらも指摘しているように,この酵素はPA以外にさまざまなリン脂質を基質とすることから,PA-PLA1という名称は酵素の実際の性質を反映していない.そこで,この酵素を便宜的にNCBIのデータベースにおける登録名DDHD domain containing 1(DDHD1)で表し,DDHD1/PA-PLA1と併記した.DDHD domainは,KIAA0725p/DDHD2,膜結合性PI輸送タンパク質,網膜変性症タンパク質などに保存されている機能未知のドメインである.
DDHD1/PA-PLA1をHEK293細胞に発現し,サイトゾルから酵素を精製した64).Triton X-100の混合ミセルを用いたアッセイで,精製酵素の基質特異性を検討すると,既報どおりPAに対し高いPLA1活性を示した(見かけのKm=4 mol%).PIに対して親和性は低いものの(見かけのKm=10 mol%),十分な活性が検出された.PAとPIの細胞内の存在比を考えると,DDHD1/PA-PLA1はPIを十分に基質としうることが示された.
さらに,HEK293細胞にDDHD1/PA-PLA1を発現させ,LPIを産生するかどうかを調べた64).しかし,DDHD1を発現させるだけでは,LPIの産生能力は低いものでしかなかった.細胞をイオノマイシンで刺激すると,LPIの著しい生成増大が観察された.DDHD1を不可逆的に阻害するメチルアラキドノイルフルオロホスホナート(MAFP)で細胞を前処理すると,LPIの産生は阻害された.これらの結果は,DDHD1が刺激に応答したLPIの産生に関与することを示している.
イオノマイシン刺激がどのような機構でDDHD1を活性化するかを次に調べた64).我々は,イオノマイシン刺激によって生成する物質,特に脂質がDDHD1を活性化する可能性を検討した.DDHD1は可溶性の酵素で,PLA1として機能するためには基質の存在する膜に結合する必要がある.細胞をイオノマイシンで刺激すると,DDHD1の一部が膜画分に移行することが確認された.精製DDHD1がリン脂質に結合するかどうかを確認したところ,DDHD1はホスファチジルコリン(PC)やホスファチジルエタノールアミン(PE)にはほとんど結合しないが,PAには強く結合することがわかった.また,イオノマイシンで刺激した細胞においてホスホリパーゼD(PLD)の活性が亢進していること,PLDの作用を阻害するn-ブタノールを培地に添加すると,イオノマイシンで刺激した細胞のLPI生成亢進がほぼ完全に阻害されたことなどから,DDHD1の活性化にPLDによるPAの産生系が関与していることがわかった(図6).
精製DDHD1にPAを添加すると,PIに対するPLA1活性は顕著に上昇し,PAがDDHD1を直接活性化することも明らかになった.なお,DDHD1はPAに対するPLA1活性も持つため,PAが分解されて減少するとDDHD1の活性化は減衰することになる.
DDHD1は脳をはじめとする各臓器に広く発現しており,2-アラキドノイルLPIの産生酵素として実際に機能している可能性が高い.また,PLDと共役して活性化されるので,2-アシル型LPAの産生経路としても機能する可能性がある.我々はGPR55と近縁の受容体GPR35が(図3),Δ9-THCに反応すること,2-アシル型LPAが内在性アゴニストであること29)(図1,表1)を報告した.DDHD1はリガンドの供給を介して,GPR55だけでなく,GPR35とも共役している可能性が考えられる.これらの受容体が新規の内在性カンナビノイド受容体であることを考えると興味深い.
興味深いことに,遺伝性痙性対麻痺(hereditary spastic paraplegia)のSPG28亜種の原因遺伝子として,DDHD1が同定された65).痙性対麻痺は下肢の痙縮と筋力低下を呈する神経変性疾患で,非常に多くの原因遺伝子が同定され,現在,SPG1~72に分類されている.DDHD1のSPG28変異はミトコンドリアの呼吸低下を示し,このことが神経細胞死,神経変性につながることが考えられる.DDHD1とミトコンドリアの形態の関連は,DDHD1のノックアウトマウスを用いた解析によっても明らかにされた66).また,DDHD1の類似酵素であるDDHD2/KIAA0725pの変異も遺伝性痙性対麻痺SPG54を起こすことも報告された67).PLA1の変異が病態と関連することを意味している.LPIやLPAはグルタミン酸や虚血で誘導される神経細胞死に保護効果があることが知られている68, 69).神経変性過程で,PLA1の変異がLPIやLPAの産生低下と関連するか,GPR55やGPR35は関与するかなど,今後の研究の発展が期待される.
