脳内D-セリンの合成経路は長らく不明であったが,L-セリンからの異性化反応を触媒する酵素としてセリンラセマーゼ(SRR)が1999年に初めて遺伝子クローニングされた2).SRRを培養細胞に発現させると,L-セリン濃度依存的に培地中のD-セリン濃度が増加することから,SRRがD-セリンの合成を担う酵素であることが明らかになった.しかし,SRRはL-セリンの異性化反応だけでなく,D-セリンおよびL-セリンのデヒドラターゼ反応(α, β脱離)も触媒することが明らかになり3),SRRが哺乳類の脳内D-セリンの合成にどの程度関与するかについては結論に至っていなかった.そこで我々は,SRR遺伝子欠損マウス(SRRKO)を作製し,成体脳におけるD-セリン量を野生型(WT)マウスと比較検討した4, 5).その結果,SRRKOマウスの大脳皮質と海馬におけるD-セリン量がWTマウスの10%しかないことが判明した.その後,他の研究グループで作製されたSRRKOマウスにおいても顕著な脳内D-セリン量の減少が認められ,前脳部位では約80~90%のD-セリンがSRRにより合成されると考えられている6, 7).残り10%の脳内D-セリンについては,腸内細菌で産生されるものと食物由来のものの可能性が示唆されているが,まだ実証されていない.
SRRは,最初はglial fibrillary acidic protein(GFAP)陽性のグリア細胞であるアストロサイトに発現すると報告された8).しかし,SRRKOマウス脳を陰性コントロールとして行った免疫組織化学解析では,SRRは神経細胞以外の細胞種では検出されないことが報告され4, 9, 10),生理的状態の脳では,SRRは主に神経細胞に発現することが広く認識されている.
哺乳類の脳内D-セリンの分解酵素としてD-アミノ酸酸化酵素(D-amino acid oxidase:DAO)が知られている11).しかし,脳内DAOの分布はD-セリンの分布と逆相関し,小脳,脊髄などの部位では多く,D-セリンが高濃度に分布している前脳部では少ない12).さらに,DAO遺伝子欠損マウスでは,WTマウスでD-セリンのレベルが低い小脳,延髄でD-セリン量が顕著に増加することから,DAOは後部脳のD-セリンの分解に関わることが考えられる11).一方,前脳部位に高発現するSRRはD-セリンの合成だけでなくD-セリン,L-セリンのデヒドラターゼ反応も触媒し,D-セリンからピルビン酸を産生していることが報告されている3).このことから,前脳部位のD-セリンの一部はSRRによって分解されると考えられている.
3. 興奮性神経伝達とシナプス可塑性の制御におけるD-セリンの役割
脳の神経細胞はシナプスを介して情報伝達を行い,シナプスでの情報の伝わりやすさは経験に依存して動的に変化する.このような「シナプス可塑性」と呼ばれる現象は学習・記憶の細胞学的基盤として考えられており,海馬歯状回で最初に報告された.海馬歯状回の神経細胞に入力する貫通線維に電気的にテタヌス刺激(100 Hzの高頻度刺激)を加えると,シナプス伝達効率の増強が数時間~数週間持続する現象[長期増強(long-term potentiation:LTP)]が観察された.その後,海馬のCA3-CA1シナプス,視床-扁桃体外側核(LA)シナプスにおいても,入力線維に100 Hzやシータバースト刺激(TBS)などの短い高頻度刺激を加えることでLTPが誘導されることが報告された.これらのシナプスでは,1 Hzの低頻度刺激を与えると,シナプス伝達効率の低下が持続する現象が観察され,長期抑圧(long-term depression:LTD)と呼ばれている.
海馬や扁桃体におけるLTPの誘導にはNMDARの活性化が必須である.NMDARは,シナプス前終末から放出されるグルタミン酸による刺激と,シナプス後膜の脱分極が同時に起こったときに活性化され,シナプス前終末とシナプス後膜の同期的な神経活動の検出器として機能し,シナプス後膜でCa2+の流入を起こす.この細胞内Ca2+の上昇が各種のリン酸化酵素を活性化することによりLTPが誘導され,発現することが報告されている.海馬や扁桃体のNMDARは,GRIN1とGRIN2サブユニットが組み合わさり非常に活性の高いイオンチャネルを形成するが,チャネルの十分な活性化には,GRIN2サブユニットにグルタミン酸が結合すると同時に,GRIN1サブユニットのグリシン結合部位にグリシンあるいはD-セリンが結合する必要がある.NMDARグリシン結合部位の飽和状態は日内変動があるものの,ほとんどの時間帯では飽和されない状態にあるため,シナプス間隙におけるD-セリンあるいはグリシン量の変動でNMDARの活性化レベルがダイナミックに制御される可能性が考えられる13).神経活動依存的にD-セリンがシナプス間隙に放出される機構についてはいまだに結論に至ってないが,以下に述べる二つの仮説が主に議論されている.
一つは,Ca2+濃度に依存してアストロサイトがD-セリンを放出し,NMDAR依存性のシナプス伝達を調節するとの仮説であり,次にあげるような研究報告により支持されている.
1)培養アストロサイトを用いたin vitro実験系において,α-アミノ-3-ヒドロキシ-5-メチル-4-イソオキサゾールプロピオン酸(AMPA)型グルタミン酸受容体(AMPAR)の活性化による細胞内Ca2+濃度上昇後,培地中に分泌されるD-セリン量が増加する14, 15).
