哺乳類では全タンパク質の約30%,言い換えれば約6000個の遺伝子を構成する膜タンパク質は,生体内の適切な膜へ仕分けられ局在する1).これらの膜タンパク質が膜間コミュニケーション,輸送,膜形態変化などの広範な細胞機能を脂質二重層にもたらす.それゆえに膜挿入機構は原核生物から存在し,進化過程を経て,より高度に適応発達した.
高疎水性の膜貫通ドメインを持つ膜タンパク質は,脂質二重層内に挿入されて働く.したがって膜タンパク質の一生における一大イベントは,水溶性の細胞質領域から最もエネルギー的に安定となる疎水性脂質二重層内への膜貫通ドメイン挿入ステップである2).この過程は複数段階からなる.つまり,選択的な膜貫通ドメインの認識,水溶性環境からの膜貫通ドメイン保護,標的膜上への輸送,正しい向きに応じた膜内挿入である.これら全段階を適切に進行するため,細胞質内または標的膜上に特異的かつ厳密な制御因子による介助が行われる.
真核細胞では,細胞質のリボソームで合成された膜タンパク質は小胞体,ミトコンドリア,ペルオキシソーム,葉緑体(植物)の膜へ運ばれる3–6).小胞体はこの中で最も多数の膜タンパク質に対応する.細胞膜,ゴルジ体,エンドソーム,核膜などの主に分泌経路をつかさどるすべての膜タンパク質が小胞体から運ばれるからである.小胞体膜への挿入は翻訳共役型膜挿入経路(co-translation)または翻訳後膜挿入経路(post-translation)のどちらかによりなされる.翻訳共役型膜挿入経路では,最初の膜タンパク質認識から最後の膜挿入まで全段階が当該膜タンパク質合成中に行われる.一方で,翻訳後膜挿入経路における認識と挿入段階は,膜タンパク質合成が完了した後に行われる.したがってリボソームは,翻訳共役型膜挿入経路の全段階で主要な機能因子として働くが,翻訳後膜挿入経路では初期段階のみに限定的な関わりを持つ.
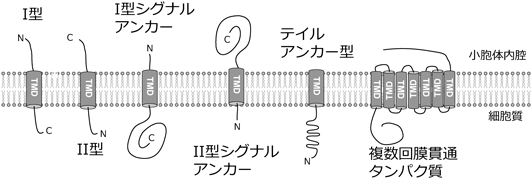
膜タンパク質は,挿入される方向やシグナルペプチドの有無,タンパク質内の膜貫通ドメインの位置によって複数のタイプに分けられる(図1).まず,(1)I型はN末端が内腔側になる.II型はC末端が内腔側になる.(2)シグナルアンカーはシグナルペプチドを持たない膜タンパク質の疎水性ドメインがシグナルと膜貫通ドメイン両方を兼ねるものを指す.(3)複数回膜貫通ドメインは二つ以上の膜貫通ドメインを持つ膜タンパク質である.(4)テイルアンカー型はC末端に一つの膜貫通ドメインを持つため,リボソームから放出後にC末端の膜貫通ドメインが認識・挿入され,N末端領域を細胞質側に露出する.前者の三つは翻訳共役型膜挿入経路によって膜挿入される.一方で,テイルアンカー型膜タンパク質は膜貫通ドメインがC末端から40アミノ酸以内にあるため,アミノ酸長40個分しかないリボソームトンネルから翻訳中の膜貫通ドメインは認識できない.そのためテイルアンカー型膜タンパク質は翻訳後膜挿入経路を利用して膜挿入される.本稿では扱わないが,GPIアンカー型膜タンパク質や,両親媒性ドメインを持つ膜タンパク質,βバレル型膜タンパク質など高疎水性膜貫通ドメインとは異なる挿入ドメインを持つ膜タンパク質も存在する.
翻訳共役型膜挿入経路は40年以上前に発見され,詳細な機構が解明された7).しかし小胞体膜タンパク質の翻訳後膜挿入経路はつい10年ほど前に必要因子が同定された.本機構は,細胞質のシャペロン(認識と保護)と特異的小胞体膜上受容体(標的化と挿入)の相互作用を必要とする点で他オルガネラへの水溶性タンパク質の翻訳後輸送と類似している.本稿では,翻訳後膜挿入経路を利用する膜タンパク質の輸送経路とタンパク質品質管理システムを中心に概説する.
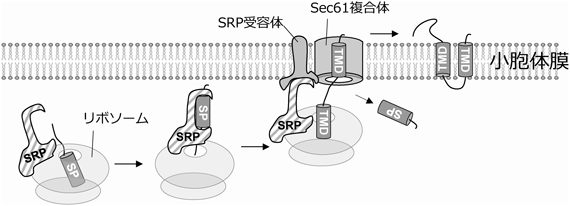
これまで盛んに研究されてきた翻訳共役型膜挿入経路は,翻訳後膜挿入経路を理解する上で基礎となる重要な概念をもたらした.そのため翻訳後膜挿入経路を説明する前に,翻訳共役型膜挿入経路を介した膜タンパク質の挿入機構をまず解説する8).この経路はリボソーム出口トンネルから翻訳中タンパク質の膜貫通ドメインが細胞質に露出して開始する(図2).数種のタンパク質とRNAから構成されるリボヌクレオタンパク質SRP(signal recognition particle)がその膜貫通ドメインを認識する9).SRPのサブユニットSRP54はリボソームに対して高親和性であり,リボソーム出口トンネル近傍に結合している.SRP54が持つMドメイン(メチオニンリッチドメイン)はシグナルペプチドなどの高疎水性ドメインに結合するため,リボソーム出口トンネル表層で膜貫通ドメインを持つ新生鎖の出現を待ち構えている.SRPが膜貫通ドメインに結合すると,SRP結合型リボソーム新生鎖複合体は小胞体膜上のSRP受容体へ輸送される.リボソーム新生鎖複合体はSRPから解離し,小胞体トランスロコンの中心因子であるSEC61複合体上に移り翻訳を続ける.その後,リボソームはSEC61トランスロコンに結合したまま膜タンパク質合成を完了する.SEC61トランスロコンはリボソームから出現したそれぞれの膜貫通ドメインを認識するよう配置されており,膜タンパク質の脂質二重層へのトランスロケーションを促進する.