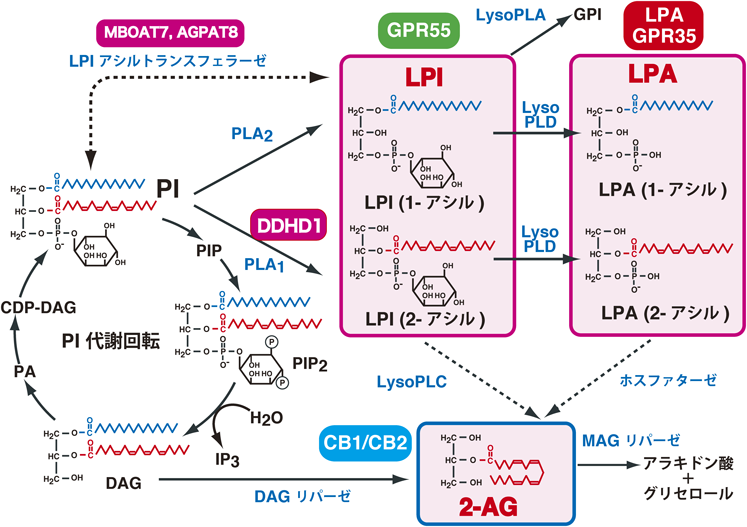
ラットの脳には,1-アラキドノイルLPIの他に,1-ステアロイルLPIが多量に存在している(表2).この事実は,ラットの脳にはPIを基質とするPLA2が存在することを示唆するものである.今のところ,PIに特異的なPLA2は知られていないが,Mariggiòらは細胞質PLA2(cPLA2α, PLA2G4A)がPIを段階的に分解し,1-アシルLPI, GPIを生成することを報告している70).前立腺がんや卵巣がん細胞のcPLA2αをsiRNAでノックダウンすると細胞増殖が低下することが報告された50).これらのがん細胞はLPI-GPR55によって,細胞増殖が促進されることが知られているので,cPLA2αによって産生される1-ステアロイルLPIが増殖の鍵分子となっていることが考えられる.cPLA2αはがん細胞だけでなく,脳をはじめとするさまざまな臓器に広く発現しているので,1-ステアロイルLPIの産生酵素として,動物の体内で実際に機能している可能性がある.
LPIの分解経路について詳細はわかっていない.LPIは前述したcPLA2αやリゾホスホリパーゼによって分解され,GPIと脂肪酸になる.一方,LPIはPLCによって分解され,モノアシルグリセロールとイノシトールリン酸になる(図7).
ところで,ラットの脳には古くからLPIに特異的なPLCが存在していることが報告されている71, 72).興味深いことに,この酵素はシナプトソームの膜画分に局在しており,PIやPC, PEをほとんど分解しない.基質となるLPIが,2-アラキドノイルLPIの場合,CB1受容体の内在性リガンドの2-AGが生成する.したがって,この酵素はLPIの分解酵素であるとともに,2-AGの生成酵素の一つでもあることになる.LPAの産生酵素のオートタキシン(リゾホスホリパーゼD)は,LPIも基質とすることができ,LPIからLPAを産生する73).2-アラキドノイルLPIが基質の場合,2-アラキドノイルLPAが生成し,これはGPR35, LPA3, LPA6など2-アシルLPAをリガンドとする受容体を活性化する.さらに,2-アラキドノイルLPAがホスファターゼで分解されると,2-AGが生成する74).これらの酵素は,単に脂質メディエーターを分解,消去するだけでなく,別々の受容体に作用する二つの脂質メディエーターをリンクさせる重要な鍵酵素であり,その実体の分子レベルでの解明が待たれる(図7).
LPIはLPAとともに,古くから生理活性があることが報告されていた.しかし,受容体がわかっていなかったこともあり,研究は停滞していた.我々が,LPIの受容体を同定したことを契機に,LPI-GPR55の新しい機能が次々に明らかになっている.
