環状ホスファチジン酸の生理活性Insights of cyclic phosphatidic acid function
お茶の水女子大学ヒューマンライフイノベーション研究所Institute for Human Life Innovation ◇ 〒112–8610 東京都文京区大塚2–1–1 ◇ 2–1–1 Ohtsuka, Bunkyo-ku, Tokyo 112–8610, Japan
発行日:2018年12月25日Published: December 25, 2018
環状ホスファチジン酸(cyclic phosphatidic acid:cPA)は,我々が1985年に真性(真正)粘菌Physarum polycephalumから初めて単離し,構造や生理活性に関して1992年に論文発表した生理活性脂質である.その後の研究によって,cPAが粘菌のような特殊な生物のみに存在する物質ではなく,多くの生物種に普遍的に存在する生理活性脂質であることがわかってきた.cPAは,リゾホスファチジン酸(lysophosphatidic acid:LPA)と構造が類似しているが,グリセロール骨格のsn-2位とsn-3位に環状リン酸基を持つ非常に特徴的な構造を有している.本稿では,cPAの多様な生理活性を紹介し,今後の課題について概説する.
© 2018 公益社団法人日本生化学会© 2018 The Japanese Biochemical Society
我々は,細胞分裂や細胞分化のモデル生物として注目されていた真性(真正)粘菌Physarum polycephalumを材料として,抗がん作用を持つ物質を探索し,Physarumの単相体・ミクソアメーバの脂溶性画分中に,細胞増殖を顕著に抑制する活性を見いだした.それがDNAポリメラーゼα活性を抑制することも明らかになり,学会で発表したのが1985年のことだった.我々は興味深い生理活性物質の存在を確信して,その分離・精製を進め,18 cmシャーレ約2700枚のミクソアメーバからようやく2 mgの試料を得た.構造決定を行った結果,その生理活性物質は脂肪酸鎖にシクロヘキサン構造を持ち,リゾホスファチジン酸(lysophosphatidic acid:LPA,図1A)と構造が類似しているが,グリセロール骨格のsn-2位とsn-3位に環状リン酸基を持つ非常に特徴的な構造を有する脂質であることがわかった(図1B).ちょうどそのころ,1989年にLPAの細胞増殖促進作用に関する興味深い論文がCellに掲載され1),その構造の類似性から,我々の物質をPhysarum由来のLPA類似物質という意味で“PHYLPA”と名づけ,1992年にJ. Biol. Chem.に発表した2).その後,PHYLPA類似物質が哺乳類の血液や脳にも存在することを見いだし,これが粘菌のような特殊な生物のみに含まれる物質ではないことを明らかにした3).PHYLPAを発表した論文と同じ号に,Gabor Tigyi氏(テネシー大学)がLPAの生理作用に関する興味深い論文を発表しており,その縁で彼らとの共同研究も始まった.そして,彼の提案で環状リン酸基構造を持つ脂質を“環状ホスファチジン酸(cyclic phosphatidic acid:cPA)”と総称することとした.以後,この名称が一般的に使用されている(図1C).
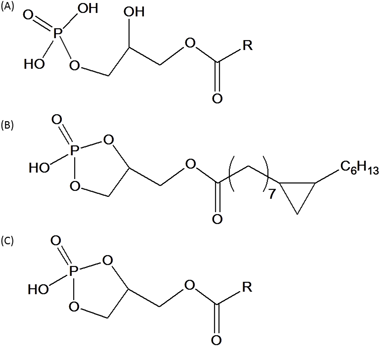
最初に見つかったPhysarum由来のcPAは,脂肪酸鎖にシクロヘキサン構造を持つユニークな微量物質であったが,その後,ヒトの血清中にも,一般的な脂肪酸側鎖を持つcPAが微量存在していることがわかった.cPAは血清アルブミンに抱合された形で存在すると考えられることから,市販されているヒト血清アルブミン2 kg(血清約50 Lに相当)を材料としてcPAを抽出し,200 µgのcPAを得て分析を行ったところ,cPA16 : 0(16 : 0の脂肪酸鎖を持つcPA)が最も多く,他にcPA14 : 0, cPA18 : 0が検出された3).存在量は,血清に換算するとおおむねcPA16 : 0が100 nM程度で,この量はLPAの約1/10である.なお,ShanらがESIをイオン源としたLC/MS/MSを用いた解析で,ヒト血清中にcPA16 : 0と18 : 1が10~15 nM程度存在し,cPA16 : 0の存在量は卵巣がん患者の手術前では健常者よりも少なく,手術後に健常者と同等量まで増加することを報告している4).我々はLC-MS/MSを用いてマウスの脳,心臓,肺,肝臓,脾臓,腎臓,血漿中にcPA16 : 0, 18 : 0, 18 : 1, 18 : 2が含まれていることを明らかにした5).そのうち,cPA16 : 0が最も多く,脳6.27±1.01,心臓1.08±0.51,肺2.07±1.04,肝臓5.83±2.82,脾臓0.23±0.11,腎臓4.27±1.24 nmol/g-wet tissue,血漿0.34±0.08 nmol/mLであった.これらの実験では,臓器回収の際に,酵素反応や分解によりcPAの量が変動してしまうことを懸念し,血液を潅流せずに臓器を回収している.結果として脳や肝臓にcPAが多く含まれていることが示されたが,正確な値を得るためには,それぞれの臓器中に含まれる血液量がcPAの量として反映されていることを考慮することが必要である.