翻訳共役型の大きな利点は,標的化と挿入機構がリボソームのポリペプチド出口トンネルと物理的に共役している点である.水溶性細胞質領域への露出を最低限に抑えつつ,疎水性膜貫通ドメインの認識・保護・挿入を可能にすることができるのは,このような空間的調節の賜物である.つまり,翻訳共役型膜挿入経路は膜貫通ドメインを認識・結合する点で,非特異的に結合しうる他の細胞質因子よりも相当な利点を得ている.とりわけ複数回膜貫通型膜タンパク質においては,リボソームから出現した高疎水性かつ冗長な複雑きわまりない膜タンパク質の可溶性を維持する必要性を実質的に取り除くことができる.このため,複数回膜貫通型膜タンパク質がトランスロケーションするために翻訳共役型を利用することはほぼ普遍的である.実際に,トポロジーが相同な細菌の細胞膜も共翻訳型で膜タンパク質をトランスロケーションする10).また最近では,一部のミトコンドリア膜タンパク質でも観察された11).
SRP依存的経路の解明は,他経路の発見を導いた.つまり少なくともいくつかのタンパク質群はSRP非依存的に膜挿入をなしていることが示唆された.タンパク質合成中にSRPによって認識されないタンパク質を論理的に考えてみると,浮かび上がってくるものは,C末端に一つの膜貫通ドメインを保持するテイルアンカー型膜タンパク質である12).上述したとおり,リボソーム出口トンネル長はアミノ酸40個分しかない.一方でテイルアンカー型膜タンパク質の膜貫通ドメインからC末端までは40アミノ酸残基に満たない.すなわち,テイルアンカー型膜タンパク質の膜貫通ドメインがリボソームから露出するときには翻訳終結してリボソームから放出されるため,SRP認識から逃れてしまう.
SRP非依存性が証明された最初のテイルアンカー型膜タンパク質は,細胞内小胞融合の主要因子SNAREタンパク質synaptobrevin(別名VAMP)である.予想どおりsynaptobrevinがSRPとSEC61非依存的翻訳後経路を利用して小胞体膜に挿入されることがはっきりと立証されたのは1995年のことであった13).後続の研究から,この経路には未知のタンパク質因子とATPが要求されることも示された.しかし,この挿入に関わる遺伝子とエネルギー要求性の分子機構は10年以上にわたり日の目をみることがなかった6, 14).
挿入経路の解明が未解決のままであった一方で,テイルアンカー型膜タンパク質が持つ広範な生理的重要性が認知された.全膜タンパク質の約3~5%を占めるテイルアンカー型膜タンパク質は細胞内のあらゆる膜に存在しており,膜生合成や,アポトーシス,小胞輸送,タンパク質分解など数多の機能を担っていることがわかった.テイルアンカー型膜タンパク質の注目が高まるに従って,その未知の挿入経路についても多くの研究者による解析が進められた.しかしながら,それぞれの研究者による異なる手法を用いた別々のテイルアンカー型膜タンパク質の研究は多様な相容れない結果を導いてしまった.たとえば,テイルアンカー型膜タンパク質の一つcytochrome b5のin vitro実験からはATPもタンパク質因子も必要ない単独挿入経路が示唆される一方で15),HSP70(heat shock protein 70)が仲介する挿入経路や16),SRPもしくはSEC61のどちらか片方が翻訳後経路にも働いていることなどが示唆された17).これらの経路の生理的妥当性は現在でも不明なまま取り残されている.
最終的に,タンパク質架橋法と無細胞タンパク質翻訳システムを組み合わせたテイルアンカー型膜タンパク質挿入の生化学的実験からテイルアンカー型膜タンパク質の膜貫通ドメインと相互作用する細胞質因子が同定され,TRC40(TMD-recognition complex protein of 40 kDa,別名Asna1)と名づけられた18, 19).TRC40が候補因子となった理由を以下に示す.第一に,TRC40はテイルアンカー膜タンパク質のC末端の膜貫通ドメインに結合するが,タンパク質内側の膜貫通ドメインには結合しない.第二に,TRC40はATPase活性を持つ(翻訳後膜挿入経路はATPを要求する).第三に,すべての真核生物で高い保存性を持ち,哺乳類では必須遺伝子である.さらに酵母における変異体はテイルアンカー型膜タンパク質が関わる機能の欠損がみられた.第四に,ドミナントネガティブとなるATPase欠損変異体は,テイルアンカー型膜タンパク質に結合するが解離できないことで,膜への挿入が抑制される.第五に,TRC40の細胞画分の一部は小胞体にみられ,ATPによって放出される.最後に,小胞体ミクロソーム上のタンパク質性因子と結合する.これらの観察に基づき,TRC40は細胞質テイルアンカー型膜タンパク質の選択的な標的化因子であり,ATP依存的な挿入のため小胞体へ搬送されることが考えられた.
TRC40との高い相同性から出芽酵母ホモログ因子Get3はすぐに同定された.当時すでにGet3は,遺伝学と物理的相互作用の研究から少なくとも二つの異なるタンパク質(Get1とGet2)とともにGet(Golgi–ER trafficking pathway)と呼ばれる経路を構成する因子群であることが知られており,実際にこれらのうちどれかを欠損するとゴルジ体–小胞体間輸送の異常を引き起こす表現型が報告されていた20).しかし,ゴルジ体–小胞体間輸送はテイルアンカー型膜タンパク質に依存する(前述したようにSNAREタンパク質はテイルアンカー型膜タンパク質に属する)という事実を考慮すると,Get経路について再考する必要が生じた20).実際に,Get経路が示す表現型はテイルアンカー型膜タンパク質挿入の欠損とよく相関した.したがって,Get遺伝子は“guided entry of TA proteins”と現在は呼ばれている.後続した酵母Get経路の物理的・遺伝学的相互作用解析によってさらなる三つの因子Get4, Get5, Sgt2が同定された.これにより小胞体翻訳後膜挿入経路の主要キャストが決定し,その後,構造解析や生化学実験を組み合わせる研究から膜挿入メカニズムの詳細がもたらされた.