2)アストロサイトには神経伝達物質放出に関与するSNARE(soluble NSF attachment protein receptor)タンパク質であるVAMP2, VAMP3が局在するが,VAMP2とVAMP3を分解し神経伝達物質のエキソサイトーシス(exocytosis)を抑制する破傷風毒素(TeNT),または小胞への神経伝達物質の取り込みを抑制する抗生物質(concanamycin A)をアストロサイトの培養系に投与すると,D-セリンの放出が抑制される15).
3)視床下部視索上核(SON)を含む脳スライスを用いた電気生理学的解析では,シナプスを取り巻くアストロサイトが多い未出産ラット(1~3か月齢)において,シナプス後細胞の脱分極と入力線維への刺激を同時に与えることにより誘導されるLTPがD-セリンの分解酵素であるDAO処理により完全に消失し,D-セリン投与により回復する.一方,シナプスを取り巻くアストロサイトが少ない7~10日間授乳しているラットでは,DAO処理による影響が少ない16).
4)ラットの海馬スライスを用いた実験系で,アストロサイトのCa2+ clampingによりD-セリンの放出を抑制すると,CA3-CA1シナプスにおけるLTPが障害される17).
5)アストロサイト内のCa2+濃度の制御に関わるTRAP1遺伝子ノックアウトマウスにおいて,海馬CA3-CA1シナプスで誘導されるLTPが顕著に減弱し,D-セリン投与により回復する18).
6)海馬のCA3-CA1シナプスではアストロサイトCB1受容体の活性化後,シナプス部位で利用できるD-セリン量が増加する19).
一方,もう一つの仮説としてWoloskerらはD-セリンが活動依存的に神経細胞からも放出され,NMDARを介する興奮性シナプス伝達とシナプス可塑性を制御するとの考えを提唱している.神経細胞ではシナプス前終末,樹状突起と細胞体においてD-セリンに高親和性を示すアミノ酸輸送体としてナトリウム非依存性alanine-serine-cysteine transporter-1(Asc-1)が同定されており,D-セリンの放出と再取り込みに関与することが示唆されている20).海馬スライスにAsc-1依存的にD-セリンの放出を促進するD-イソロイシンを投与すると,誘導されるLTPが増大するが,D-セリンを分解する細菌由来の酵素D-セリンデヒドラターゼ(DsdA)を投与するとこの効果が消失する.また,Asc-1ノックアウトマウスでは細胞外D-セリン量の減少が認められており,持続的なD-セリン放出にAsc-1が関与する可能性が示唆された21).以上の所見から,神経細胞とアストロサイト両方から神経活動依存的にD-セリンが放出される可能性が考えられるが,それぞれの細胞由来のD-セリンがNMDAR依存性のシナプス伝達とシナプス可塑性の制御にどの程度貢献するかについては今後の検討を必要とする.
海馬のCA1領域では,NMDARを構成するGRIN2AとGRIN2Bサブユニットの分布がシナプス領域とシナプス外領域で異なり,GRIN2Aサブユニットがシナプス間隙に面したシナプス膜領域に多く発現するのに対して,GRIN2Bはシナプス外の膜領域に多く発現することが報告されている.このようなシナプスとシナプス外の膜領域におけるGRIN2AとGRIN2Bサブユニットの分布の違いは,NMDARのコ・アゴニストであるグリシンとD-セリンの量と相関することが示唆されている22).シナプス間隙におけるグリシンはアストロサイトに発現するグリシントランスポーターにより積極的に細胞内に取り込まれるため,シナプス間隙ではグリシン濃度が低く,D-セリンの濃度が高い.D-セリンはGRIN1/GRIN2Aヘテロ四量体で構成されるNMDARに対し高い親和性を持っており,グリシンはGRIN1/GRIN2Bヘテロ四量体で構成されるNMDARに対し高い親和性を示している.最近,量子ドットを用いて培養海馬神経細胞のNMDARの動態を調べた研究で,グリシンとD-セリンがそれぞれGRIN2Aを含むNMDARとGRIN2Bを含むNMDARのシナプス膜上の側方拡散を抑制することが報告され,D-セリンとグリシンがNMDARのシナプス局在のダイナミクスの制御において重要な役割を果たすことが示唆された23).
NMDAR依存性の興奮性シナプス伝達ならびにシナプス可塑性の誘導・発現にNMDARの二つのコ・アゴニストがどのように関与しているかは,D-セリンとグリシンの分解酵素を用いて主に検討されてきた.D-セリンの分解酵素であるDAOあるいはDsdAを海馬スライス標本に灌流すると,NMDAR依存性の興奮性シナプス後電流が減少し,LTPが減弱する.このような変化は,グリシンの分解酵素であるグリシンオキシダーゼ(GO)を投与しても観察されないことから,海馬のNMDAR依存性の興奮性シナプス伝達ならびにシナプス可塑性誘導において,D-セリンが主要なコ・アゴニストとしてNMDARの活性化レベルを制御することが示唆された24).D-セリンが興奮性シナプス伝達とシナプス可塑性において重要な役割を果たすことは,多くの遺伝子改変マウスを用いた研究でも報告されている.脳内D-セリン含量がWTマウスに比べ有意に高いDAO遺伝子欠損マウスでは,NMDAR依存性の神経伝達が促進される25).一方,脳内D-セリン量がWTマウスの10~20%しかないSRRKOマウスでは,海馬のCA3-CA1シナプス,貫通線維–歯状回シナプスにおいて微小興奮性シナプス後電流(mEPSCs)の振幅の減少とLTP障害が報告されている6, 7, 26).また,神経細胞特異的SRRKOマウスでは,弱いLTP誘導条件ではLTPの障害が認められるが,強いLTP誘導条件では対照マウスと同程度のLTPが誘導されることが報告された9).
