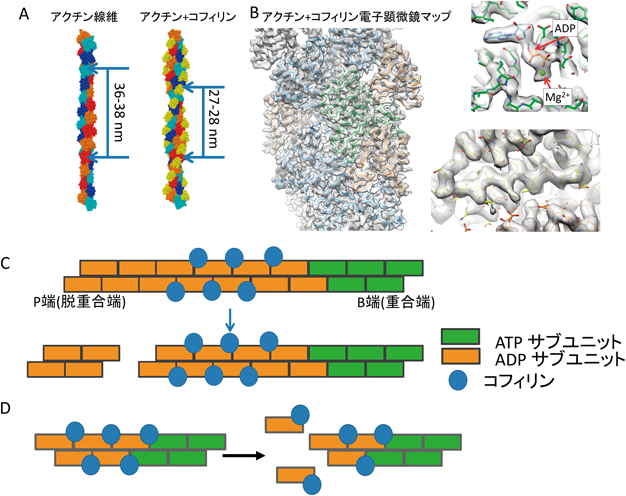
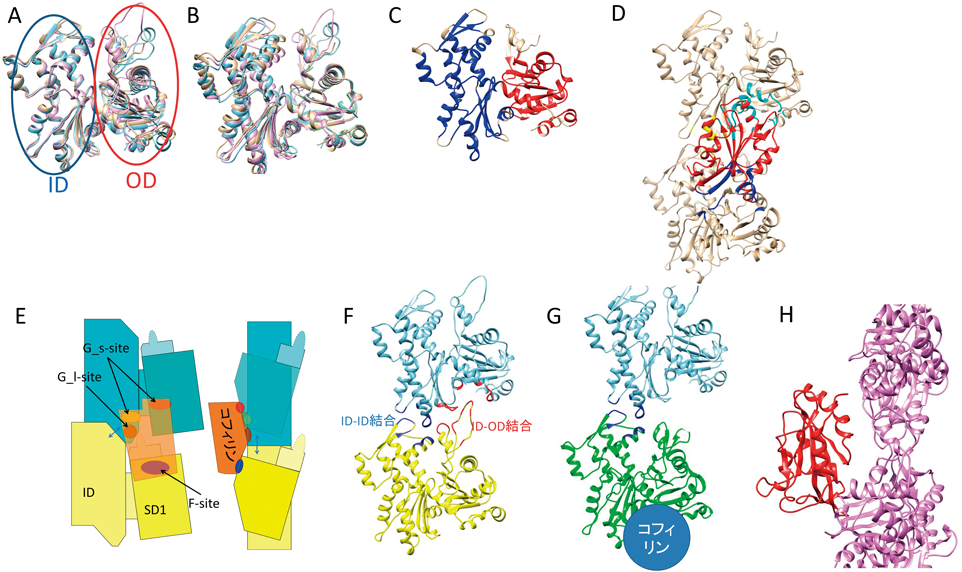
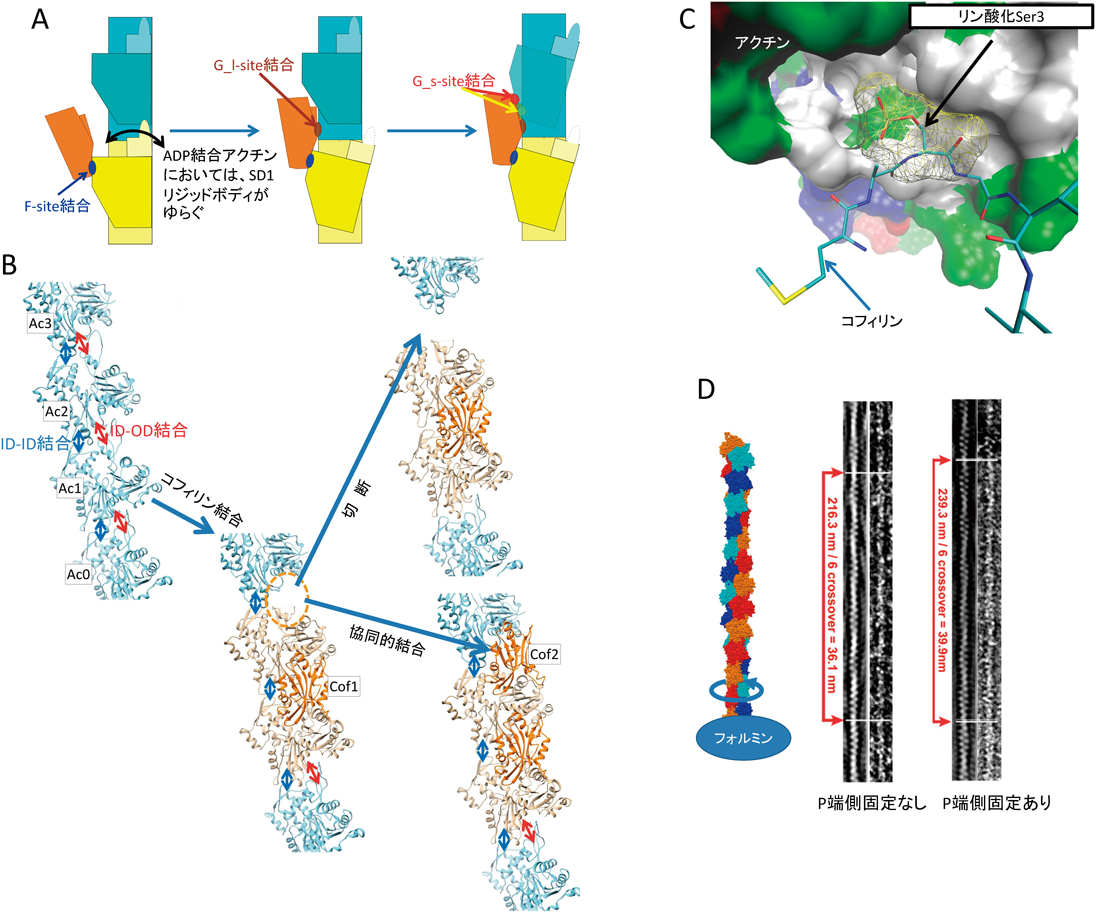
コフィリンによるアクチン線維切断とその制御Severing actin filaments by cofilin and its regulation
名古屋大学大学院理学研究科構造生物学研究センターStructural Biology Research Center, Graduate School of Science, Nagoya University ◇ 名古屋市千種区不老町 ◇ Furo-cho, Chikusa-ku, Nagoya, Japan
発行日:2019年2月25日Published: February 25, 2019