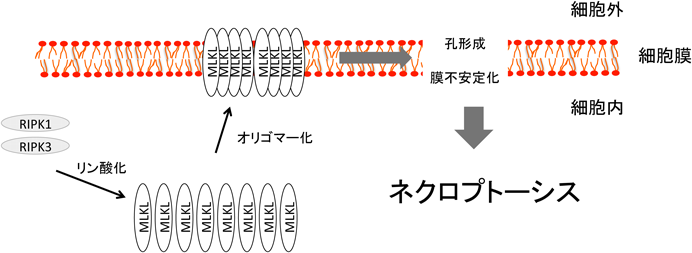
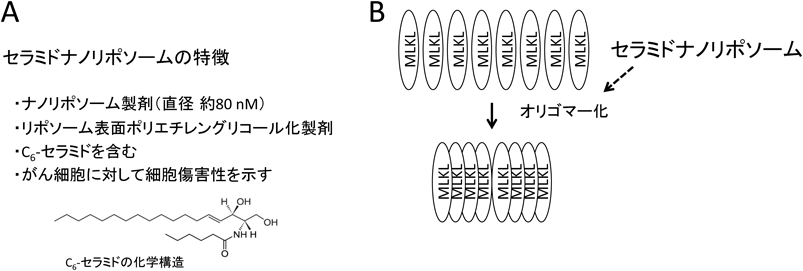
脂質を介したネクロプトーシス制御Regulation of necroptosis by lipids
摂南大学薬学部薬効薬理学Laboratory of Immunopharmacology, Faculty of Pharmaceutical Sciences, Setsunan University ◇ 〒573–0101 大阪府枚方市長尾峠町45–1 ◇ 45–1 Nagaotouge-cho, Hirakata, Osaka 573–0101, Japan
発行日:2018年12月25日Published: December 25, 2018