4. 小胞体テイルアンカー型膜タンパク質のターゲッティング
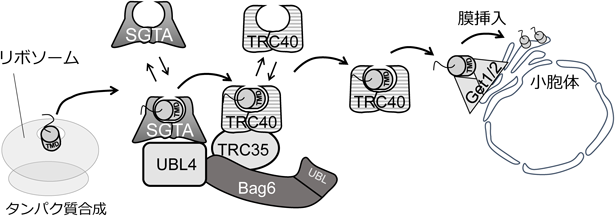
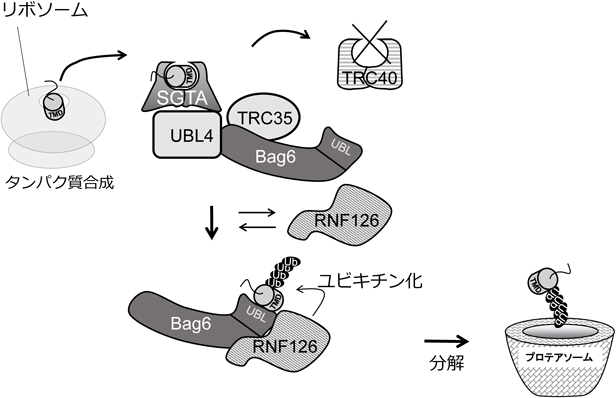
遺伝学的解析から同定された主要因子を用いて,酵母および哺乳類細胞による生化学的解析が進められ,小胞体テイルアンカー型膜タンパク質の翻訳後膜挿入経路の分子機構が解明された.本節では哺乳類での知見を中心に紹介する(図3).TRC40は膜貫通ドメインを小胞体膜上のGet1/Get2複合体へ搬送する重要な因子であるが,そもそもリボソームから放出されたテイルアンカー型膜タンパク質にTRC40が直接結合できるわけではない.なぜならSRPとは異なり,TRC40はリボソーム上に存在しないからである.そこで活躍するのがSGTA(酵母Sgt2のホモログ),TRC35(酵母Get4のホモログ),UBL4A(酵母Get5のホモログ)である.TRC35とUBL4Aは複合体を形成しており,さらにSGTAと結合能を有する.テイルアンカー型膜タンパク質の膜貫通ドメインはSGTAに結合するとTRC35/UBL4A複合体に結合する.重要なことに膜タンパク質結合能を持つ閉鎖型TRC40はTRC35/UBL4A複合体と選択的に結合できる.つまり膜タンパク質を連れてきたSGTAが閉鎖型TRC40へ膜タンパク質を受け渡す足場としてTRC35/UBL4A複合体が働く.哺乳類ではさらにBag6がTRC35/UBL4A複合体に含まれ,SGTAと同様に膜貫通ドメインに直接結合することができる21).またBag6を欠損するとTRC40の膜タンパク質への結合が減少する.つまりSGTA/TRC35/UBL4A/Bag6がプレターゲッティング因子複合体として働く.Bag6はリボソームと結合することもできると考えられている.したがって,Bag6が他のプレターゲッティング因子とともにリボソーム上にリクルートされることで,放出された膜タンパク質を速やかにSGTAもしくはBag6が捉えるようになっていると考えられている.この捕捉段階の詳しい分子機構はさらなる研究から明らかにする必要がある.
膜タンパク質と結合したTRC40は小胞体へ向かう.小胞体膜上受容体となるのがGet1/Get2複合体である.テイルアンカー型膜タンパク質の挿入段階はTRC40とGet1/Get2複合体の三者のみで実行できることがin vitro実験から実証されている.まず膜タンパク質と複合体形成した閉鎖型TRC40はGet2の細胞質側の領域と結合する.続いてGet2からGet1へTRC40が受け渡される.このときGet1のコイルドコイルドメインがTRC40の疎水性の溝に入り込み,ATPase活性依存的に開放型TRC40へ変換させることで基質が放出され膜挿入されることがわかっている.
5. 細胞質のテイルアンカー型膜タンパク質結合因子:SGTAとカルモジュリン
SGTAとカルモジュリンは膜タンパク質のターゲッティングやテザリングに直接関わらない膜貫通ドメイン結合タンパク質である.これらのタンパク質は主に膜タンパク質が細胞質で凝集体や不適当な結合を防ぐために働く.基質である膜タンパク質が標的膜または標的因子まで搬送されるまで,SGTAもしくはカルモジュリンが動的に結合する.
SGTAは小胞体テイルアンカー型膜タンパク質の最上流因子だと考えられていた22).しかし結合は小胞体テイルアンカー型膜タンパク質に特異的というわけではなく,またin vitro実験や酵母での欠損から,他のGet遺伝子変異体と比べて軽度な表現型を示す.そのため,SGTAはダイナミックかつ広範囲な特異性を持つ膜貫通ドメイン結合因子と注目されている.もしSGTAが結合した膜貫通ドメインが十分に高疎水性であれば,小胞体へ運ぶためTRC40へ速やかに移される.もし疎水性が十分でなければ,膜タンパク質はSGTAから離れ,ミトコンドリアターゲッティング経路や,分解機構へ送られる.このようにSGTAは基質の凝集を防ぐための一般的な膜貫通ドメイン結合タンパク質であり,さらにTRC40への適合性を判定する.SGTAへの基質結合のメカニズムは不明なままだが,TRC40とSRPの膜貫通ドメイン結合ドメインと似たメチオニンリッチな結合ドメインを持つと考えられている.