海馬のシナプスと異なり,扁桃体の視床–LAシナプスでは神経細胞の活動レベルが低い状態ではD-セリン,活動レベルが高い条件ではグリシンがNMDARの主要なコ・アゴニストとして機能することが示唆されている27).D-セリンの分解酵素であるDAOを扁桃体スライスに与えると,視床–LAシナプスで記録される自発的興奮性シナプス後電流の振幅が減少するが,このような変化は,グリシンの分解酵素であるGOを投与すると観察されない.一方,LAへの入力線維を刺激することで誘導されるシナプス電流は,GO処理で減少するがDAOを与えても変化が認められない.これらの結果から,扁桃体神経細胞の活性化レベルに依存して,視床–LAシナプスでは2種類のコ・アゴニストが使い分けられている可能性が初めて示唆された.
4. 発達期の神経回路形成におけるD-セリンの役割
マウスSRRタンパク質の発現は脳の発達に伴い変化し,脳部位によって異なる発現パターンを示すことが報告されている28).大脳皮質および海馬では,生後7日から徐々に発現量が増加し,生後28日目には成体レベルまで達する.一方小脳では,生後14日から28日まで一過性に発現が増加した後,急速に減少する.このようなSRRタンパク質の発現変化は脳発達過程におけるD-セリンの量および分布変化と一致する29).
小脳の発達過程で,SRRの発現が一過性に増加する時期に,バーグマングリア細胞内に高濃度のD-セリンが検出される.この時期は,小脳プルキンエ細胞と登上線維および平行線維との間でシナプスが形成される時期と一致する.生後11日~17日の小脳では,D-セリンはGRID2サブユニットに結合し,平行線維–プルキンエ細胞シナプスにおけるシナプス可塑性の長期抑圧(LTD)を制御することが報告されている30).さらに,発達期のマウス小脳培養切片をSRR阻害剤またはDAOのいずれかで処理すると,顆粒細胞遊走が60%減少することから,D-セリンがNMDARの活性に依存した小脳顆粒細胞の遊走を制御し,小脳の神経回路の形成において重要な役割を果たすことが示唆された14).
経験や学習により形成される記憶は,獲得直後は不安定であり,固定化過程を経て安定した長期記憶として脳内に保存される.興味深いことに,安定した長期記憶は想起に伴い再び不安定な状態になり,その後再固定化過程を経て強固な記憶となる.固定化された記憶は,消去学習(extinction)および忘却(forgetting)というプロセスによって減弱することも知られている.近年,これらの記憶形成の各プロセスにおけるD-セリンの役割が広範に研究され,その機能が明らかにされつつある.
学習・記憶におけるD-セリンの役割は,空間学習課題であるモリス水迷路試験,情動記憶の検討に用いる恐怖条件づけ課題で多く検討された.脳内D-セリン量がWTマウスに比べ有意に高いDAOノックアウトマウスを用いたモリス水迷路試験では,訓練初期(1~5日目)における課題の学習能力はWTマウスと変わらなかったが,後期(6~14日目)ではWTマウスに比べ有意に促進された.また,プローブテストにおいて,空間記憶の指標となる逃避台のあった位置近傍での滞在時間がDAOノックアウトマウスで有意に長くなることが認められた25).一方,SRRKOマウスでは,訓練中の課題の学習能力はWTマウスと変わらなかったが,プローブテストで顕著な空間記憶の障害が認められた6).これらの結果から,D-セリンが空間記憶の獲得と維持に関与することが示唆された.
痕跡恐怖条件づけ(trace fear conditioning)は,動物に明確な手がかり(cue)となる音と電気ショックを,時間間隔を少し空けて与えることで成立する情動学習課題で,文脈ならびに音依存的恐怖記憶の両方とも海馬の機能を必要とする.SRRKOマウスでは,痕跡恐怖条件づけ24時間後の文脈依存的ならびに音依存的恐怖記憶テストにおいて,恐怖記憶の指標となるすくみ反応の割合(フリージング率)が有意に低下し,これらの異常はD-セリン投与により改善されることが報告された26).この結果に一致して,SRRKOマウスでは文脈依存的恐怖条件づけ(contextual fear conditioning)24時間後のテストでも有意な恐怖記憶低下が認められた31).一方,文脈依存的恐怖条件づけ15分後の短期記憶テストでは,SRRKOマウスはWTマウスと同程度のフリージング率を示すことから,恐怖記憶の固定化過程においてD-セリンが重要な役割を果たすことが示唆された.