cPAの量の変動は多発性硬化症動物モデルであるクプリゾンマウスにおいても観察され,ミエリンが脱落しているクプリゾンマウスでは脳のcPA量が減少することが示された5).さらに我々は,ラットの血清中にはcPA16 : 0, 18 : 0, 18 : 1, 18 : 2に加えて,cPA20 : 4が存在していることを明らかにした6).存在量はcPA16 : 0が12.01±0.72 nMと最も多く,次いで18 : 0, 18 : 2, 20 : 4, 18 : 1であった.LPAに結合している脂肪酸鎖は不飽和脂肪酸が多いが,cPAでは飽和脂肪酸の存在比が高く,血清中のリゾホスファチジルコリン(lysophosphatidylcholine:LPC)の側鎖の存在比と相関があった(表1)6).このことは,血清中のcPAの由来が主にLPCであることを示唆する.
| cPA [nM] | LPA [nM] | LPC [µM] | |
|---|---|---|---|
| 16 : 0 | 12.01±0.72 | 115.34±13.04 | 210.68±6.85 |
| 16 : 1 | 検出限界以下 | 42.43±6.04 | 13.40±0.91 |
| 18 : 0 | 11.25±0.76 | 84.04±9.75 | 135.31±8.80 |
| 18 : 1 | 4.42±0.15 | 167.99±15.91 | 52.87±1.33 |
| 18 : 2 | 7.63±1.04 | 980.36±62.73 | 83.87±5.55 |
| 20 : 3 | 検出限界以下 | 84.70±4.13 | 5.69±0.34 |
| 20 : 4 | 6.05±0.40 | 1303.74±80.85 | 46.12±2.37 |
近年,我々以外のグループが生体内でのcPA量を測定し,病態との相関を報告している.たとえば,Calaらは,コロンビアのヒスパニックの女性の乳がん患者では血漿中のcPA18 : 0の量が減少していることを報告した7).また,Woodらは,レプトスピラに感染,またはレプトスピラワクチンを接種したウマではcPAの量が増加していることを報告している8).これらの報告は,cPAが生体内でさまざまな状況において生理活性脂質としてその存在量が調節され,生理的機能を有していることを示唆するものである.
現在,LC-MS/MSの技術が進み,高感度で精度の高い測定が可能となった.上述のcPAの測定もLC-MS/MSで行われたものがほとんどである.高感度分析では,サンプル調製におけるハンドリング精度が測定に影響を与えることから,特に効率・精度のよいcPAの抽出法の確立が必要である.Shanらは,塩酸を用いた酸性Bligh-Dyer法では,塩酸の濃度を上げると血清中のcPAの存在量が増加することを指摘していた4).LPAなどの酸性脂質は,塩酸を用いた酸性Bligh-Dyer法により抽出されることが多いが,我々は塩酸を用いた酸性Bligh-Dyer法ではLPCがcPAへと(LPAにもわずかに)変換されてしまう現象を見つけた6).
我々は,LPCからcPAが抽出時に人工的に産生されることなく,かつ効率よくcPAやLPAを抽出する方法として,種々の条件を検討した.その結果,クエン酸を用いたBligh-Dyer法では,LPCからcPAやLPAへの変換がなく,cPA, LPAの抽出効率も高いことが示された6).また,酸性メタノールを用いた,除タンパクを目的とした精製でも,LPCからcPAやLPAが産生されることはなかった.どのような方法で抽出・精製するかは,分析に用いるカラムとの相性や分析に用いることが可能な試料の量などを考慮して選択しなければならないが,条件によって組織中にcPAの10,000倍程度大量に含まれるLPCから人工的にcPAが産生されてしまわないように,また抽出・精製過程において微量の成分を失わないように細心の注意を払わなければならない.
精製されたcPAの安定性については,粉末の場合,−80°C保存で数年間の長期保存が可能である.中性有機溶媒中での保存の場合は−80°Cで1年以上の長期保存が可能であり,4°Cでは少なくとも数日間は保存可能である.しかし,酸性有機溶媒中では水含有量により加水分解が進むことを確認している(data not shown).
cPAの生合成を触媒する酵素の候補として,ホスホリパーゼD(phospholipase D:PLD)類似の酵素の存在が予想された.PLDはリン脂質を加水分解すると同時に,リン酸基転移反応を触媒する酵素であることから,LPCに働いて,主な反応産物であるLPAとともに,cPAを生じると考えられる.1996年にFriedmanらは,Streptomyces chromofuscusのPLDによってLPCからLPAの他にcPAが合成されることを報告した9).その後,我々はさまざまな生物種由来のPLDによるLPAとcPAの合成量を測定したところ,Actinomadura sp. NO362由来のPLDがLPAよりもcPAを主な生成物として産生することが明らかになった10).また,動物の血清中にもcPAを産生する酵素が存在することを見いだし,この酵素を精製してアミノ酸配列を調べたところ,これが血清中のオートタキシン(autotaxin:ATX)と一致することがわかった(図2)11).ATXは以前,細胞のがん化やがんの増悪化と関連しているタンパク質として報告されたが,その後,ATXがLPA合成酵素であること,またがん化やがんの増悪化を引き起こす機能分子はATXによって産生されるLPAであることが明らかにされていた12).我々は,in vitroでの反応溶液を2層に分けた(体積の0.4倍量のジエチルエーテル,1 M NaClを添加)反応条件下では,ATXによるLPCからのcPA合成量が増加することを示した11).これらの条件は,生体内での膜の表面を模したものであると考えているが,実際にそのようなことが生体内で起こっているか否かについては今後の検証が必要である.LPCミセルも膜の表面を模したものと考えられ,Lajoieらは,250 µMのLPCを基質としたときにLoxosceles arizonicaのPLDによる主な反応産物がLPAではなくcPAであることを報告している13).Loxosceles由来のPLDは,哺乳類の細胞表面に結合し,LPCやスフィンゴミエリン(sphingomyelin:SM)に作用する.これらのことから,細胞表面において,PLDのリン酸基転移活性が加水分解活性にまさって,LPAではなくcPAを主な生成物として産生する微小環境をつくるのかもしれない.また,興味深いことにLajoieらは,Loxosceles由来のPLDがグリセロホスホジエステラーゼ(リン脂質から2本の脂肪酸鎖が切断されたグリセロホスホジエステルを分解する酵素)と進化の歴史を共有していると述べている13).グリセロホスホジエステラーゼの中には,環状ホスホジエステルを最終産物として産生するものもあり,cPAを主な最終産物として生成するPLDあるいはグリセロホスホジエステラーゼが哺乳類にも存在する可能性は高いと考えられる.実際に,マウスのグリセロホスホジエステラーゼ(CDE4, 7)がLPAを産生することが報告されており14),これらの酵素にもcPA合成活性がある可能性が考えられる.