しかし,GPR55のリガンド探索の過程で,その活性が真かどうか,混乱した時期があった.我々はHEK293細胞の発現系を用いて慎重にリガンド探索を行ったが,他の研究者が報告しているO-1602, abn-CBDやパルミトイルエタノールアミン(PEA)のアゴニスト活性を検出することはできなかった.最近,GPR55の脳高次機能への関与が報告されたが,アゴニストとしてPEAを用いているものもあり,解釈に注意を要する.一つの実験系では,アゴニスト活性を正確に判断できない可能性がある.
合成カンナビノイドをはじめとするさまざまな化合物の薬理研究は,CB1, CB2, GPR55, GPR35など,マリファナに反応する受容体の研究に貢献してきた.合成カンナビノイドのアゴニストとアンタゴニストの活性が受容体によって反転することが知られている.CB1受容体アンタゴニストのSR141716A(リモナバン)は食欲を抑制するので,抗肥満薬としての応用が試されていた.しかし,臨床試験で有害事象が発生し,開発を断念した経緯がある.リモナバンがGPR55のアゴニストとして働くことが原因の一つである可能性がある.その応用には十分な注意が必要である.
LPIとその受容体に関する研究は始まったばかりであり,まだ多くの点が不明のまま残されている.たとえば「LPIの最も主要な生理機能は何なのか?」,この問いに関しては,さまざまな機能が提唱されているので,近い将来,明らかになるであろう.また,「LPIはどのようなときに産生され放出されるのか?」「LPIの産生酵素はDDHD1/PA-PLA1だけなのか?」という問いにも正確に答えられていない.「LPIの受容体はGPR55だけか?」という問いは重要なテーマで,他のリゾリン脂質メディエーターの受容体が複数存在することを考えると,必ず他の受容体が存在することが考えられる.これらの問いに答えることは,脂質メディエーターとしてのLPIとその受容体の全容を明らかにすることにつながると思われる.遠くない将来に解明されることを期待したい.
引用文献References
1) Devane, W.A., Dysarz, F.A. III, Johnson, M.R., Melvin, L.S., & Howlett, A.C. (1988) Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol., 34, 605–613.
2) Matsuda, L.A., Lolait, S.J., Brownstein, M.J., Young, A.C., & Bonner, T.I. (1990) Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature, 346, 561–564.
3) Munro, S., Thomas, K.L., & Abu-Shaar, M. (1993) Molecular characterization of a peripheral receptor for cannabinoids. Nature, 365, 61–65.
4) Devane, W.A., Hanus, L., Breuer, A., Pertwee, R.G., Stevenson, L.A., Griffin, G., Gibson, D., Mandelbaum, A., Etinger, A., & Mechoulam, R. (1992) Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science, 258, 1946–1949.
5) Sugiura, T., Kondo, S., Sukagawa, A., Nakane, S., Shinoda, A., Itoh, K., Yamashita, A., & Waku, K. (1995) 2-Arachidonoylglycerol:a possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun., 215, 89–97.
6) Mechoulam, R., Ben-Shabat, S., Hanus, L., Ligumsky, M., Kaminski, N.E., Schatz, A.R., Gopher, A., Almog, S., Martin, B.R., Compton, D.R., et al. (1995) Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol., 50, 83–90.
7) Sugiura, T., Kodaka, T., Kondo, S., Tonegawa, T., Nakane, S., Kishimoto, S., Yamashita, A., & Waku, K. (1996) 2-Arachidonoylglycerol, a putative endogenous cannabinoid receptor ligand, induces rapid, transient elevation of intracellular free Ca2+ in neuroblastoma x glioma hybrid NG108-15 cells. Biochem. Biophys. Res. Commun., 229, 58–64.
8) Sugiura, T., Kodaka, T., Nakane, S., Miyashita, T., Kondo, S., Suhara, Y., Takayama, H., Waku, K., Seki, C., Baba, N., et al. (1999) Evidence that the cannabinoid CB1 receptor is a 2-arachidonoylglycerol receptor. Structure–activity relationship of 2-arachidonoylglycerol, ether-linked analogues, and related compounds. J. Biol. Chem., 274, 2794–2801.