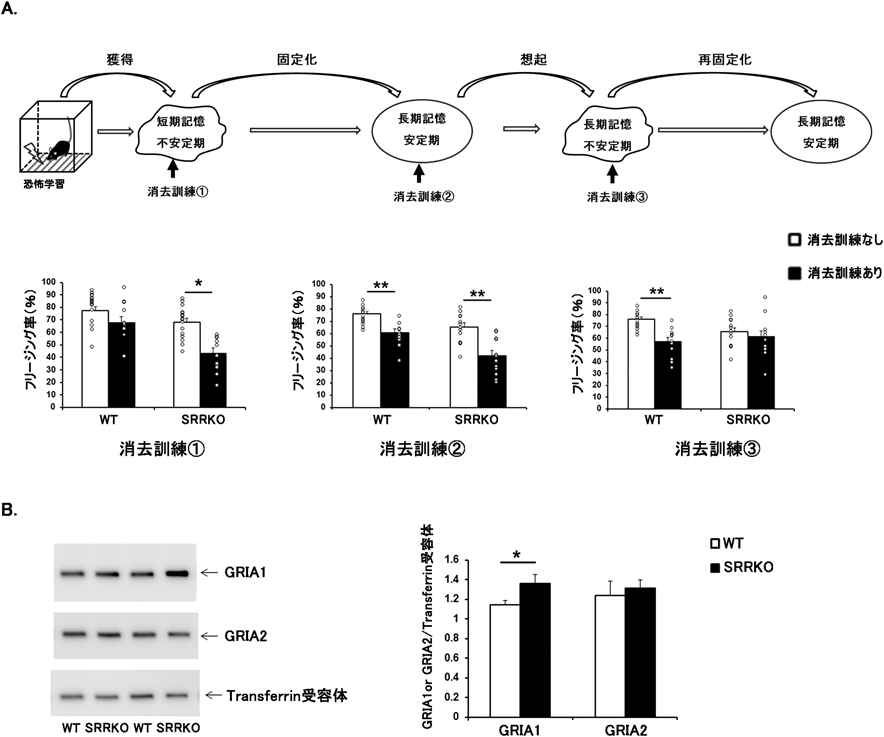
近年,恐怖記憶の消去におけるD-セリンの機能解析が盛んに行われ,多くの知見が蓄積されている.げっ歯類を用いた薬理学的検討では,消去訓練の前にD-セリンを投与すると恐怖記憶の消去が促進されることが,音依存的恐怖条件づけ(auditory cued fear conditioning),抑制性回避学習課題(inhibitory avoidance task),痕跡恐怖条件づけといった恐怖学習課題で報告された32–34).これらの研究は,記憶の固定化過程が終了した安定期に消去訓練を行ったもので,消去された恐怖記憶は時間経過とともに再燃する確率が高いことが示唆されている.最近,記憶の不安定期に消去訓練を行うと恐怖記憶そのものを削除(erasure)することができ,恐怖記憶の再燃が起こりにくいと報告されている35).記憶の形成過程には二つの不安定な時期があり,一つは恐怖条件づけの直後で,もう一つは想起テストの直後である.そこで我々はSRRKOマウスを用いて,記憶の安定期と不安定期にそれぞれ消去訓練を行い,恐怖記憶消去におけるD-セリンの役割を比較検討した31)(図1A).記憶の安定期(文脈依存的恐怖条件づけ24時間後)に消去訓練を行った条件(図1Aの消去訓練②)では,SRRKOマウスはWTマウスと同程度の恐怖反応の低下を示した.記憶の固定化進行中(文脈依存的恐怖条件づけ15分後)に消去訓練(図1Aの消去訓練①)を行うと,SRRKOマウスはWTマウスに比べ恐怖記憶の消去が顕著に促進された.一方,記憶の再固定化進行中(文脈依存的恐怖条件づけ24時間後に3分間の想起テストを行い,その2時間後)に消去訓練(図1Aの消去訓練③)を行うと,WTマウスでは恐怖反応が低下したのに対し,SRRKOマウスでは恐怖反応の低下が認められなかった.恐怖記憶の消去にはAMPARのエンドサイトーシスが関わることが示唆されている.消去訓練後,AMPARがシナプス膜から細胞質内へと移行し,シナプス膜上のAMPARの数が減少し,恐怖記憶が軽減されると考えられている.そこで,再固定化進行中に消去訓練を行い,その1時間後の海馬シナプス膜画分におけるAMPARの発現量をウェスタンブロットで検討したところ,SRRKOマウスはWTマウスに比べAMPARのGRIA1サブユニットの発現が有意に高かった(図1B).SRRKOマウスで認められた再固定化進行中の恐怖記憶消去障害ならびに海馬シナプス膜におけるGRIA1サブユニットの発現上昇はD-セリンを投与することで回復することから,SRRKOマウスではD-セリン量の不足により消去訓練後のAMPARのエンドサイトーシスが抑制され,恐怖記憶の消去に障害が生じた可能性が考えられた.本研究により,D-セリンは記憶の固定化過程では恐怖記憶の消去を抑制し,再固定化過程では逆に恐怖記憶の消去を促進することが示唆され,消去訓練の効果は実施するタイミングに大きく依存することが明らかになった.
7. 神経細胞死を伴う神経疾患におけるD-セリンの役割
D-セリンは神経発達,シナプス可塑性,学習・記憶などの生理機能の制御に重要な役割を果たす一方,脳虚血や脳梗塞,アルツハイマー病(AD),筋萎縮性側索硬化症(ALS)などの多くの疾患の病態形成にも深く関与することが報告されている.つまり,D-セリンは正常状態では神経細胞の機能維持に必要であるが,過剰な働きはNMDARを介する興奮神経毒性を引き起こすと考えられる.虚血や変性疾患に対してきわめて脆弱な脳部位である海馬にNMDAを投与すると神経細胞死が誘導されるが,海馬スライスにD-セリンの分解酵素であるDsdAを投与すると神経細胞死が顕著に抑制されることがShleperらによって報告され,グルタミン酸興奮毒性とD-セリン代謝異常との関連が初めて示された38).その後,我々もSRRKOマウスを用い,海馬へのNMDA注入により引き起こされる脳障害がSRRKOマウスにおいて有意に緩和されることを見いだした5).さらに,SRRKOを用いて中大脳動脈閉塞による局所脳虚血モデルを作製した実験では,脳梗塞の範囲がWTマウスに比べ有意に縮小されることが報告され39),D-セリンがNMDARを介する興奮性神経細胞死に重要な役割を果たすことが明らかになった.