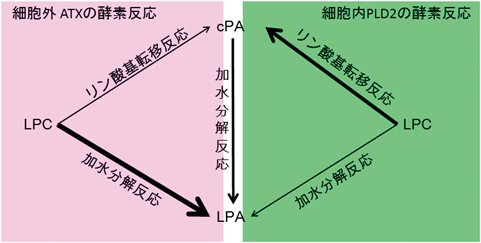
矢印の太さは反応の起こりやすさを示し,ATXのcPA : LPA合成量比は条件によって1 : 4~1 : 10, PLD2のcPA : LPA合成量比は約3 : 1である.
我々は,哺乳類の細胞膜に局在するPLD2が,刺激に応じてcPAを主な反応産物として産生することを見いだしており(図2)15),これらの結果から,cPAがATXによってLPAと同時に細胞外で産生される経路と,刺激に応じてPLD2の主要な反応産物として細胞内で合成される経路があると考えている.
cPAは生体内にきわめて微量しか含まれていないため,生理作用を明らかにする実験に使用できるだけの量を生物体から精製して得ることは困難であった.現在は,cPAの合成品が市販されているが,我々がcPAの生理活性の研究に用いたcPA試料は小林進教授(当時:東京大学薬学部,後に東京理科大学薬学部)に化学合成していただいたものである16).小林教授のご助力によって我々の研究は大きく進展した.
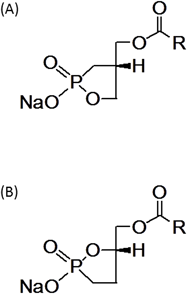
また,生体内において,cPAのリン酸エステルの分解による不活性化,加水分解によるLPAへの変化などが懸念されるため,分解されにくいさまざまな誘導体を小林教授に合成していただいたが,その中で,sn-2あるいはsn-3位の酸素原子をメチレン基に置換した誘導体,すなわち2-カルバcPA(2-carba-cPA:2ccPA),3-カルバcPA(3-carba-cPA:3ccPA)に,高い活性があることが示された(図3)17–20).以下にcPAの生理活性について紹介するが,誘導体を用いた研究結果についてもまとめて記載することとする.また現在までにわかっている経路とともに図4にcPAの細胞内での生合成とcPA, 2ccPAの生理活性を示す.
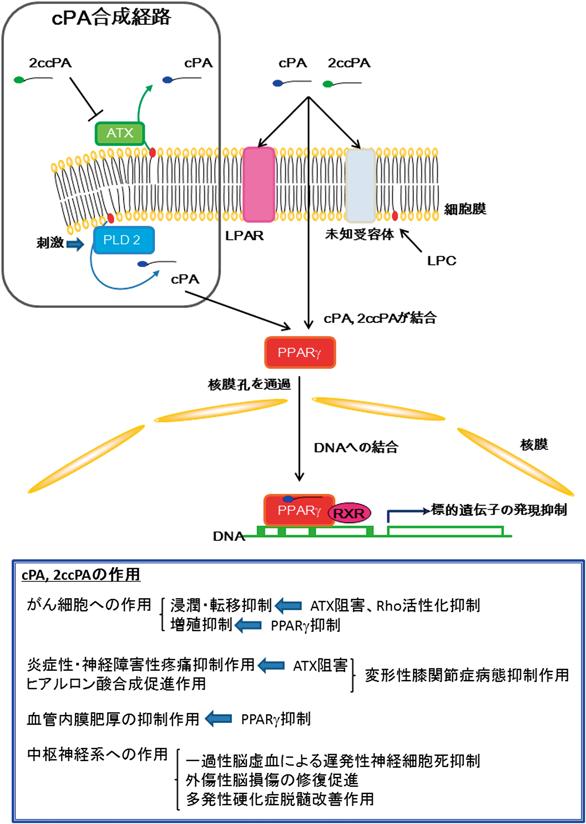
ATX:オートタキシン,cPA:環状ホスファチジン酸,LPAR:リゾホスファチジン酸受容体,LPC:リゾホスファチジルコリン,PLD2:ホスホリパーゼD2,PPARγ:ペルオキシソーム増殖因子活性化受容体γ,RXR:レチノイン酸X受容体,2ccPA:2−カルバ環状ホスファチジン酸.
細胞のがん化やがんの増悪化の作用因子がLPAであること21)を明らかにした大阪府立成人病センター研究所の明渡均所長のグループから共同研究の申し込みを受け,cPAのがん細胞の浸潤・転移への作用について検討を行った.その結果,驚くことにcPAと誘導体2ccPA, 3ccPAが,種々のがん細胞の浸潤を顕著に抑制すること17–19),さらにはマウスを用いたin vivo実験で,メラノーマ細胞の肺転移を顕著に抑制することが明らかになった17, 19).また,cPAは化学発がん物質によって形成されるがん病巣からの浸潤と転移も著しく抑制した22).cPAによって,低分子量Gタンパク質Rhoの活性化が抑制されることも確認された23).一方で,cPA, 3ccPA,特に2ccPAが,LPA合成酵素であるATXの活性を顕著に抑制することも明らかになった19).現在,さまざまなATX阻害によるがんの制御が検討されており24),我々は,2ccPAはATXを阻害することで,がん細胞の浸潤・転移を誘導するLPAの産生を抑え,その結果としてがんの浸潤・転移が抑制されると考えている.