9) Sugiura, T., Kondo, S., Sukagawa, A., Tonegawa, T., Nakane, S., Yamashita, A., & Waku, K. (1996) Enzymatic synthesis of anandamide, an endogenous cannabinoid receptor ligand, through N-acylphosphatidylethanolamine pathway in testis:involvement of Ca2+-dependent transacylase and phosphodiesterase activities. Biochem. Biophys. Res. Commun., 218, 113–117.
10) Sugiura, T., Kondo, S., Sukagawa, A., Tonegawa, T., Nakane, S., Yamashita, A., Ishima, Y., & Waku, K. (1996) Transacylase-mediated and phosphodiesterase-mediated synthesis of N-arachidonoylethanolamine, an endogenous cannabinoid-receptor ligand, in rat brain microsomes. Comparison with synthesis from free arachidonic acid and ethanolamine. Eur. J. Biochem., 240, 53–62.
11) Sugiura, T., Kondo, S., Sukagawa, A., Tonegawa, T., Nakane, S., Yamashita, A., & Waku, K. (1996) N-arachidonoylethanolamine (anandamide), an endogenous cannabinoid receptor ligand, and related lipid molecules in the nervous tissues. J. Lipid Mediat. Cell Signal., 14, 51–56.
12) Sugiura, T., Yoshinaga, N., Kondo, S., Waku, K., & Ishima, Y. (2000) Generation of 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand, in picrotoxinin-administered rat brain. Biochem. Biophys. Res. Commun., 271, 654–658.
13) Maejima, T., Oka, S., Hashimotodani, Y., Ohno-Shosaku, T., Aiba, A., Wu, D., Waku, K., Sugiura, T., & Kano, M. (2005) Synaptically driven endocannabinoid release requires Ca2+-assisted metabotropic glutamate receptor subtype 1 to phospholipase Cβ4 signaling cascade in the cerebellum. J. Neurosci., 25, 6826–6835.
14) Oka, S., Arai, S., Waku, K., Tokumura, A., & Sugiura, T. (2007) Depolarization-induced rapid generation of 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand, in rat brain synaptosomes. J. Biochem., 141, 687–697.
15) Oka, S., Yanagimoto, S., Ikeda, S., Gokoh, M., Kishimoto, S., Waku, K., Ishima, Y., & Sugiura, T. (2005) Evidence for the involvement of the cannabinoid CB2 receptor and its endogenous ligand 2-arachidonoylglycerol in 12-O-tetradecanoylphorbol-13-acetate-induced acute inflammation in mouse ear. J. Biol. Chem., 280, 18488–18497.
16) Kishimoto, S., Muramatsu, M., Gokoh, M., Oka, S., Waku, K., & Sugiura, T. (2005) Endogenous cannabinoid receptor ligand induces the migration of human natural killer cells. J. Biochem., 137, 217–223.
17) Gokoh, M., Kishimoto, S., Oka, S., Metani, Y., & Sugiura, T. (2000) 2-Arachidonoylglycerol, an endogenous cannabinoid receptor ligand, enhances the adhesion of HL-60 cells differentiated into macrophage-like cells and human peripheral blood monocytes. FEBS Lett., 579, 6473–6478.
18) Oka, S., Wakui, J., Gokoh, M., Kishimoto, S., & Sugiura, T. (2006) Suppression by WIN55212-2, a cannabinoid receptor agonist, of inflammatory reactions in mouse ear:Interference with the actions of an endogenous ligand, 2-arachidonoylglycerol. Eur. J. Pharmacol., 538, 154–162.
19) Brown, A.J. & Wise, A. (2001) GlaxoSmithKline. WO 0186305.
20) Drmota, T., Greasley, P., & Groblewski, T. (2004) Astrazeneca. WO 2004074844.
21) Sawzdargo, M., Nguyen, T., Lee, D.K., Lynch, K.R., Cheng, R., Heng, H.H., George, S.R., & O’Dowd, B.F. (1999) Identification and cloning of three novel human G protein-coupled receptor genes GPR52, PsiGPR53 and GPR55:GPR55 is extensively expressed in human brain. Brain Res. Mol. Brain Res., 64, 193–198.