カルモジュリンもメチオニンリッチなαヘリックスを含む構造を介して基質に結合し,いろいろなペプチド配列に対して非常に広い結合性を持つ23).細胞質内の生理的なカルシウム濃度下において,カルモジュリンはシグナルペプチドや膜貫通ドメインなど,特に中程度の疎水性の配列へ動的に接着する.さまざまな基質とカルモジュリンの構造解析から,カルモジュリンの二つの突出部が疎水性ドメインに巻きつくことで水溶性領域を保護することが示唆されている.カルモジュリンによって保護できる疎水性領域長(約18アミノ酸まで)はHsp70のような普遍的なシャペロンタンパク質よりも長い.可溶性タンパク質の折りたたみでは,シャペロンが3~5アミノ酸程度の疎水性領域を扱うが,シグナルペプチドや膜貫通ドメインの疎水性領域はより長いため,結合能に違いがあると考えられる.細胞質のカルモジュリンはとても豊富に存在し,広範囲の生理的カルシウム濃度下で動的結合できることから,シグナルペプチドや膜貫通ドメインの凝集体化を和らげる作用を持つ.しかし,カルモジュリンはSGTAと異なり,標的化因子に直接結合をしない.そのため,カルモジュリンは膜タンパク質の輸送ではなく,輸送されていない余剰な細胞質の膜タンパク質を一時的に保護するために働くのだろう.
さて,前述したように動物細胞は小胞体,ミトコンドリア,ペルオキシソームに膜タンパク質ターゲッティングシステムを備えている.言い換えれば,ゴルジ体や細胞膜など他のオルガネラへ膜タンパク質が直接“突き刺さる”ということはない.だが他のオルガネラにも実際数多くの膜タンパク質が存在する.ゴルジ体や細胞膜は小胞体から小胞を介した膜間輸送によって膜タンパク質が運ばれ,必要な膜タンパク質を獲得できる.では他オルガネラと膜間輸送が不明なオートファゴソーム膜はどうやって膜タンパク質を獲得するのだろうか?
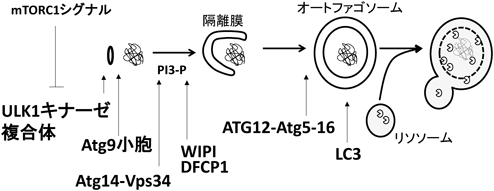
筆者は博士課程から哺乳類オートファゴソーム形成の分子機構に関する研究に従事した.オートファジーとは,細胞質内に出現した隔離膜が弧を描くように膜伸張しながら細胞内成分を囲い込んでオートファゴソームを形成し,オートファゴソーム外膜とリソソーム膜が融合することで内容物をリソソーム酵素に曝露し分解する大規模タンパク質分解システムである24).一過的な膜構造体であるオートファゴソームのダイナミックな形成機構は,10数個からなるATGタンパク質群が担う25).ATG遺伝子は酵母を用いたスクリーニングによって同定され,その哺乳類ホモログも同定されたが,各ATGタンパク質がオートファゴソーム形成時にどのような働きをするのかわかっていなかった.筆者らはATGどうしをタンパク質複合体ごとにグループ化し,それぞれのATGタンパク質複合体がオートファゴソーム形成に働く順番を調べた(図4)26).その結果,まずオートファジー誘導に伴いULK1キナーゼ複合体が小胞体上のオートファゴソーム形成部位にリクルートされる.同じタイミングでAtg9小胞も同部位に局在する.両者がそろうと,続いてリクルートされるAtg14-Vps34複合体がホスファチジルイノシトール3-リン酸を産生することで隔離膜形成が開始される.隔離膜近傍に産生されたホスファチジルイノシトール3-リン酸にWIPIとDFCP1が結合する.その後,Atg7とAtg10によるAtg12ユビキチン様コンジュゲーションシステムによって,Atg12と共有結合したAtg5複合体も隔離膜にリクルートされる.さらにLC3コンジュゲーションシステムによって,ホスファチジルエタノールアミン(PE)と共有結合したLC3-PEも隔離膜にリクルートされ,オートファゴソームが完成する.オートファゴソームが完成するとLC3を除く他のAtgタンパク質は解離する.完成したオートファゴソームの外膜がリソソーム膜と膜融合を引き起こし,オートファゴソームの内容物がリソソーム酵素に曝露され分解される.このAtgタンパク質の階層性はオルガネラを分解する選択的オートファジーにも対応する27, 28).
Atgタンパク質群はダイナミックな膜形成に必須な役割を持つが,膜貫通ドメインを持つAtgタンパク質はAtg9のみである.しかしそのAtg9は隔離膜の形成初期に関わるが,オートファゴソーム膜に含まれていない.そのためオートファゴソーム膜には膜貫通ドメインを有する膜タンパク質が存在するのか知られていなかった.言い換えると,オートファゴソーム膜に膜タンパク質が局在する経路があるのかどうかもわからなかった.だが膜タンパク質がないと仮定すると説明できないことが生じる.リソソーム膜との融合過程である.細胞内膜系の膜融合は多くの場合,双方の膜上に存在するテイルアンカー型膜タンパク質であるSNAREタンパク質が仲介する.実際にリソソームはSNAREタンパク質を介してエンドソーム小胞と融合する.オートファゴソームとリソソームの膜融合がSNAREタンパク質を介するのであれば,オートファゴソーム膜にもSNAREタンパク質が存在するはずである.
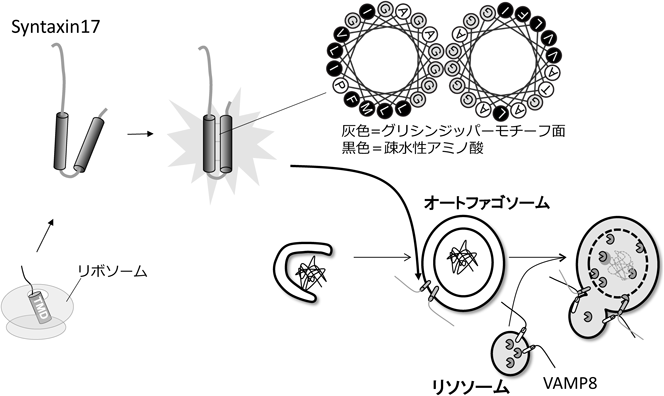
筆者らはオートファゴソーム形成部位に関わる因子を探索している過程でSNAREタンパク質Syntaxin17(Stx17)がオートファゴソームに局在することを発見した.オートファゴソーム膜の一部は小胞体膜から供給されていると考えられるため,小胞体膜とStx17がともに隔離膜形成時にオートファゴソーム膜に供給されると予想した.しかし隔離膜上にStx17はまったく観察されないにもかかわらず,完成したオートファゴソームにStx17が局在した.筆者らはこの結果に大変困惑した.いかにして膜構造と連結されていないオートファゴソーム膜に膜タンパク質が新しく供給されるのか? ところが,この現象を丁寧にひもとくことで,とてもユニークなオートファゴソームとテイルアンカー型膜タンパク質の関係が鮮明になった.