D-セリンの機能異常は急性的な神経細胞死だけでなく,緩慢な神経細胞死を伴う神経変性疾患の病態にも深く関与することがADやALSの動物モデルを用いた研究で示唆されている.ADの発症機序にはβアミロイド(Aβ)の異常蓄積,特にそのオリゴマーの蓄積による初期のシナプス機能障害,そして最終段階の神経細胞死という仮説が広く受け入れられている.D-セリンはシナプス可塑性の調節や神経細胞死の誘導両方に関与していることから,認知機能改善と神経保護という視点から有効な治療ターゲットとして注目され,多くの研究が行われてきた.AβとD-セリンの関連性は,最初は海馬の培養ミクログリア細胞を用いて検討され,Aβを投与するとSRR発現量が増加し,培地中に放出されるD-セリン量が有意に増加することが報告された40).さらに,海馬にAβを慢性的に投与したADモデルマウスにおいてもD-セリン量の増加が認められることから41),Aβの蓄積はD-セリン量の制御異常を引き起こす原因の一つであると考えられる.さらに,海馬にAβを直接注入することにより引き起こされる神経細胞死がSRRKOマウスで有意に緩和されることが我々の研究で確認され,Aβの蓄積により引き起こされるSRR発現とD-セリン量の増加が神経細胞死の誘導に関与することが強く示唆された5).これらの研究結果に一致して,AD患者の死後脳の海馬と頭頂葉でD-セリン量が対照群に比べ増加していることが報告された41).脳脊髄液におけるD-セリン量の増加は,ADの可能性があると診断されている患者においても認められ,最近ではADの新規バイオマーカーの候補としてのD-セリンの妥当性が検討されている.脳脊髄液におけるD-セリン量とAD患者の認知機能との関連性はまだ明らかではないが,D-セリンの役割はADの病態進行に伴い変化すると考えられている.D-セリンは正常なシナプス機能の発揮に必要であり,海馬の神経新生ならびに新生神経細胞の生存を促進することから,ADの病態進行の初期段階では,D-セリンの増加は認知機能促進効果を発揮すると推測されている.実際プラセボ対照二重盲検試験では,DAOの抑制薬である安息香酸ナトリウムが初期段階のAD患者の認知機能を促進することが報告されている.一方,ADの病態進行が中期から末期に入ると,D-セリン量の増加はNMDAR依存性の興奮性神経細胞死の誘導に関与し,認知機能の悪化をもたらす可能性が推測される.したがって,今後はADの病態進行過程におけるD-セリン量の変化を経時的に計測することで,D-セリン動態とADの病態進行度との関連性の解明が期待される.
D-セリンは運動神経疾患の一つであるALSの病態にも深く関与することが示唆されている.ALSモデル動物であるSOD1(superoxide dismutase)-G37R変異マウスでは,運動神経変性の発症後に脊髄におけるSRR発現が上昇し,脊髄前角でD-セリンが蓄積することが報告された42).SRRの発現上昇は運動神経変性の進行に伴い増加する活性化ミクログリアにおいて特に顕著であることから,脊髄前角で蓄積するD-セリンの一部はミクログリア由来と考えられている.また,SOD1-G37R変異マウスでは脊髄全体のDAO酵素活性が低下し,mRNA,タンパク質の発現ともに有意に減少することも報告され43),D-セリンの合成と分解のバランスの崩壊がALSにおける興奮神経毒性を引き起こす原因の一つであると考えられている.
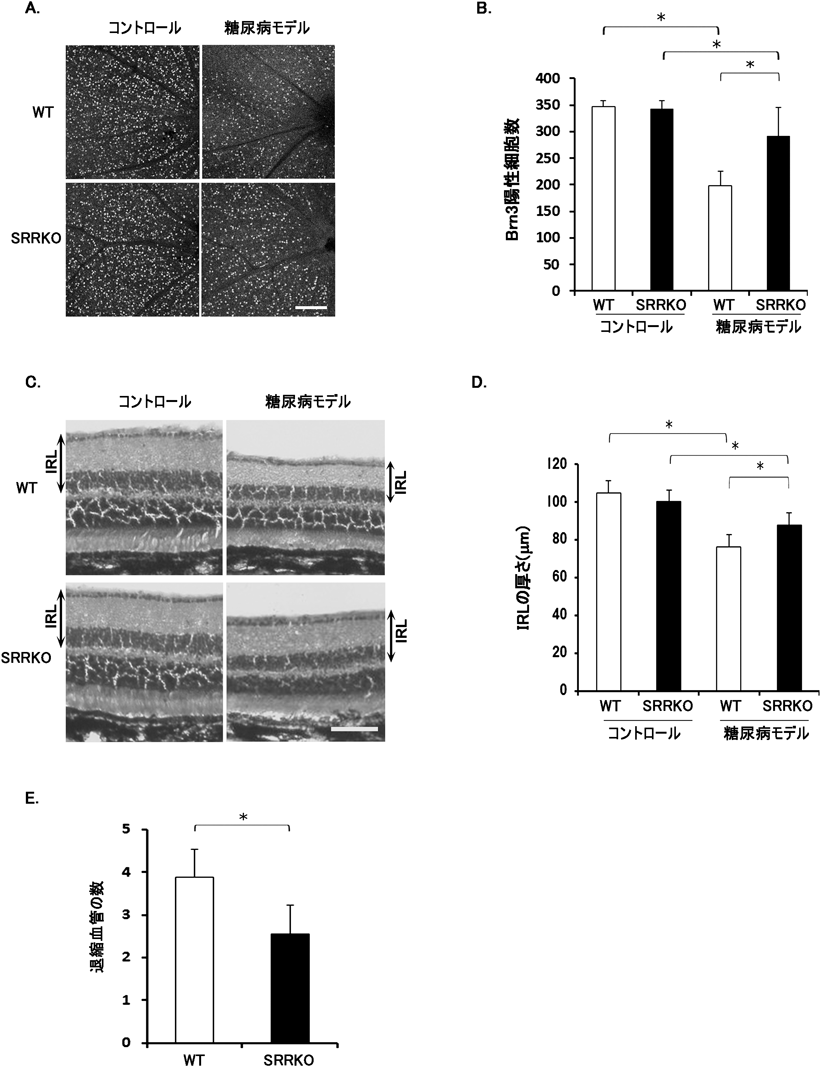
糖尿病網膜症は糖尿病に特徴的な合併症であり,成人の失明の主な原因である.我々の研究室では,SRRKOマウスを用いてI型糖尿病モデルを作製し,糖尿病網膜症の主な病態である網膜の神経変性(網膜神経節細胞数の減少と網膜内層の厚みの減少)ならびに微小血管障害がSRRKOマウスで軽減されることを報告した44)(図2).糖尿病モデルでは,網膜におけるSRR発現が増加し,硝子体液中のD-セリン量が増加することが報告されている45).これらの結果から,網膜D-セリン量の増加によるNMDARの過剰活性化が,網膜の神経変性を引き起こす原因の一つであると考えられる.現時点で,SRRとD-セリンがどのようなメカニズムを介し,糖尿病網膜症における微小血管障害に関与するかは不明である.糖尿病網膜症では神経変性が網膜血管障害に先行して生じる.さらに,網膜の神経節細胞は血小板由来増殖因子(PDGF)-Aを分泌し,PDGF受容体を持つアストロサイトに作用することで血管内皮細胞の生存を促進することから,D-セリン依存的な網膜神経細胞の障害が微小血管障害を悪化させる可能性が考えられる.
引用文献References
1) Hashimoto, A., Nishikawa, T., Hayashi, T., Fujii, N., Harada, K., Oka, T., & Takahashi, K. (1992) The presence of free D-serine in rat brain. FEBS Lett., 296, 33–36.
2) Wolosker, H., Blackshaw, S., & Snyder, S.H. (1999) Serine racemase: A glial enzyme synthesizing D-serine to regulate glutamate-N-methyl-D-aspartate neurotransmission. Proc. Natl. Acad. Sci. USA, 96, 13409–13414.