22) Oka, S., Nakajima, K., Yamashita, A., Kishimoto, S., & Sugiura, T. (2007) Identification of GPR55 as a lysophosphatidylinositol receptor. Biochem. Biophys. Res. Commun., 362, 928–934.
23) Johns, D.G., Behm, D.J., Walker, D.J., Ao, Z., Shapland, E.M., Daniels, D.A., Riddick, M., Dowell, S., Staton, P.C., Green, P., et al. (2007) The novel endocannabinoid receptor GPR55 is activated by atypical cannabinoids but does not mediate their vasodilator effects. Br. J. Pharmacol., 152, 825–831.
24) Whyte, L.S., Ryberg, E., Sims, N.A., Ridge, S.A., Mackie, K., Greasley, P.J., Ross, R.A., & Rogers, M.J. (2009) The putative cannabinoid receptor GPR55 affects osteoclast function in vitro and bone mass in vivo. Proc. Natl. Acad. Sci. USA, 106, 16511–16516.
25) Oka, S., Toshida, T., Maruyama, K., Nakajima, K., Yamashita, A., & Sugiura, T. (2009) 2-Arachidonoyl-sn-glycero-3-phosphoinositol:a possible natural ligand for GPR55. J. Biochem., 145, 13–20.
26) Henstridge, C.M., Balenga, N.A., Schröder, R., Kargl, J.K., Platzer, W., Martini, L., Arthur, S., Penman, J., Whistler, J.L., Kostenis, E., et al. (2010) GPR55 ligands promote receptor coupling to multiple signalling pathways. Br. J. Pharmacol., 160, 604–614.
27) Kapur, A., Zhao, P., Sharir, H., Bai, Y., Caron, M.G., Barak, L.S., & Abood, M.E. (2009) Atypical responsiveness of the orphan receptor GPR55 to cannabinoid ligands. J. Biol. Chem., 284, 29817–29827.
28) Yin, H., Chu, A., Li, W., Wang, B., Shelton, F., Otero, F., Nguyen, D.G., Caldwell, J.S., & Chen, Y.A. (2009) Lipid G protein-coupled receptor ligand identification using beta-arrestin PathHunter assay. J. Biol. Chem., 284, 12328–12338.
29) Oka, S., Ota, R., Shima, M., Yamashita, A., & Sugiura, T. (2010) GPR35 is a novel lysophosphatidic acid receptor. Biochem. Biophys. Res. Commun., 395, 232–237.
30) Lauckner, J.E., Jensen, J.B., Chen, H.Y., Lu, H.C., Hille, B., & Mackie, K. (2008) GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA, 105, 2699–2704.
31) Henstridge, C.M., Balenga, N.A., Ford, L.A., Ross, R.A., Waldhoer, M., & Irving, A.J. (2009) The GPR55 ligand L-alpha-lysophosphatidylinositol promotes RhoA-dependent Ca2+ signaling and NFAT activation. FASEB J., 23, 183–193.
32) Guy, A.T., Nagatsuka, Y., Ooashi, N., Inoue, M., Nakata, A., Greimel, P., Inoue, A., Nabetani, T., Murayama, A., Ohta, K., et al. (2015) Glycerophospholipid regulation of modality-specific sensory axon guidance in the spinal cord. Science, 349, 974–977.
33) Oka, S., Kimura, S., Toshida, T., Ota, R., Yamashita, A., & Sugiura, T. (2010) Lysophosphatidylinositol induces rapid phosphorylation of p38 mitogen-activated protein kinase and activating transcription factor 2 in HEK293 cells expressing GPR55 and IM-9 lymphoblastoid cells. J. Biochem., 147, 671–678.
34) Ryberg, E., Larsson, N., Sjögren, S., Hjorth, S., Hermansson, N.O., Leonova, J., Elebring, T., Nilsson, K., Drmota, T., & Greasley, P.J. (2007) The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol., 152, 1092–1101.
35) Staton, P.C., Hatcher, J.P., Walker, D.J., Morrison, A.D., Shapland, E.M., Hughes, J.P., Chong, E., Mander, P.K., Green, P.J., Billinton, A., et al. (2008) The putative cannabinoid receptor GPR55 plays a role in mechanical hyperalgesia associated with inflammatory and neuropathic pain. Pain, 139, 225–236.