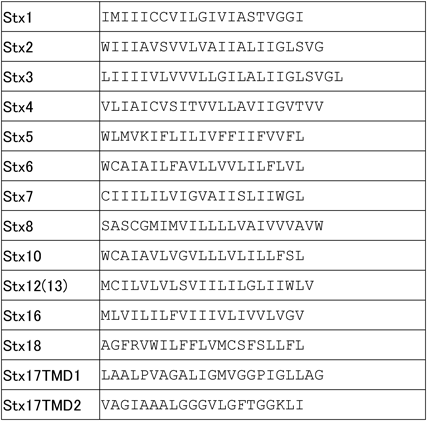
オートファゴソーム局在に必要なドメインをStx17タンパク質から抽出してみると,膜貫通ドメイン領域がオートファゴソームへの局在に必須であることが示唆された.そこでStx17膜貫通ドメインに秘密があるだろうと考え,他のSNAREタンパク質の膜貫通ドメインのアミノ酸配列と比較を行った(図5).すると,興味深い二つの特徴が浮かび上がった.一つ目は,Stx17の膜貫通ドメインがグリシンリッチであること,二つ目は,Stx17が二つの膜貫通ドメインを持つことである.両者ともテイルアンカー型膜タンパク質として非普遍的な特徴である.さらにアミノ酸三つごとに三つのグリシンが並ぶグリシンジッパー配列も発見した.これらの事実から,二つのStx17膜貫通ドメインはグリシンジッパーが形成する溝に向かい合うように密着したヘアピン型膜貫通ドメイン形態をとることが予想される.興味深いことに,内側のグリシンジッパーどうしが結合すると,外側には高疎水性アミノ酸が露出する(図6).つまり二つのドメインが一つの膜貫通ドメインの体をなす.グリシンジッパーによるヘアピン構造が重要であるか調べるために,グリシンをロイシンに置換した変異体を作製した.その結果,変異体Stx17はオートファゴソームに局在せず,リソソームとの融合能を欠損した.つまりStx17は特殊なヘアピン型膜貫通ドメインを利用してオートファゴソーム膜に局在することを発見した(図6)29).さらにStx17はエンドソームの係留に働くHOPS複合体にも結合する30).つまり,オートファゴソームとリソソームはHOPS複合体によって近接し,Stx17はリソソームSNAREであるVAMP8,さらにSNAP29とSNARE複合体を形成することでオートファゴソームとリソソーム膜の融合を促進する.
膜タンパク質のターゲッティングの視点から考察すると大きな謎が残る.Stx17はどこからオートファゴソーム膜に挿入されるのだろうか? 前述したように,Stx17は隔離膜に局在しない.そして最も注目すべきことは,Stx17の膜貫通ドメインの疎水性が比較的低いことである.実際に我々の実験からStx17の一部は細胞質に存在する.細胞質Stx17がオートファゴソーム膜に直接挿入される可能性がある.一方で,同時期からStx17に関する機能解析が進み,オートファゴソーム形成初期や,ミトコンドリアにおける機能も他のグループから報告された31, 32).挿入機構も含めStx17の詳細な分子機構は今後解明すべき課題である.
8. 小胞体テイルアンカー型膜タンパク質の品質管理
合成された膜タンパク質がすべて膜に挿入できるわけではなく,挿入できない膜タンパク質が少なからず確率的に生じる.また小胞体ストレスや,膜タンパク質の過剰生産も小胞体膜タンパク質の効率に影響する.疎水性ドメインの露出が引き起こす凝集体形成を防ぐため,細胞質膜タンパク質品質管理経路が存在する.Bag6は小胞体テイルアンカー型膜タンパク質の標的化に関わるだけでなく,N末端のUBL(ubiquitin-like)ドメインにユビキチンリガーゼRNF126をリクルートすることで,基質のユビキチン化とプロテアソーム分解も仲介することが発見された22, 33, 34).膜貫通ドメインである20アミノ酸長の高疎水性ドメインにBag6は高い親和性を持つ.この結合の解離速度はとても遅いので,生理的時間上では不可逆的な結合である.他の膜貫通ドメイン結合因子と異なり,Bag6の膜貫通ドメイン結合部位の構造はまだわかっておらず,またメチオニンリッチドメインを含む領域が結合に関与している可能性もない.
Bag6のC末端領域はTRC40基質複合体が小胞体へターゲッティングするために必須な構造を作る.実際に,このC末端領域はUBL4AとTRC35に結合することで,それぞれSGTAとTRC40を同複合体にリクルートする.膜貫通ドメインと直接結合できなくなったUBLドメイン欠損変異体Bag6でもTRC40の結合を介したテイルアンカー型膜タンパク質のターゲッティングが正常に機能する.一方で,酵母にはBag6のホモログが存在しないため,UBL4A(Get5)とTRC35(Get4)は直接Get3基質複合体に結合することがわかっている35).つまり,Bag6はテイルアンカー型膜タンパク質の小胞体ターゲッティング経路上でターゲッティングできない基質を分解へ導く.テイルアンカー型膜タンパク質以外の膜タンパク質や他の疎水性ドメインなど広範囲な高疎水性ドメインに結合できるBag6は,多くの誤局在した膜タンパク質の品質管理を担っている.