これまでの研究から得られたcPAの生理活性やLPAの作用との比較から,cPAに鎮痛作用がある可能性を考え,検討を行った.その結果,2ccPAにはモルヒネに匹敵する鎮痛作用があることが明らかになった25).さらに,当時,LPAによる痛み誘導のメカニズムを明らかにしていた長崎大学の植田弘師教授26)との共同研究で,2ccPAが神経のC繊維の興奮を抑制し,炎症性疼痛や神経障害性疼痛を抑制することを示した25).これは,2ccPAが障害を受けた部位で活性化されるATXを抑制することで,痛み促進物質であるLPAの産生を抑え,鎮痛作用を示す結果であると考えている.また,これらの研究で得られた結果から,2ccPAが神経の障害部位を治癒・改善する作用がある可能性が示された25).
変形性膝関節症は,筋力低下,加齢,肥満,怪我などのきっかけにより関節機能が低下し,軟骨の摩耗,骨の変形等を起こすことで関節の可動域が制限され,また,滑膜の炎症による痛みも伴う疾患である.我々はcPA, 2ccPAの炎症性疼痛抑制作用25)が変形性膝関節症の症状を緩和する可能性を考え,ウサギの膝関節半月板切除による変形性膝関節症モデルへの2ccPAの効果を検討した.その結果,2ccPA投与群は,vehicle投与群と比較して軟骨変性に伴う疼痛を経時的に抑制し,また,腫脹も顕著に抑制することが明らかになった.病理組織学的解析からは,2ccPA投与群では,vehicle投与群でみられた関節軟骨表層の消失とプロテオグリカン量の低下などが顕著に抑制されることが示された27).これらの結果は,2ccPAが変形性膝関節症の病態を改善する医薬品として有用であることを示唆している.
変形性膝関節症では,関節腔内中のヒアルロン酸の減少,軟骨破壊酵素であるマトリックスメタロプロテアーゼ(matrix metalloproteinase:MMP)の発現上昇に伴う軟骨破壊の亢進が観察される.そこで,2ccPAの効果の作用機序についての検討を行うために,変形性膝関節症の発症に大きな役割を担っているヒアルロン酸を合成している滑膜細胞と,MMPのいくつかの分子種(MMP-1, MMP-3, MMP-13)を発現している軟骨細胞を用いた実験を行った.その結果,滑膜細胞を用いた実験では,2ccPAが濃度ならびに時間依存的にヒアルロン酸産生量を顕著に増加させることがわかった27).次に,軟骨細胞を用いて,2ccPAがMMP-1, MMP-3, MMP-13の発現量に与える影響について検討した.その結果,軟骨細胞において,2ccPAは炎症性サイトカインIL-1βにより誘導されたMMP-1とMMP-13の遺伝子発現とタンパク質合成を有意に抑制した27).In vitroの実験から,2ccPAによる変形性膝関節症病態の改善効果は,2ccPAの滑膜細胞へのヒアルロン酸合成促進作用や炎症状態にある軟骨細胞におけるMMPの発現抑制作用が複合的に作用したためであるという可能性が示唆された27).
変形性膝関節症はQOLを著しく低下させるため,根本的治療が望まれているが,現在行われている治療は,消炎鎮痛剤や局部へのヒアルロン酸注射等の対症療法のみである.現在,我々は2ccPAを変形性膝関節症の治療薬としての開発を目指し,米国FDAから正式承認を受け,Phase 1/b試験を進めているところである(FDA IND number:124374).
他方で,皮膚は身体を構成する最大の臓器であり,また健康状態や老化の状態を外から観察できるため,皮膚を健康に保つことは心と体を健康に保つためにもきわめて重要である.我々は,2ccPAの滑膜細胞に対するヒアルロン酸合成促進作用が皮膚の線維芽細胞においてもみられるか否かを検討した.その後の化粧品開発を見据えて,天然体であるcPAを用いて検討を行った結果,cPAは皮膚の線維芽細胞に対してもヒアルロン酸合成促進作用を示し,その作用は,LPA受容体の一つであるLPAR1を介してヒアルロン酸合成酵素(HAS2)の発現を上昇させたためであることがわかった28).また,cPAはさまざまな年齢ドナー由来の線維芽細胞においてもヒアルロン酸合成促進作用を示し,興味深いことに,老化した細胞ほど促進効果は高かった29).さらにセラミド分解を抑制したり,表皮細胞どうしの結合を強化するなどの作用があることもわかって30),さまざまな安全性試験を経た後に,化粧品原料としてcPAを配合した化粧品が,複数の企業(雅Grace/SANSHO株式会社,グラヴィティレジスタンスAD/株式会社ALBION,潤Mellow/株式会社冨山房インターナショナルなど)から販売されることになった.これは我々の研究成果が商品として結実し,社会に出ていった最初の例である.
これまでの実験から,cPAが動物の脳に比較的高濃度で存在していることが示されていた10).そこで我々は,cPAが神経に対して重要な役割を果たしている可能性を考え,海馬の初代培養細胞に添加することで,その効果を調べた.その結果,cPAが神経成長因子(nerve growth factor:NGF)と同等程度に,神経細胞の生存促進作用と神経突起の伸長促進作用を持つことがわかった31).その後,神経細胞を低酸素に曝露したときに引き起こされるアポトーシスが,cPAによって抑制されることも明らかになった32).また,神経細胞以外にも,2ccPAがアストロサイトおよびオリゴデンドロサイト前駆細胞に対して細胞増殖作用を示すことがわかった33).