36) Pietr, M., Kozela, E., Levy, R., Rimmerman, N., Lin, Y.H., Stella, N., Vogel, Z., & Juknat, A. (2009) Differential changes in GPR55 during microglial cell activation. FEBS Lett., 583, 2071–2076.
37) Deliu, E., Sperow, M., Console-Bram, L., Carter, R.L., Tilley, D.G., Kalamarides, D.J., Kirby, L.G., Brailoiu, G.C., Brailoiu, E., Benamar, K., et al. (2015) The lysophosphatidylinositol receptor GPR55 modulates pain perception in the periaqueductal gray. Mol. Pharmacol., 88, 265–272.
38) Obara, Y., Ueno, S., Yanagihata, Y., & Nakahata, N. (2011) Lysophosphatidylinositol causes neurite retraction via GPR55, G13 and RhoA in PC12 cells. PLoS One, 6, e24284.
39) Cherif, H., Argaw, A., Cécyre, B., Bouchard, A., Gagnon, J., Javadi, P., Desgent, S., Mackie, K., & Bouchard, J.F. (2015) Role of GPR55 during axon growth and target innervation. eNeuro, 2, pii: ENEURO.0011–15.2015.
40) Sylantyev, S., Jensen, T.P., Ross, R.A., & Rusakov, D.A. (2013) Cannabinoid- and lysophosphatidylinositol-sensitive receptor GPR55 boosts neurotransmitter release at central synapses. Proc. Natl. Acad. Sci. USA, 110, 5193–5198.
41) Kramar, C., Loureiro, M., Renard, J., & Laviolette, S.R. (2017) Palmitoylethanolamide modulates GPR55 receptor signaling in the ventral hippocampus to regulate mesolimbic dopamine activity, social interaction, and memory processing. Cannabis Cannabinoid Res., 2, 8–20.
42) Musella, A., Fresegna, D., Rizzo, F.R., Gentile, A., Bullitta, S., De Vito, F., Guadalupi, L., Centonze, D., & Mandolesi, G. (2017) A novel crosstalk within the endocannabinoid system controls GABA transmission in the striatum. Sci. Rep., 7, 7363.
43) Ishiguro, H., Onaivi, E.S., Horiuchi, Y., Imai, K., Komaki, G., Ishikawa, T., Suzuki, M., Watanabe, Y., Ando, T., Higuchi, S., et al. (2011) Functional polymorphism in the GPR55 gene is associated with anorexia nervosa. Synapse, 65, 103–108.
44) Kaplan, J.S., Stella, N., Catterall, W.A., & Westenbroek, R.E. (2017) Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc. Natl. Acad. Sci. USA, 114, 11229–11234.
45) Celorrio, M., Rojo-Bustamante, E., Fernández-Suárez, D., Sáez, E., Estella-Hermoso de Mendoza, A., Müller, C.E., Ramírez, M.J., Oyarzábal, J., Franco, R., & Aymerich, M.S. (2017) GPR55:A therapeutic target for Parkinson’s disease? Neuropharmacology, 125, 319–332.
46) Falasca, M. & Corda, D. (1994) Elevated levels and mitogenic activity of lysophosphatidylinositol in k-ras-transformed epithelial cells. Eur. J. Biochem., 221, 383–389.
47) Falasca, M., Silletta, M.G., Carvelli, S., Di Francesco, A.L., Fusco, A., Ramakrishna, V., & Corda, D. (1995) Signalling pathways involved in the mitogenic action of lysophosphatidylinositol. Oncogene, 10, 2113–2124.
48) Falasca, M., Iurisci, C., Carvelli, A., Sacchetti, A., & Corda, D. (1998) Release of the mitogen lysophosphatidylinositol from H-Ras-transformed fibroblasts;a possible mechanism of autocrine control of cell proliferation. Oncogene, 16, 2357–2365.
49) Hu, G., Ren, G., & Shi, Y. (2011) The putative cannabinoid receptor GPR55 promotes cancer cell proliferation. Oncogene, 30, 139–141.