Bag6は結合したテイルアンカー型膜タンパク質の膜ターゲッティングと分解をどのように選別するのだろうか? 光架橋技術を利用したin vitro実験からBag6複合体のタンパク質結合速度を求め詳細がわかった.フリーのTRC40が存在する場合,Bag6複合体上のSGTAに結合したテイルアンカー型膜タンパク質はTRC40へすぐさま受け渡され,小胞体へのターゲッティングが続いて起こる.しかしTRC40が不足するときや,TRC40と親和性が高くない膜タンパク質の場合は,SGTAに結合した膜タンパク質はBag6-SGTA複合体に,より長い時間結合することになる.膜タンパク質は,Bag6-SGTA複合体上に20秒以上存在するとBag6のUBLドメインに移され,RNF126によるユビキチン化を受ける.このような時間依存的なタンパク質品質管理モデルが提唱されている(図7)22).
9. UBQLNによるミトコンドリア膜タンパク質の品質管理経路
ミトコンドリアのほとんどのタンパク質は核ゲノムにコードされており,哺乳類では約1000~1500個のミトコンドリア遺伝子が細胞質で合成される36).すべてのミトコンドリア膜タンパク質は疎水性のミトコンドリアターゲッティングシグナルもしくは疎水性の膜貫通ドメインを有する.Hsp70などのシャペロンが細胞質のミトコンドリアタンパク質に結合することは知られていたが,Hsp70が疎水性ドメインを直接保護することは難しい.そのため,ミトコンドリアへ輸送されるまでこれらの疎水性ドメインは他のタンパク質によって保護されるはずである.
筆者らはミトコンドリアテイルアンカー型膜タンパク質Omp25をモデル基質として細胞質ミトコンドリア膜タンパク質の品質管理経路の分子機構解明を試みた.ミトコンドリア膜へ挿入される細胞質膜タンパク質の挙動を捉えることは,培養細胞を用いたin vivo実験では非常に難儀である.なぜなら培養細胞内に発現させた膜タンパク質は膜へすぐさま挿入されてしまうからである.そのためウサギ網状赤血球ライセート(RRL)を利用したin vitroタンパク質合成を用いてOmp25を合成した.重要なポイントはRRLにはタンパク質翻訳因子だけでなく,一般的な細胞質因子も含まれているが,オルガネラなどの膜成分は含まれていないことである.そのため合成したOmp25はミトコンドリアへ挿入されないが,もし細胞質因子中に膜貫通ドメインを保護するシャペロンが存在するならば,Omp25と結合することが期待される.実際にRRLで合成したOmp25に結合するタンパク質を質量分析解析すると,UBQLN1, 2, 4が同定された37).またUBQLNはミトコンドリアテイルアンカー型膜タンパク質だけでなく,ミトコンドリアの2回膜貫通型タンパク質ATP5G1にも結合することがわかった.UBQLNはUBLドメインとUBA(ubiquitin associating)ドメインを持つUBL-UBAタンパク質の一つである.UBLドメインはプロテアソームと結合し,UBAドメインはユビキチンと結合するため,UBL-UBAタンパク質はユビキチン化タンパク質をプロテアソームへ輸送する単純なシャトリング因子であると一般的に考えられている.
UBQLNとOmp25の結合部位を調べてみると,UBQLNのSti1ドメインがOmp25との結合に重要であり,Sti1ドメインを欠損したUBQLNはOmp25の凝集体形成を抑制できなくなった事実と機能的に一致した.興味深いことに,UBQLNのSti1ドメインはメチオニンが豊富に存在するため,他の膜貫通ドメイン結合タンパク質と同じようにメチオニンリッチのポケットを形成して結合するのかもしれない.また凝集体蓄積を示す神経変性疾患の一つである筋萎縮性側索硬化症(ALS)の原因遺伝子としてUBQLN2の変異が報告されていることからも,UBQLNが品質管理システムとして機能することが予想された38).
UBQLN1と2と4のタンパク質すべてを欠損したUBQLN KO細胞を作製し解析すると,過剰発現したミトコンドリア膜タンパク質はミトコンドリアに局在できるが,一部が界面活性剤不溶性の凝集体を形成することがわかった.この凝集体はミトコンドリアシグナルペプチドが切断されていなかったことから,ミトコンドリアに挿入される前に凝集体化したと考えられた.さらにミトコンドリア脱分極剤処理によってミトコンドリアへの移行を阻害すると,野生型の細胞では細胞質のミトコンドリアタンパク質は分解されるが,UBQLN KO細胞では分解が減少した.これらのことからUBQLNがミトコンドリア膜タンパク質シャペロンと分解に関与していることがわかった.
次にUBQLN複合体にユビキチン化活性があるかどうか検討した.RRLから精製したOmp25-UBQLN複合体をユビキチン化アッセイすると,Omp25のユビキチン化や,ユビキチン化Omp25とUBQLNの結合強度増加がみられた.またユビキチン化Omp25と結合したUBQLNはプロテアソームとの結合能も増加した.
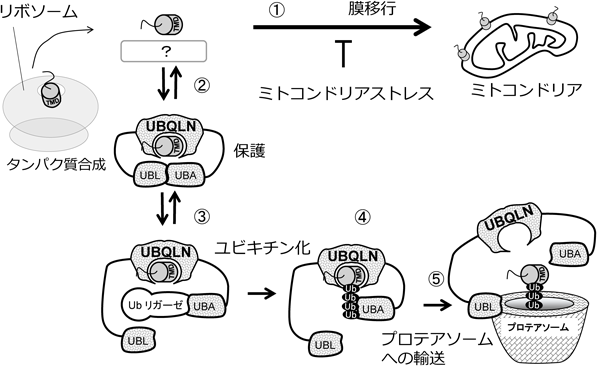
これらの結果から次のモデルが考えられる(図8)37).①リボソームから放出されたミトコンドリア膜タンパク質は,未知の機構によってミトコンドリアへターゲッティングされ膜挿入される.②ミトコンドリアストレスによってミトコンドリアへの移行が阻害されると,細胞質の余剰ミトコンドリア膜タンパク質の膜貫通ドメインにUBQLNのSti1ドメインが結合して保護する.これにより膜タンパク質の凝集体化を防ぐ.③ミトコンドリアストレスが回復しない場合は,時間依存的にUBQLN複合体のUBAドメインに未知のユビキチンリガーゼがリクルートされる.④ユビキチンリガーゼによってUBQLN複合体の膜タンパク質がユビキチン化される.さらに,UBQLNのUBAドメインがユビキチン鎖に結合することでUBQLNと膜タンパク質の結合が強固になり,ミトコンドリアへの輸送が完全に抑制される.⑤UBAドメインと自己会合していたUBLドメインがフリーとなるため,UBLドメインがプロテアソームをリクルートし,ユビキチン化膜タンパク質の分解を促す.つまりUBQLNは細胞質のミトコンドリア膜タンパク質品質管理システムとして働き,膜タンパク質の保護・ユビキチン化・プロテアソームへの輸送を一挙に担う.膜タンパク質を他タンパク質間で受け渡す必要がなく,膜貫通ドメインの露出を最低限に保つことができる.