これまでのin vitroでの研究成果を踏まえ,我々は,cPA, 2ccPAが,実際に脳のさまざまな疾患に対してin vivoで効果を示すか否かを,①一過性脳虚血ラット,②外傷性脳損傷マウス,③多発性硬化症マウスの3種類の動物疾患モデルを用いて検討した.
これらの結果から,脳梗塞の後遺症予防や交通事故後の脳損傷の病状改善,多発性硬化症の治療薬としてcPAや2ccPAが利用できることが期待される.
先に述べたように,cPAは細胞外からの刺激に応じてPLD2により合成誘導されることが示されている15).産生されたcPAは細胞内でセカンドメッセンジャーとして働き,核内受容体であるペルオキシソーム増殖因子活性化受容体γ(PPARγ)のアンタゴニストとして働く可能性がある15, 40).LPAが,大腸がんに関与することが示唆されているが,cPAはPPARγに働いて大腸がん細胞の増殖を阻害し,さらにアポトーシスを誘導することが確認された41).これらの結果から,生理活性脂質の生体内バランスが生体の恒常性維持にとって重要であることが示唆される.cPAによる大腸がん細胞の増殖抑制作用は,新規な大腸がん治療法につながるものと強く期待される41).
また,PPARγは,血液中に含まれるある種の脂肪酸により活性化され,標的遺伝子の発現を介して血管内膜肥厚を亢進するが,cPAは,その活性化を抑えることにより,血管内膜肥厚を有意に抑制する15).血液と接している血管内膜および内皮細胞の傷害から発生する肥厚の予防・治療戦略は動脈硬化の予防・治療においてきわめて有効であるが,内膜肥厚の抑制は,血管学における永年の研究テーマであるにもかかわらず,現状では有効な予防,治療方法は報告されていない.このことから,我々はcPAのPPARγへのはたらきを活用して,cPAをリード化合物とした血管内皮機能改善を目指した創薬を行っていくことを考えている.
残念なことに,まだ,cPA, 2ccPAの特異的な細胞膜受容体は見つかっていないが,これまでの実験から,cPAと2ccPAがLPA受容体の1~6を活性化することが示されている19, 42–44).ただし,天然体であるcPAの場合は,cPA中にわずかに含まれるLPA,あるいは細胞に添加した際に瞬間的にcPAがLPAへと変換され,変換されたLPAが受容体を活性化している可能性も否定できないため,さらなる検討が必要であろう.しかし,化学合成された2ccPAの場合は,cPAの混入もなく,加水分解されてもLPAを生じないことから,2ccPAあるいはその代謝物がLPA受容体を直接活性化していると考えてよさそうである.
cPA, 2ccPAによるLPA受容体1~6の活性化には,LPAよりも高い濃度が必要な場合が多いが,cPAとLPAが相反する作用を持つことは,きわめて興味深い10, 45, 46).これには,いくつか可能性が考えられる.①cPAに特異的な未同定の受容体がある可能性.これは,cPA特異的な活性を説明するのに都合がよいが,前に述べたように,まだ特異的受容体の発見には到達できていない.②通常,細胞は同時に複数のLPA受容体を発現しているが,LPA受容体が複数発現している中で,LPAとcPAの受容体に対する親和性に違いがある可能性.これまでに我々は,同じ細胞においてLPAはLPAR1を主に活性化するのに対し,cPAはLPAR2を主に活性化するケースがあることを観察している32).LPAとcPAのLPA受容体に対する親和性や活性化の程度の違いから,受容体の細胞内への取り込みや活性化の違いが生じていることが考えられる.時に,cPAはLPAと同様の働きをすることがあることもわかっているが28, 32),これは各々の細胞に発現しているLPA受容体の割合が関係しているのではないかと考えている.
cPAが生体中で重要な働きをする物質であることを示すためには,これが生体中で安定に合成され,代謝されることを示す,あるいはcPAに特異的な受容体,結合タンパク質を見つけることが必要である.我々は約30年の間に,cPAを発見し,次いでcPAの持つさまざまな生理活性について明らかにしてきた.この物質が生体内において,どのようにして多様な働きの制御に関わっているのかを知ることが重要な課題であるので,今後は,cPAの生体における生理学的存在意義について解明していくことが必要であると考えている.
また,cPAやcPAの誘導体である2ccPAには,さまざまな疾患の病状を改善・緩和する作用があることがわかっており,これらの活性を活かした医薬品の開発を早急に進めたいと考えている10, 15, 25, 27, 34, 37–41).2ccPA, 3ccPAに関しては構造中に不斉炭素原子があるために,R体,S体の活性の違いについて検討する必要があると考え,2ccPA, 3ccPAのR体,S体を化学合成して,それらのATX阻害,がん細胞の浸潤・抑制,痛み抑制などの生理活性について調べ,R体,S体の間に差がないことを確認している47, 48).また,これまでにも約30種類の誘導体を合成したが17–20),さらに2ccPA以外に効果の高い化合物がないか,cPA誘導体の最適化を進めている.さらには,薬剤として開発するためには生体内に安定的にcPA, 2ccPAを投与できる方法を検討しなければならないことから,現在,我々は,徐放剤49)や経口投与によるcPA, 2ccPAの投与方法の検討を進めている.これらの検討を進めることにより,生理活性脂質を起源とした薬剤の開発が日本で進むことを期待している.
本稿で紹介した研究は,お茶の水女子大学理学部およびヒューマンウェルフェアサイエンス研究教育寄附研究部門の室伏きみ子研究室に所属していた学生やスタッフ,多くの学内外の共同研究者の協力のもとで行われたものです.研究に携わっていただいた多くの皆様に深く感謝申し上げます.また,本稿を執筆するにあたり,長崎大学の塚原完先生,埼玉医科大学の吉川圭介先生,お茶の水女子大学の深澤桂子さん,芝池由樹さんにご協力いただきました.心より御礼申し上げます.
