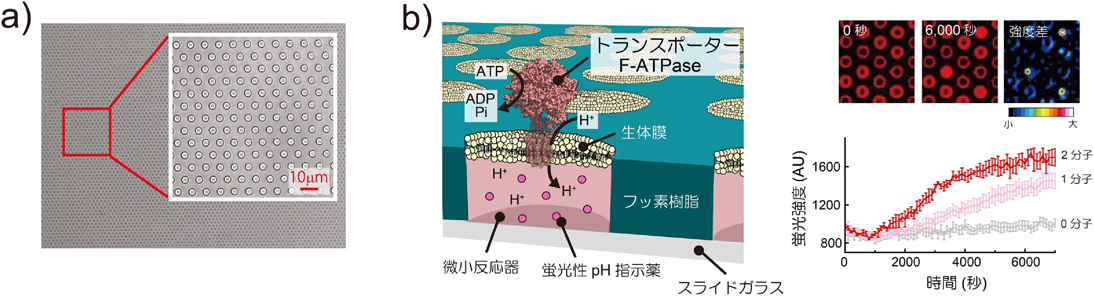
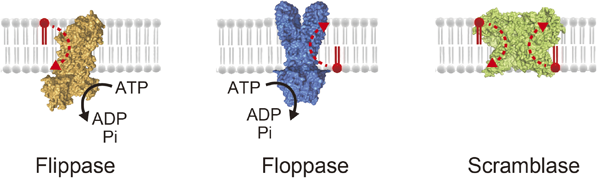
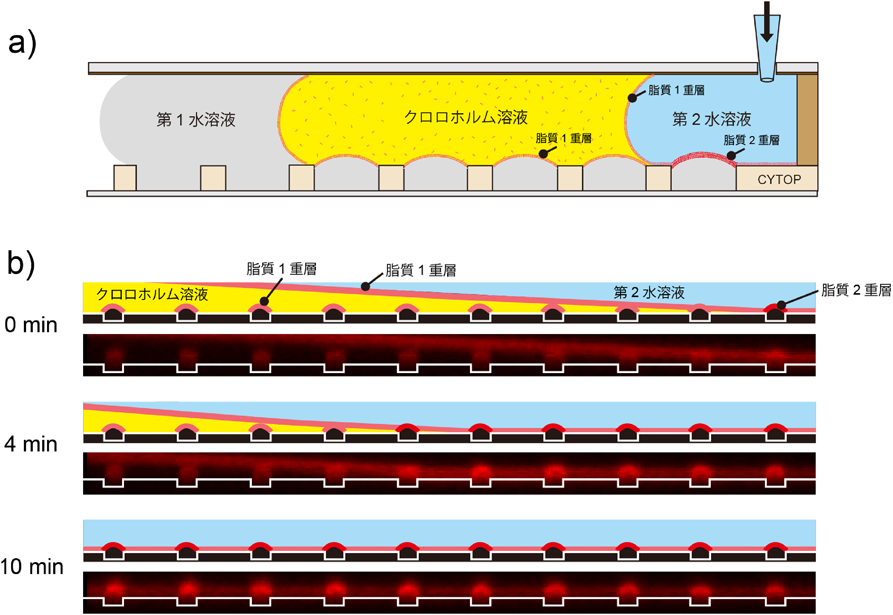
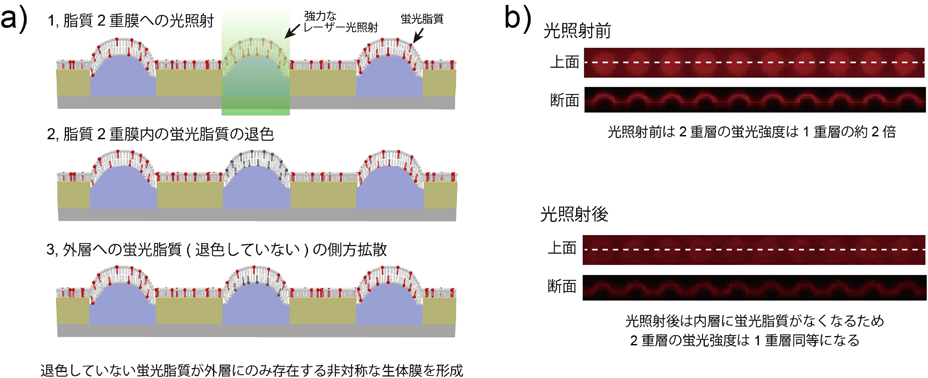
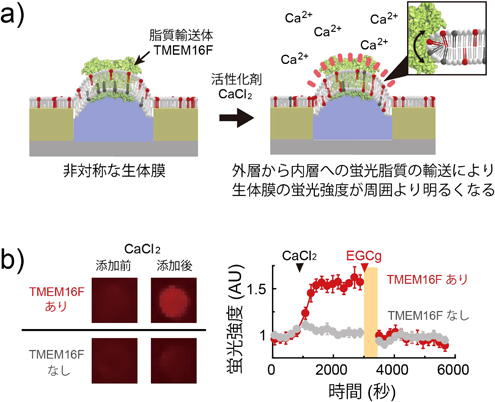
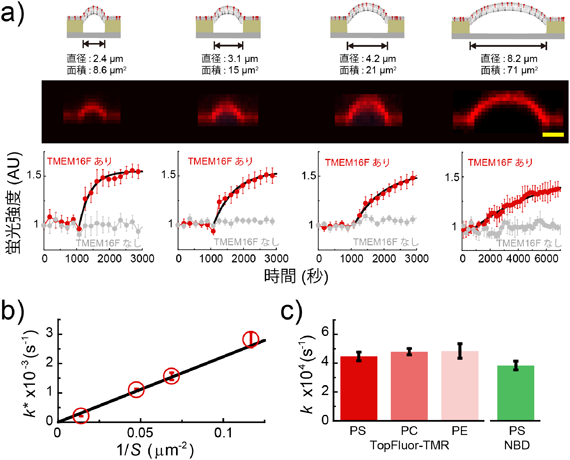
マイクロチップが実現した脂質輸送体の1分子計測Microsystems allow single molecule analysis of phospholipid transport protein
理化学研究所・渡邉分子生理学研究室Molecular Physiology Laboratory, RIKEN ◇ 〒351–0198 埼玉県和光市広沢2–1 ◇ 2–1 Hirosawa, Wako, Saitama 351–0198, Japan
発行日:2019年4月25日Published: April 25, 2019