3) Foltyn, V.N., Bendikov, I., De Miranda, J., Panizzutti, R., Dumin, E., Shleper, M., Li, P., Toney, M.D., Kartvelishvily, E., & Wolosker, H. (2005) Serine racemase modulates intracellular D-serine levels through an α,β-elimination activity. J. Biol. Chem., 280, 1754–1763.
4) Miya, K., Inoue, R., Takata, Y., Abe, M., Natsume, R., Sakimura, K., Hongou, K., Miyawaki, T., & Mori, H. (2008) Serine racemase is predominantly localized in neurons in mouse brain. J. Comp. Neurol., 510, 641–654.
5) Inoue, R., Hashimoto, K., Harai, T., & Mori, H. (2008) NMDA- and beta-amyloid1-42-induced neurotoxicity is attenuated in serine racemase knock-out mice. J. Neurosci., 28, 14486–14491.
6) Basu, A.C., Tsai, G.E., Ma, C.L., Ehmsen, J.T., Mustafa, A.K., Han, L., Jiang, Z.I., Benneyworth, M.A., Froimowitz, M.P., Lange, N., et al. (2009) Targeted disruption of serine racemase affects glutamatergic neurotransmission and behavior. Mol. Psychiatry, 14, 719–727.
7) Labrie, V., Fukumura, R., Rastogi, A., Fick, L., Wang, W., Boutros, P., Kennedy, J., Semeralul, M., Lee, F., Baker, G., et al. (2009) Serine racemase is associated with schizophrenia susceptibility in humans and in a mouse model. Hum. Mol. Genet., 18, 3227–3243.
8) Schell, M.J., Molliver, M.E., & Snyder, S.H. (1995) D-serine, an endogenous synaptic modulator: Localization to astrocytes and glutamate-stimulated release. Proc. Natl. Acad. Sci. USA, 92, 3948–3952.
9) Benneyworth, M.A., Li, Y., Basu, A.C., Bolshakov, V.Y., & Coyle, J.T. (2012) Cell selective conditional null mutations of serine racemase demonstrate a predominate localization in cortical glutamatergic neurons. Cell. Mol. Neurobiol., 32, 613–624.
10) Wolosker, H., Balu, D.T., & Coyle, J.T. (2016) The rise and fall of the D-serine-mediated gliotransmission hypothesis. Trends Neurosci., 39, 712–721.
11) Hashimoto, A., Nishikawa, T., Konno, R., Niwa, A., Yasumura, Y., Oka, T., & Takahashi, K. (1993) Free D-serine, D-aspartate and D-alanine in central nervous system and serum in mutant mice lacking D-amino acid oxidase. Neurosci. Lett., 152, 33–36.
12) Moreno, S., Nardacci, R., Cimini, A., & Ceru, M.P. (1999) Immunocytochemical localization of D-amino acid oxidase in rat brain. J. Neurocytol., 28, 169–185.
13) Papouin, T., Dunphy, J.M., Tolman, M., Dineley, K.T., & Haydon, P.G. (2017) Septal cholinergic neuromodulation tunes the astrocyte-dependent gating of hippocampal NMDA receptors to wakefulness. Neuron, 94, 840–854.