50) Piñeiro, R., Maffucci, T., & Falasca, M. (2011) The putative cannabinoid receptor GPR55 defines a novel autocrine loop in cancer cell proliferation. Oncogene, 30, 142–152.
51) Andradas, C., Caffarel, M.M., Pérez-Gómez, E., Salazar, M., Lorente, M., Velasco, G., Guzmán, M., & Sánchez, C. (2011) The orphan G proteincoupled receptor GPR55 promotes cancer cell proliferation via ERK. Oncogene, 30, 245–252.
52) Ford, L.A., Roelofs, A.J., Anavi-Goffer, S., Mowat, L., Simpson, D.G., Irving, A.J., Rogers, M.J., Rajnicek, A.M., & Ross, R.A. (2010) A role for L-alpha-lysophosphatidylinositol and GPR55 in the modulation of migration, orientation and polarization of human breast cancer cells. Br. J. Pharmacol., 160, 762–771.
53) Xiao, Y.J., Schwartz, B., Washington, M., Kennedy, A., Webster, K., Belinson, J., & Xu, Y. (2001) Electrospray ionization mass spectrometry analysis of lysophospholipids in human ascitic fluids:comparison of the lysophospholipid contents in malignant vs nonmalignant ascitic fluids. Anal. Biochem., 290, 302–313.
54) Xu, Y., Xiao, Y.J., Baudhuin, L.M., & Schwartz, B.M. (2001) The role and clinical applications of bioactive lysolipids in ovarian cancer. J. Soc. Gynecol. Investig., 8, 1–13.
55) Murph, M., Tanaka, T., Pang, J., Felix, E., Liu, S., Trost, R., Godwin, A.K., Newman, R., & Mills, G. (2007) Liquid chromatography mass spectrometry for quantifying plasma lysophospholipids:potential biomarkers for cancer diagnosis. Methods Enzymol., 433, 1–25.
56) Sutphen, R., Xu, Y., Wilbanks, G.D., Fiorica, J., Grendys, E.C. Jr., LaPolla, J.P., Arango, H., Hoffman, M.S., Martino, M., Wakeley, K., et al. (2004) Lysophospholipids are potential biomarkers of ovarian cancer. Cancer Epidemiol. Biomarkers Prev., 13, 1185–1191.
57) Metz, S.A. (1986) Lysophosphatidylinositol, but not lysophosphatidic acid, stimulates insulin release. A possible role for phospholipase A2 but not de novo synthesis of lysophospholipid in pancreatic islet function. Biochem. Biophys. Res. Commun., 138, 720–727.
58) Metz, S.A. (1998) Mobilization of cellular Ca2+ by lysophospholipids in rat islets of Langerhans. Biochim. Biophys. Acta, 968, 239–252.
59) Moreno-Navarrete, J.M., Catalán, V., Whyte, L., Díaz-Arteaga, A., Vázquez-Martínez, R., Rotellar, F., Guzmán, R., Gómez-Ambrosi, J., Pulido, M.R., Russell, W.R., et al. (2012) The L-α-lysophosphatidylinositol/GPR55 system and its potential role in human obesity. Diabetes, 61, 281–291.
60) Imbernon, M., Whyte, L., Diaz-Arteaga, A., Russell, W.R., Moreno, N.R., Vazquez, M.J., Gonzalez, C.R., Díaz-Ruiz, A., Lopez, M., Malagón, M.M., et al. (2014) Regulation of GPR55 in rat white adipose tissue and serum LPI by nutritional status, gestation, gender and pituitary factors. Mol. Cell. Endocrinol., 383, 159–169.
61) Yamashita, A., Hayashi, Y., Nemoto-Sasaki, Y., Ito, M., Oka, S., Tanikawa, T., Waku, K., & Sugiura, T. (2014) Acyltransferases and transacylases that determine the fatty acid composition of glycerolipids and the metabolism of bioactive lipid mediators in mammalian cells and model organisms. Prog. Lipid Res., 53, 18–81.
62) Yamashita, A., Oka, S., Tanikawa, T., Hayashi, Y., Nemoto-Sasaki, Y., & Sugiura, T. (2013) The actions and metabolism of lysophosphatidylinositol, an endogenous agonist for GPR55. Prostaglandins Other Lipid Mediat., 107, 103–116.