UBQLNはミトコンドリア膜タンパク質に結合する一方で,Bag6は小胞体膜タンパク質に結合する.この識別機構はそれぞれが結合する膜タンパク質の疎水性度に応じた指向性の違いであり,UBQLNは低疎水性のドメインと,Bag6は高疎水性のドメインと結合する.実際にミトコンドリアの膜貫通ドメインは低疎水性であるが,小胞体の膜貫通ドメインは高疎水性である39).そのため,厳密にいえば,小胞体の膜タンパク質でも疎水性度が比較的低い膜貫通ドメインにはUBQLNは結合する.
膜タンパク質の疎水性ドメインは細胞質では非常に不安定だが,膜の疎水性領域に挿入されると安定化する.真核生物はさまざまなオルガネラを持つため,挿入する膜を厳密に調節しなければならない.しかし,in vitro実験をしてみると,小胞体のターゲッティング因子にミトコンドリアの膜タンパク質はある程度結合できることがわかる.またミトコンドリアへのターゲッティングに関わる因子はいまだ不明ではあるが,ミトコンドリア膜に他オルガネラの膜タンパク質が挿入されてしまうことがある.このような場合は,ミトコンドリア外膜タンパク質MSP1が誤局在したミトコンドリア上の膜タンパク質を引っこ抜き分解へ導く別の品質管理システムが存在する40).したがって,膜タンパク質の局在は本稿で紹介した以外にもさまざまな段階で調節されており,細胞はそれらが適切な目的地で機能するよう厳格な校正機能を持っているのだろう.
引用文献References
1) Fagerberg, L., Jonasson, K., von Heijne, G., Uhlén, M., & Berglund, L. (2010) Prediction of the human membrane proteome. Proteomics, 10, 1141–1149.
2) White, S.H. & von Heijne, G. (2008) How translocons select transmembrane helices. Annu. Rev. Biophys., 37, 23–42.
3) Chacinska, A., Koehler, C.M., Milenkovic, D., Lithgow, T., & Pfanner, N. (2009) Importing mitochondrial proteins:machineries and mechanisms. Cell, 138, 628–644.
4) Heiland, I. & Erdmann, R. (2005) Biogenesis of peroxisomes. Topogenesis of the peroxisomal membrane and matrix proteins. FEBS J., 272, 2362–2372.
5) Inaba, T. & Schnell, D.J. (2008) Protein trafficking to plastids:one theme, many variations. Biochem. J., 413, 15–28.
6) Shao, S. & Hegde, R.S. (2011) Membrane protein insertion at the endoplasmic reticulum. Annu. Rev. Cell Dev. Biol., 27, 25–56.
7) Blobel, G. & Dobberstein, B. (1975) Transfer of proteins across membranes. I. Presence of proteolytically processed and unprocessed nascent immunoglobulin light chains on membrane-bound ribosomes of murine myeloma. J. Cell Biol., 67, 835–851.
8) Keenan, R.J., Freymann, D.M., Stroud, R.M., & Walter, P. (2001) The signal recognition particle. Annu. Rev. Biochem., 70, 755–775.
9) Walter, P. & Blobel, G. (1980) Purification of a membrane-associated protein complex required for protein translocation across the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA, 77, 7112–7116.
10) Ulbrandt, N.D., Newitt, J.A., & Bernstein, H.D. (1997) The E. coli signal recognition particle is required for the insertion of a subset of inner membrane proteins. Cell, 88, 187–196.
11) Williams, C.C., Jan, C.H., & Weissman, J.S. (2014) Targeting and plasticity of mitochondrial proteins revealed by proximity-specific ribosome profiling. Science, 346, 748–751.
12) Kutay, U., Hartmann, E., & Rapoport, T.A. (1993) A class of membrane proteins with a C-terminal anchor. Trends Cell Biol., 3, 72–75.
13) Kutay, U., Ahnert-Hilger, G., Hartmann, E., Wiedenmann, B., & Rapoport, T.A. (1995) Transport route for synaptobrevin via a novel pathway of insertion into the endoplasmic reticulum membrane. EMBO J., 14, 217–223.
14) Hegde, R.S. & Keenan, R.J. (2011) Tail-anchored membrane protein insertion into the endoplasmic reticulum. Nat. Rev. Mol. Cell Biol., 12, 787–798.
15) Brambillasca, S., Yabal, M., Soffientini, P., Stefanovic, S., Makarow, M., Hegde, R.S., & Borgese, N. (2005) Transmembrane topogenesis of a tail-anchored protein is modulated by membrane lipid composition. EMBO J., 24, 2533–2542.
16) Abell, B.M., Rabu, C., Leznicki, P., Young, J.C., & High, S. (2007) Post-translational integration of tail-anchored proteins is facilitated by defined molecular chaperones. J. Cell Sci., 120, 1743–1751.
17) Abell, B.M., Pool, M.R., Schlenker, O., Sinning, I., & High, S. (2004) Signal recognition particle mediates post-translational targeting in eukaryotes. EMBO J., 23, 2755–2764.