14) Kim, P.M., Aizawa, H., Kim, P.S., Huang, A.S., Wickramasinghe, S.R., Kashani, A.H., Barrow, R.K., Huganir, R.L., Ghosh, A., & Snyder, S.H. (2005) Serine racemase: Activation by glutamate neurotransmission via glutamate receptor interacting protein and mediation of neuronal migration. Proc. Natl. Acad. Sci. USA, 102, 2105–2110.
15) Mothet, J.P., Pollegioni, L., Ouanounou, G., Martineau, M., Fossier, P., & Baux, G. (2005) Glutamate receptor activation triggers a calcium-dependent and SNARE protein-dependent release of the gliotransmitter D-serine. Proc. Natl. Acad. Sci. USA, 102, 5606–5611.
16) Panatier, A., Theodosis, D.T., Mothet, J.P., Touquet, B., Pollegioni, L., Poulain, D.A., & Oliet, S.H. (2006) Glia-derived D-serine controls NMDA receptor activity and synaptic memory. Cell, 125, 775–784.
17) Henneberger, C., Papouin, T., Oliet, S.H., & Rusakov, D.A. (2010) Long-term potentiation depends on release of D-serine from astrocytes. Nature, 463, 232–236.
18) Shigetomi, E., Jackson-Weaver, O., Huckstepp, R.T., O’Dell, T.J., & Khakh, B.S. (2013) TRPA1 channels are regulators of astrocyte basal calcium levels and long-term potentiation via constitutive D-serine release. J. Neurosci., 33, 10143–10153.
19) Robin, L.M., Oliveira da Cruz, J.F., Langlais, V.C., Martin-Fernandez, M., Metna-Laurent, M., Busquets-Garcia, A., Bellocchio, L., Soria-Gomez, E., Papouin, T., Varilh, M., et al. (2018) Astroglial CB1 receptors determine synaptic D-serine availability to enable recognition memory. Neuron, 98, 935–944.
20) Rosenberg, D., Artoul, S., Segal, A.C., Kolodney, G., Radzishevsky, I., Dikopoltsev, E., Foltyn, V.N., Inoue, R., Mori, H., Billard, J.M., et al. (2013) Neuronal D-serine and glycine release via the Asc-1 transporter regulates NMDA receptor-dependent synaptic activity. J. Neurosci., 33, 3533–3544.
21) Sason, H., Billard, J.M., Smith, G.P., Safory, H., Neame, S., Kaplan, E., Rosenberg, D., Zubedat, S., Foltyn, V.N., Christoffersen, C.T., et al. (2017) Asc-1 transporter regulation of synaptic activity via the tonic release of D-serine in the forebrain. Cereb. Cortex, 27, 1573–1587.
22) Papouin, T., Ladépêche, L., Ruel, J., Sacchi, S., Labasque, M., Hanini, M., Groc, L., Pollegioni, L., Mothet, J.P., & Oliet, S.H. (2012) Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell, 150, 633–646.
23) Ferreira, J.S., Papouin, T., Ladépêche, L., Yao, A., Langlais, V.C., Bouchet, D., Dulong, J., Mothet, J.P., Sacchi, S., Pollegioni, L., et al. (2017) Co-agonists differentially tune GluN2B-NMDA receptor trafficking at hippocampal synapses. eLife, 6, 25492.
24) Yang, Y., Ge, W., Chen, Y., Zhang, Z., Shen, W., Wu, C., Poo, M., & Duan, S. (2003) Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine. Proc. Natl. Acad. Sci. USA, 100, 15194–15199.
25) Maekawa, M., Watanabe, M., Yamaguchi, S., Konno, R., & Hori, Y. (2005) Spatial learning and long-term potentiation of mutant mice lacking D-amino-acid oxidase. Neurosci. Res., 53, 34–38.
26) Balu, D.T., Li, Y., Puhl, M.D., Benneyworth, M.A., Basu, A.C., Takagi, S., Bolshakov, V.Y., & Coyle, J.T. (2013) Multiple risk pathways for schizophrenia converge in serine racemase knockout mice, a mouse model of NMDA receptor hypofunction. Proc. Natl. Acad. Sci. USA, 110, E2400–E2409.
27) Li, Y., Sacchi, S., Pollegioni, L., Basu, A.C., Coyle, J.T., & Bolshakov, V.Y. (2013) Identity of endogenous NMDAR glycine site agonist in amygdala is determined by synaptic activity level. Nat. Commun., 4, 1760.
28) Wang, L.Z. & Zhu, X.Z. (2003) Spatiotemporal relationships among D-serine, serine racemase, and D-amino acid oxidase during mouse postnatal development. Acta Pharmacol. Sin., 24, 965–974.
29) Schell, M.J., Brady, R.O. Jr., Molliver, M.E., & Snyder, S.H. (1997) D-Serine as a neuromodulator: Regional and developmental localizations in rat brain glia resemble NMDA receptors. J. Neurosci., 17, 1604–1615.
30) Kakegawa, W., Miyoshi, Y., Hamase, K., Matsuda, S., Matsuda, K., Kohda, K., Emi, K., Motohashi, J., Konno, R., Zaitsu, K., et al. (2011) D-serine regulates cerebellar LTD and motor coordination through the δ2 glutamate receptor. Nat. Neurosci., 14, 603–611.
31) Inoue, R., Talukdar, G., Takao, K., Miyakawa, T., & Mori, H. (2018) Dissociated role of D-serine in extinction during consolidation vs. reconsolidation of context conditioned Fear. Front. Mol. Neurosci., 11, 161.