63) Higgs, H.N., Han, M.H., Johnson, G.E., & Glomset, J.A. (1998) Cloning of a phosphatidic acid-preferring phospholipase A1 from bovine testis. J. Biol. Chem., 273, 5468–5477.
64) Yamashita, A., Kumazawa, T., Koga, H., Suzuki, N., Oka, S., & Sugiura, T. (2010) Generation of lysophosphatidylinositol by DDHD domain containing 1 (DDHD1):Possible involvement of phospholipase D/phosphatidic acid in the activation of DDHD1. Biochim. Biophys. Acta, 1801, 711–720.
65) Tesson, C., Nawara, M., Salih, M.A., Rossignol, R., Zaki, M.S., Al Balwi, M., Schule, R., Mignot, C., Obre, E., Bouhouche, A., et al. (2012) Alteration of fatty-acid-metabolizing enzymes affects mitochondrial form and function in hereditary spastic paraplegia. Am. J. Hum. Genet., 91, 1051–1064.
66) Baba, T., Kashiwagi, Y., Arimitsu, N., Kogure, T., Edo, A., Maruyama, T., Nakao, K., Nakanishi, H., Kinoshita, M., Frohman, M.A., et al. (2014) Phosphatidic acid (PA)-preferring phospholipase A1 regulates mitochondrial dynamics. J. Biol. Chem., 289, 11497–11511.
67) Gonzalez, M., Nampoothiri, S., Kornblum, C., Oteyza, A.C., Walter, J., Konidari, I., Hulme, W., Speziani, F., Schöls, L., Züchner, S., et al. (2013) Mutations in phospholipase DDHD2 cause autosomal recessive hereditary spastic paraplegia (SPG54). Eur. J. Hum. Genet., 21, 1214–1218.
68) Blondeau, N., Lauritzen, I., Widmann, C., Lazdunski, M., & Heurteaux, C. (2002) A potent protective role of lysophospholipids against global cerebral ischemia and glutamate excitotoxicity in neuronal cultures. J. Cereb. Blood Flow Metab., 22, 821–834.
69) Savitz, S.I., Dhallu, M.S., Malhotra, S., Mammis, A., Ocava, L.C., Rosenbaum, P.S., & Rosenbaum, D.M. (2006) EDG receptors as a potential therapeutic target in retinal ischemia-reperfusion injury. Brain Res., 1118, 168–175.
70) Mariggiò, S., Sebastià, J., Filippi, B.M., Iurisci, C., Volonté, C., Amadio, S., De Falco, V., Santoro, M., & Corda, D. (2006) A novel pathway of cell growth regulation mediated by a PLA2α-derived phosphoinositide metabolite. FASEB J., 20, 2567–2569.
71) Ueda, H., Kobayashi, T., Kishimoto, M., Tsutsumi, T., & Okuyama, H. (1993) A possible pathway of phosphoinositide metabolism through EDTA-insensitive phospholipase A1 followed by lysophosphoinositide-specific phospholipase C in rat brain. J. Neurochem., 61, 1874–1881.
72) Tsutsumi, T., Kobayashi, T., Ueda, H., Yamauchi, E., Watanabe, S., & Okuyama, H. (1994) Lysophosphoinositide-specific phospholipase C in rat brain synaptic plasma membranes. Neurochem. Res., 19, 399–406.
73) Aoki, J., Taira, A., Takanezawa, Y., Kishi, Y., Hama, K., Kishimoto, T., Mizuno, K., Saku, K., Taguchi, R., & Arai, H. (2002) Serum lysophosphatidic acid is produced through diverse phospholipase pathways. J. Biol. Chem., 277, 48737–48744.
74) Nakane, S., Oka, S., Arai, S., Waku, K., Ishima, Y., Tokumura, A., & Sugiura, T. (2002) 2-Arachidonoyl-sn-glycero-3-phosphate, an arachidonic acid-containing lysophosphatidic acid:occurrence and rapid enzymatic conversion to 2-arachidonoyl-sn-glycerol, a cannabinoid receptor ligand, in rat brain. Arch. Biochem. Biophys., 402, 51–58.