1) van Corven, E.J., Groenink, A., Jalink, K., Eichholtz, T., & Moolenaar, W.H. (1989) Lysophosphatidate-induced cell proliferation:identification and dissection of signaling pathways mediated by G proteins. Cell, 59, 45–54.
2) Murakami-Murofushi, K., Shioda, M., Kaji, K., Yoshida, S., & Murofushi, H. (1992) Inhibition of eukaryotic DNA polymerase alpha with a novel lysophosphatidic acid (PHYLPA) isolated from myxoamoebae of Physarum polycephalum. J. Biol. Chem., 267, 21512–21517.
3) Kobayashi, T., Tanaka-Ishii, R., Taguchi, R., Ikezawa, H., & Murakami-Murofushi, K. (1999) Existence of a bioactive lipid, cyclic phosphatidic acid, bound to human serum albumin. Life Sci., 65, 2185–2191.
4) Shan, L., Li, S., Jaffe, K., & Davis, L. (2008) Quantitative determination of cyclic phosphatidic acid in human serum by LC/ESI/MS/MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci., 862, 161–167.
5) Shimizu, Y., Ishikawa, M., Gotoh, M., Yamamoto, S., Iwasa, K., Yoshikawa, K., & Murakami-Murofushi, K. (2018) Quantitative determination of cyclic phosphatidic acid and its carba analog in mouse organs and plasma using LC-MS/MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci., 1076, 15–21.
6) Fukasawa, K., Nakajima, S., Gotoh, M., Tanaka, S., Murofushi, H., & Murakami-Murofushi, K. (2018) Qualitative and quantitative comparison of cyclic phosphatidic acid and its related lipid species in rat serum by LC-MS/MS using hydrophilic interaction liquid chromatography. J. Chromatogr. A, 1567, 177–184.
7) Cala, M.P., Aldana, J., Medina, J., Sanchez, J., Guio, J., Wist, J., & Meesters, R.J.W. (2018) Multiplatform plasma metabolic and lipid fingerprinting of breast cancer: A pilot control-case study in Colombian Hispanic women. PLoS One, 13, e0190958.
8) Wood, P.L., Steinman, M., Erol, E., Carter, C., Christmann, U., & Verma, A. (2018) Lipidomic analysis of immune activation in equine leptospirosis and Leptospira-vaccinated horses. PLoS One, 13, e0193424.
9) Friedman, P., Haimovitz, R., Markman, O., Roberts, M.F., & Shinitzky, M. (1996) Conversion of lysophospholipids to cyclic lysophosphatidic acid by phospholipase D. J. Biol. Chem., 271, 953–957.
10) Murakami-Murofushi, K., Uchiyama, A., Fujiwara, Y., Kobayashi, T., Kobayashi, S., Mukai, M., Murofushi, H., & Tigyi, G. (2002) Biological functions of a novel lipid mediator, cyclic phosphatidic acid. Biochim. Biophys. Acta, 1582, 1–7.
11) Tsuda, S., Okudaira, S., Moria-Ito, K., Shimamoto, C., Tanaka, M., Aoki, J., Arai, H., Murakami-Murofushi, K., & Kobayashi, T. (2006) Cyclic phosphatidic acid is produced by autotaxin in blood. J. Biol. Chem., 281, 26081–26088.
12) Umezu-Goto, M., Kishi, Y., Taira, A., Hama, K., Dohmae, N., Takio, K., Yamori, T., Mills, G.B., Inoue, K., Aoki, J., et al. (2002) Autotaxin has lysophospholipase D activity leading to tumor cell growth and motility by lysophosphatidic acid production. J. Cell Biol., 158, 227–233.
13) Lajoie, D.M., Zobel-Thropp, P.A., Kumirov, V.K., Bandarian, V., Binford, G.J., & Cordes, M.H.J. (2013) Phospholipase D toxins of brown spider venom convert lysophosphatidylcholine and sphingomyelin to cyclic phosphates. PLoS One, 8, e72372.
14) Ohshima, N., Kudo, T., Yamashita, Y., Mariggiò, S., Araki, M., Honda, A., Nagano, T., Isaji, C., Kato, N., Corda, D., et al. (2015) New members of the mammalian glycerophosphodiester phosphodiesterase family:GDE4 and GDE7 produce lysophosphatidic acid by lysophospholipase D activity. J. Biol. Chem., 290, 4260–4271.
15) Tsukahara, T., Tsukahara, R., Fujiwara, Y., Yue, J., Cheng, Y., Guo, H., Bolen, A., Zhang, C., Balazs, L., Re, F., et al. (2010) Phospholipase D2-dependent Inhibition of the Nuclear Hormone Receptor PPARγ by Cyclic Phosphatidic Acid. Mol. Cell, 39, 421–432.
16) Kobayashi, S., Tokunoha, R., Shibasaki, M., Shinagawa, R., & Murakami-Murofushi, K. (1993) Synthesis of 1-O-acylglycerol 2,3-cyclic phosphate:Determination of the absolute structure of PHYLPA, a specific inhibitor of DNA polymerase α. Tetrahedron Lett., 34, 4047–4050.
17) Mukai, M., Imamura, F., Ayaki, M., Shinkai, K., Iwasaki, T., Murakami-Murofushi, K., Murofushi, H., Kobayashi, S., Yamamoto, T., Nakamura, H., et al. (1999) Inhibition of tumor invasion and metastasis by a novel lysophosphatidic acid (cyclic LPA). Int. J. Cancer, 81, 918–922.
18) Uchiyama, A., Mukai, M., Fujiwara, Y., Kobayashi, S., Kawai, N., Murofushi, H., Inoue, M., Enoki, S., Tanaka, Y., Niki, T., et al. (2007) Inhibition of transcellular tumor cell migration and metastasis by novel carba-derivatives of cyclic phosphatidic acid. Biochim. Biophys. Acta, 1771, 103–112.