32) Matsuda, S., Matsuzawa, D., Nakazawa, K., Sutoh, C., Ohtsuka, H., Ishii, D., Tomizawa, H., Iyo, M., & Shimizu, E. (2010) D-serine enhances extinction of auditory cued fear conditioning via ERK1/2 phosphorylation in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry, 34, 895–902.
33) Bai, Y., Zhou, L., Wu, X., & Dong, Z. (2014) D-serine enhances fear extinction by increasing GluA2-containing AMPA receptor endocytosis. Behav. Brain Res., 270, 223–227.
34) Balu, D.T., Presti, K.T., Huang, C.C., Muszynski, K., Radzishevsky, I., Wolosker, H., Guffanti, G., Ressler, K.J., & Coyle, J.T. (2018) Serine racemase and D-serine in the amygdala are dynamically involved in fear learning. Biol. Psychiatry, 83, 273–283.
35) Monfils, M.H., Cowansage, K.K., Klann, E., & LeDoux, J.E. (2009) Extinction-reconsolidation boundaries:key to persistent attenuation of fear memories. Science, 324, 951–955.
36) Turpin, F.R., Potier, B., Dulong, J.R., Sinet, P.M., Alliot, J., Oliet, S.H., Dutar, P., Epelbaum, J., Mothet, J.P., & Billard, J.M. (2011) Reduced serine racemase expression contributes to age-related deficits in hippocampal cognitive function. Neurobiol. Aging, 32, 1495–1504.
37) Avellar, M., Scoriels, L., Madeira, C., Vargas-Lopes, C., Marques, P., Dantas, C., Manhães, A.C., Leite, H., & Panizzutti, R. (2016) The effect of D-serine administration on cognition and mood in older adults. Oncotarget, 7, 11881–11888.
38) Shleper, M., Kartvelishvily, E., & Wolosker, H. (2005) D-Serine is the dominant endogenous coagonist for NMDA receptor neurotoxicity in organotypic hippocampal slices. J. Neurosci., 25, 9413–9417.
39) Mustafa, A.K., Ahmad, A.S., Zeynalov, E., Gazi, S.K., Sikka, G., Ehmsen, J.T., Barrow, R.K., Coyle, J.T., Snyder, S.H., & Doré, S. (2010) Serine racemase deletion protects against cerebral ischemia and excitotoxicity. J. Neurosci., 30, 1413–1416.
40) Wu, S.Z., Bodles, A.M., Porter, M.M., Griffin, W.S., Basile, A.S., & Barger, S.W. (2004) Induction of serine racemase expression and D-serine release from microglia by amyloid beta-peptide. J. Neuroinflammation, 1, 2.
41) Madeira, C., Lourenco, M.V., Vargas-Lopes, C., Suemoto, C.K., Brandão, C.O., Reis, T., Leite, R.E., Laks, J., Jacob-Filho, W., Pasqualucci, C.A., et al. (2015) D-serine levels in Alzheimer’s disease: Implications for novel biomarker development. Transl. Psychiatry, 5, e561.
42) Sasabe, J., Chiba, T., Yamada, M., Okamoto, K., Nishimoto, I., Matsuoka, M., & Aiso, S. (2007) D-serine is a key determinant of glutamate toxicity in amyotrophic lateral sclerosis. EMBO J., 26, 4149–4159.
43) Sasabe, J., Miyoshi, Y., Suzuki, M., Mita, M., Konno, R., Matsuoka, M., Hamase, K., & Aiso, S. (2012) D-amino acid oxidase controls motoneuron degeneration through D-serine. Proc. Natl. Acad. Sci. USA, 109, 627–632.
44) Ozaki, H., Inoue, R., Matsushima, T., Sasahara, M., Hayashi, A., & Mori, H. (2018) Serine racemase deletion attenuates neurodegeneration and microvascular damage in diabetic retinopathy. PLoS One, 13, e0190864.
45) Jiang, H., Fang, J., Wu, B., Yin, G., Sun, L., Qu, J., Barger, S.W., & Wu, S. (2011) Overexpression of serine racemase in retina and overproduction of D-serine in eyes of streptozotocin-induced diabetic retinopathy. J. Neuroinflammation, 8, 119.
46) Gómez-Galán, M., De Bundel, D., Van Eeckhaut, A., Smolders, I., & Lindskog, M. (2013) Dysfunctional astrocytic regulation of glutamate transmission in a rat model of depression. Mol. Psychiatry, 18, 582–594.
47) Malkesman, O., Austin, D.R., Tragon, T., Wang, G., Rompala, G., Hamidi, A.B., Cui, Z., Young, W.S., Nakazawa, K., Zarate, C.A., et al. (2012) Acute D-serine treatment produces antidepressant-like effects in rodents. Int. J. Neuropsychopharmacol., 15, 1135–1148.
48) Ishiwata, S., Hattori, K., Sasayama, D., Teraishi, T., Miyakawa, T., Yokota, Y., Matsumura, R., Nishikawa, T., & Kunugi, H. (2018) Cerebrospinal fluid D-serine concentrations in major depressive disorder negatively correlate with depression severity. J. Affect. Disord., 226, 155–162.
49) Dong, C., Zhang, J.C., Ren, Q., Ma, M., Qu, Y., Zhang, K., Yao, W., Ishima, T., Mori, H., & Hashimoto, K. (2018) Deletion of serine racemase confers D-serine-dependent resilience to chronic social defeat stress. Neurochem. Int., 116, 43–51.