18) Stefanovic, S. & Hegde, R.S. (2007) Identification of a targeting factor for posttranslational membrane protein insertion into the ER. Cell, 128, 1147–1159.
19) Favaloro, V., Spasic, M., Schwappach, B., & Dobberstein, B. (2008) Distinct targeting pathways for the membrane insertion of tail-anchored (TA) proteins. J. Cell Sci., 121, 1832–1840.
20) Schuldiner, M., Metz, J., Schmid, V., Denic, V., Rakwalska, M., Schmitt, H.D., Schwappach, B., & Weissman, J.S. (2008) The GET complex mediates insertion of tail-anchored proteins into the ER membrane. Cell, 134, 634–645.
21) Mariappan, M., Li, X., Stefanovic, S., Sharma, A., Mateja, A., Keenan, R.J., & Hegde, R.S. (2010) A ribosome-associating factor chaperones tail-anchored membrane proteins. Nature, 466, 1120–1124.
22) Shao, S., Rodrigo-Brenni, M.C., Kivlen, M.H., & Hegde, R.S. (2017) Mechanistic basis for a molecular triage reaction. Science, 355, 298–302.
23) O’Neil, K.T. & DeGrado, W.F. (1990) How calmodulin binds its targets:sequence independent recognition of amphiphilic alpha-helices. Trends Biochem. Sci., 15, 59–64.
24) Mizushima, N. (2018) A brief history of autophagy from cell biology to physiology and disease. Nat. Cell Biol., 20, 521–527.
25) Mizushima, N., Yoshimori, T., & Ohsumi, Y. (2011) The role of Atg proteins in autophagosome formation. Annu. Rev. Cell Dev. Biol., 27, 107–132.
26) Itakura, E. & Mizushima, N. (2010) Characterization of autophagosome formation site by a hierarchical analysis of mammalian Atg proteins. Autophagy, 6, 764–776.
27) Itakura, E. & Mizushima, N. (2011) p62 Targeting to the autophagosome formation site requires self-oligomerization but not LC3 binding. J. Cell Biol., 192, 17–27.
28) Itakura, E., Kishi-Itakura, C., Koyama-Honda, I., & Mizushima, N. (2012) Structures containing Atg9A and the ULK1 complex independently target depolarized mitochondria at initial stages of Parkin-mediated mitophagy. J. Cell Sci., 125, 1488–1499.
29) Itakura, E., Kishi-Itakura, C., & Mizushima, N. (2012) The hairpin-type tail-anchored SNARE syntaxin 17 targets to autophagosomes for fusion with endosomes/lysosomes. Cell, 151, 1256–1269.
30) Jiang, P., Nishimura, T., Sakamaki, Y., Itakura, E., Hatta, T., Natsume, T., & Mizushima, N. (2014) The HOPS complex mediates autophagosome-lysosome fusion through interaction with syntaxin 17. Mol. Biol. Cell, 25, 1327–1337.
31) Hamasaki, M., Furuta, N., Matsuda, A., Nezu, A., Yamamoto, A., Fujita, N., Oomori, H., Noda, T., Haraguchi, T., Hiraoka, Y., et al. (2013) Autophagosomes form at ER-mitochondria contact sites. Nature, 495, 389–393.
32) Arasaki, K., Shimizu, H., Mogari, H., Nishida, N., Hirota, N., Furuno, A., Kudo, Y., Baba, M., Baba, N., Cheng, J., et al. (2015) A role for the ancient SNARE syntaxin 17 in regulating mitochondrial division. Dev. Cell, 32, 304–317.
33) Hessa, T., Sharma, A., Mariappan, M., Eshleman, H.D., Gutierrez, E., & Hegde, R.S. (2011) Protein targeting and degradation are coupled for elimination of mislocalized proteins. Nature, 475, 394–397.
34) Rodrigo-Brenni, M.C., Gutierrez, E., & Hegde, R.S. (2014) Cytosolic quality control of mislocalized proteins requires RNF126 recruitment to Bag6. Mol. Cell, 55, 227–237.
35) Mateja, A., Paduch, M., Chang, H.Y., Szydlowska, A., Kossiakoff, A.A., Hegde, R.S., & Keenan, R.J. (2015) Protein targeting. Structure of the Get3 targeting factor in complex with its membrane protein cargo. Science, 347, 1152–1155.
36) Schmidt, O., Pfanner, N., & Meisinger, C. (2010) Mitochondrial protein import:from proteomics to functional mechanisms. Nat. Rev. Mol. Cell Biol., 11, 655–667.
37) Itakura, E., Zavodszky, E., Shao, S., Wohlever, M.L., Keenan, R.J., & Hegde, R.S. (2016) Ubiquilins Chaperone and Triage Mitochondrial Membrane Proteins for Degradation. Mol. Cell, 63, 21–33.
38) Deng, H.X., Chen, W., Hong, S.T., Boycott, K.M., Gorrie, G.H., Siddique, N., Yang, Y., Fecto, F., Shi, Y., Zhai, H., et al. (2011) Mutations in UBQLN2 cause dominant X-linked juvenile and adult-onset ALS and ALS/dementia. Nature, 477, 211–215.
39) Guna, A. & Hegde, R.S. (2018) Transmembrane Domain Recognition during Membrane Protein Biogenesis and Quality Control. Curr. Biol., 28, R498–R511.
40) Hegde, R.S. (2014) Msp1:patrolling mitochondria for lost proteins. EMBO J., 33, 1509–1510.
著者紹介Author Profile
 板倉 英祐(いたくら えいすけ)
板倉 英祐(いたくら えいすけ)千葉大学大学院理学研究院生物学研究部門助教.博士(理学).
略歴2009年埼玉大学大学院理工学研究科博士課程修了,同年東京医科歯科大学大学院医歯学総合研究科研究員,12年英国MRC Laboratory of Molecular Biology研究員を経て,15年より千葉大学大学院助教.
研究テーマと抱負細胞内を掃除するタンパク質品質管理システムの研究.
ウェブサイトhttps://chibau-cellbiology.jimdofree.com/
趣味インドホシガメ(亀吾郎)の飼育と,カメを他人に勧めること.