19) Baker, D.L., Fujiwara, Y., Pigg, K.R., Tsukahara, R., Kobayashi, S., Murofushi, H., Uchiyama, A., Murakami-Murofushi, K., Kohi, E., Bandle, R.W., et al. (2006) Carba analogs of cyclic phosphatidic acid are selective inhibitors of autotaxin and cancer cell invasion and metastasis. J. Biol. Chem., 281, 22786–22793.
20) Nozaki, E., Gotoh, M., Tanaka, R., Kato, M., Suzuki, T., Nakazaki, A., Hotta, H., Kobayashi, S., & Murakami-Murofushi, K. (2012) Pharmacological evaluation of a novel cyclic phosphatidic acid derivative 3-S-cyclic phosphatidic acid (3-S-cPA). Bioorg. Med. Chem., 20, 3196–3201.
21) Imamura, F., Horai, T., Mukai, M., Shinkai, K., Sawaa, M., & Akedo, H. (1993) Induction of in vitro tumor cell invasion of cellular monolayers by lysophosphatidic acid or phospholipase D. Biochem. Biophys. Res. Commun., 193, 497–503.
22) Ishihara, R., Tatsuta, M., Iishi, H., Baba, M., Uedo, N., Higashino, K., Mukai, M., Ishiguro, S., Kobayashi, S., & Murakami-Murofushi, K. (2004) Attenuation by cyclic phosphatidic acid of peritoneal metastasis of azoxymethane-induced intestinal cancers in Wistar rats. Int. J. Cancer, 110, 188–193.
23) Mukai, M., Iwasaki, T., Tatsuta, M., Togawa, A., Nakamura, H., Murakami-Murofushi, K., Kobayashi, S., Imamura, F., & Inoue, M. (2003) Cyclic phosphatidic acid inhibits RhoA-mediated autophosphorylation of FAK at Tyr-397 and subsequent tumor-cell invation. Int. J. Oncol., 22, 1247–1256.
24) Gotoh, M., Fujiwara, Y., Yue, J., Liu, J., Lee, S., Fells, J., Uchiyama, A., Murakami-Murofushi, K., Kennel, S., Wall, J., et al. (2012) Controlling cancer through the autotaxin-lysophosphatidic acid receptor axis. Biochem. Soc. Trans., 40, 31–36.
25) Kakiuchi, Y., Nagai, J., Gotoh, M., Hotta, H., Murofushi, H., Ogawa, T., Ueda, H., & Murakami-Murofushi, K. (2011) Antinociceptive effect of cyclic phosphatidic acid and its derivative on animal models of acute and chronic pain. Mol. Pain, 7, 33.
26) Inoue, M., Rashid, M.H., Fujita, R., Contos, J.J., Chun, J., & Ueda, H. (2004) Initiation of neuropathic pain requires lysophosphatidic acid receptor signaling. Nat. Med., 10, 712–718.
27) Gotoh, M., Nagano, A., Tsukahara, R., Murofushi, H., Morohoshi, T., Otsuka, K., & Murakami-Murofushi, K. (2014) Cyclic Phosphatidic Acid Relieves Osteoarthritis Symptoms. Mol. Pain, 10, 52.
28) Maeda-Sano, K., Gotoh, M., Morohoshi, T., Someya, T., Murofushi, H., & Murakami-Murofushi, K. (2014) Cyclic phosphatidic acid and lysophosphatidic acid induce hyaluronic acid synthesis via CREB transcription factor regulation in human skin fibroblasts. Biochim. Biophys. Acta, 1841, 1256–1263.
29) Sano, K., Gotoh, M., Dodo, K., Tajima, N., Shimizu, Y., & Murakami-Murofushi, K. (2018) Age-related changes in cyclic phosphatidic acid-induced hyaluronic acid synthesis in human fibroblasts. Hum. Cell, 31, 72–77.
30) 室伏きみ子,田中信治(2009)脂質メディエーター・環状ホスファチジン酸(cPA)の基礎化粧品への応用,月刊バイオインダストリー,26, 5–17.
31) Fujiwara, Y., Sebök, A., Meakin, S., Kobayashi, T., Murakami-Murofushi, K., & Tigyi, G. (2003) Cyclic phosphatidic acid elicits neurotrophin-like actions in embryonic hippocampal neurons. J. Neurochem., 87, 1272–1283.
32) Gotoh, M., Sano-Maeda, K., Murofushi, H., & Murakami-Murofushi, K. (2012) Protection of neuroblastoma Neuro2A cells from hypoxia-induced apoptosis by cyclic phosphatidic acid (cPA). PLoS One, 7, e51093.
33) Nakajima, S., Gotoh, M., Fukasawa, K., Murofushi, H., & Murakami-Murofushi, K. (2018) 2-O-carba-oleoyl cyclic phosphatidic acid induces glial proliferation through the activation of lysophosphatidic acid receptor. Brain Res., 1681, 44–51.
34) Gotoh, M., Hotta, H., & Murakami-Murofushi, K. (2010) Effects of cyclic phosphatidic acid on delayed neuronal death following transient ischemia in rat hippocampal CA1. Eur. J. Pharmacol., 649, 206–209.
35) Crack, P.J., Zhang, M., Morganti-Kossmann, M.C., Morris, A.J., Wojciak, J.M., Fleming, J.K., Karve, I., Wright, D., Sashindranath, M., Goldshmit, Y., et al. (2014) Anti-lysophosphatidic acid antibodies improve traumatic brain injury outcomes. J. Neuroinflammation, 11, 37.
36) Goldshmit, Y., Matteo, R., Sztal, T., Ellett, F., Frisca, F., Moreno, K., Crombie, D., Lieschke, G.J., Currie, P.D., Sabbadini, R.A., et al. (2012) Blockage of lysophosphatidic acid signaling improves spinal cord injury outcomes. Am. J. Pathol., 181, 978–992.
37) Hashimoto, K., Nakashima, M., Hamano, A., Gotoh, M., Ikeshima-Kataoka, H., Murakami-Murofushi, K., & Miyamoto, Y. (2018) 2-carba cyclic phosphatidic acid suppresses inflammation via regulation of microglial activation in the stab-wounded mouse cerebral cortex. Sci. Rep., 8, 9715.
38) Yamamoto, S., Gotoh, M., Kawamura, Y., Yamashina, K., Yagishita, S., Awaji, T., Tanaka, M., Maruyama, K., Murakami-Murofushi, K., & Yoshikawa, K. (2014) Cyclic phosphatidic acid treatment suppress cuprizone-induced demyelination and motor dysfunction in mice. Eur. J. Pharmacol., 741, 17–24.
39) Yamamoto, S., Yamashina, K., Ishikawa, M., Gotoh, M., Yagishita, S., Iwasa, K., Maruyama, K., Murakami-Murofushi, K., & Yoshikawa, K. (2017) Protective and therapeutic role of 2-carba-cyclic phosphatidic acid in demyelinating disease. J Neuroinflammation, 14, 142.
40) Tsukahara, T., Hanazawa, S., & Murakami-Murofushi, K. (2011) Cyclic phosphatidic acid influences the expression and regulation of cyclic nucleotide phosphodiesterase 3B and lipolysis in 3T3-L1 cells. Biochem. Biophys. Res. Commun., 404, 109–114.
41) Tsukahara, T., Hanazawa, S., Kobayashi, T., Iwamoto, Y., & Murakami-Murofushi, K. (2010) Cyclic phosphatidic acid decreases proliferation and survival of colon cancer cells by inhibiting peroxisome proliferator-activated receptor γ. Prostaglandins Other Lipid Mediat., 93, 126–133.
42) Williams, J.R., Khandoga, A.L., Goyal, P., Fells, J.I., Perygin, D.H., Siess, W., Parrill, A.L., Tigyi, G., & Fujiwara, Y. (2009) Unique ligand selectivity of the GPR92/LPA5 lysophosphatidate receptor indicates role in human platelet activation. J. Biol. Chem., 284, 17304–17319.
43) Konakazawa, M., Gotoh, M., Murakami-Murofushi, K., Hamano, A., & Miyamoto, Y. (2015) Effect of cyclic phosphatidic acid on the proliferation and differentiation of mouse cerebellar granule precursor cells during cerebellar development. Brain Res., 1614, 28–37.
44) Tigyi, G. (2010) Aiming drug discovery at lysophosphatidic acid targets. Br. J. Pharmacol., 161, 241–270.
45) Fujiwara, Y. (2008) Cyclic phosphatidic acid-a unique bioactive phospholipid. Biochim. Biophys. Acta, 1781, 519–524.
46) Tania, M., Khan, M.A., Zhang, H., Li, J., & Song, Y. (2010) Autotaxin:a protein with two faces. Biochem. Biophys. Res. Commun., 401, 493–497.
47) Nozaki, E., Gotoh, M., Hotta, H., Hanazawa, S., Kobayashi, S., & Murakami-Murofushi, K. (2011) Synthesis of enantiopure 2-carba-cyclic phosphatidic acid and effects of its chirality on biological functions. Biochim. Biophys. Acta, 1811, 271–277.
48) Gupte, R., Siddam, A., Lu, Y., Li, W., Fujiwara, Y., Panupinthu, N., Pham, T.C., Baker, D.L., Parrill, A.L., Gotoh, M., et al. (2010) Synthesis and pharmacological evaluation of the stereoisomers of 3-carba cyclic-phosphatidic acid. Bioorg. Med. Chem. Lett., 20, 7525–7528.
49) Tsukahara, T. & Murakami-Murofushi, K. (2012) Release of cyclic phosphatidic acid from gelatin-based hydrogels inhibit colon cancer cell growth and migration. Sci. Rep., 2, 687.

お茶の水女子大学 ヒューマンライフイノベーション研究所特任准教授.理学博士,Doctorat en chimie.
2003年お茶の水女子大学大学院博士課程入学.03~06年日仏共同博士課程派遣学生としてルイ・パスツール大学(現ストラスブール大学,Guy OURISSON・中谷陽一研究室)に留学.06年AIST特別研究員,07年よりお茶の水女子大学室伏きみ子教授に師事.テネシー大学(Gabor TIGYI研究室)を経て16年より現職.
研究テーマと抱負生理活性脂質の機能を生理学的な側面と物理化学的な側面から明らかにしていくことができれば嬉しいです.医学・薬学に貢献できるような研究を行っていきたいと思っています.
ウェブサイトhttp://www-w.cf.ocha.ac.jp/ihli/
趣味散歩,映画鑑賞.

お茶の水女子大学長.医学博士.
1970年お茶の水女子大学理学部卒業,72年同大学院理学研究科修士課程修了.76年東京大学大学院医学系研究科博士課程修了,95年お茶の水女子大学教授,ルイ・パスツール大学客員教授,2015年より現職.
研究テーマと抱負生理活性脂質・環状ホスファチジン酸とステリルグルコシドの生理作用について研究を続けて居り,研究成果を人々の健康な生活を維持するために役立てたいと考えています.
ウェブサイトhttp://www-w.cf.ocha.ac.jp/ihli/
趣味絵本作成.

This page was created on 2018-10-25T13:37:57.641+09:00
This page was last modified on 2018-12-13T17:38:47.843+09:00
このサイトは(株)国際文献社によって運用されています。