衛生環境の優れる日本において,ほとんど意識することはないが,世界的にはコレラや赤痢といった病原菌の感染者はいまだに相当数に上り,毎年多くの犠牲者を出している.最近の例としては,2010年1月に大地震に見舞われたハイチでコレラ患者が激増し,感染者だけで20万人以上,死者は5千人を超え,長期間にわたって感染に歯止めが掛からない状態が続いた.
本稿では,このような病原菌の蔓延に対応するため,細菌の生存に必須な元素である鉄に注目した.エネルギーを生産する呼吸鎖を構成するタンパク質の多くがヘム(図1)や鉄–硫黄クラスター等を補因子として有するため,細菌が増殖する際に鉄を必要とする.人の体内に存在する鉄の約70%はヘモグロビン中にヘムとして存在するため,ヘムが病原菌の格好の鉄源となる.そのため,細菌の鉄の補給路を断つことは,細菌の増殖を抑え,人体への影響を軽減するための有効な手段になると期待される.
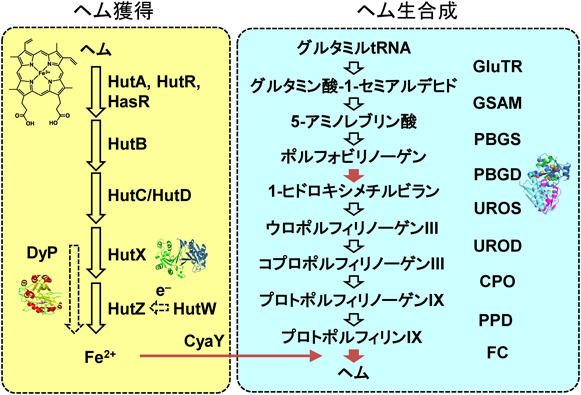
コレラ菌のゲノム配列と他の生物の遺伝子情報との比較から,Hut(heme utilization protein)と呼ばれる一群のタンパク質がヘムを通じて鉄の取り込みに関与していると予想された(図1)1).HutA, HutR, HasRという外膜タンパク質がヘモグロビンからヘムを取り込み,HutBという可溶性タンパク質が膜間部においてヘムを輸送し,内膜タンパク質であるHutC/HutD複合体を通り,細胞質内に取り込まれ,HutZがヘムを分解し,鉄を取り出すという流れであるが,タンパク質レベルでの研究はほとんど行われていなかった.
1)ヘム分解酵素HutZ
ヘムは安定な鉄–ポルフィリン錯体であるため,ヘムから鉄を取り出す反応は容易ではない.ヒトの場合,赤血球から放出されたヘモグロビンはヘムとタンパク質に分けられ,ヘムはヘム分解酵素により分解され,鉄が取り出され,再利用される.大腸菌やビフィズス菌,乳酸菌といった人体に無害な細菌にはヘム分解酵素と想定される遺伝子が存在しないものが多いが,結核菌や赤痢菌,ピロリ菌,大腸菌O157といった病原菌の多くがヘム分解酵素を持つことがわかった.遺伝子解析の結果,コレラ菌にピロリ菌のヘム分解酵素と相同性の高い遺伝子(hutZ)を発見したので,このタンパク質のヘム分解能を調べた.
ヘム分解酵素はヒト型,黄色ブドウ球菌型,ピロリ菌型の3種類におおよそ分けられる.これらのグループ間でアミノ酸配列と立体構造に相同性はなく,反応機構もヒト型と黄色ブドウ球菌型では異なることが明らかにされていたが2),ピロリ菌型の反応機構は明らかにされていなかった.我々は,時間分解スペクトルの測定等から,HutZが,ヒト型の酵素がヘムを分解するときに観測される中間体と同じ中間体を経由し,ヘムを分解することを明らかにした3).
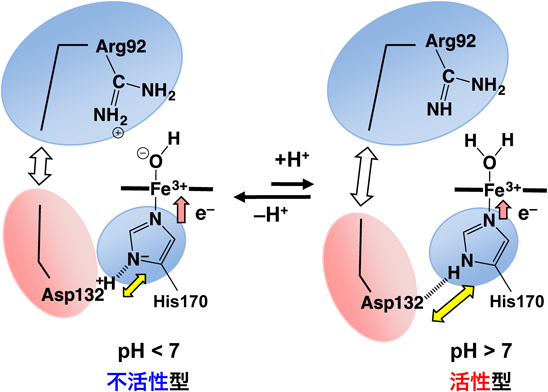
HutZによるヘム分解反応は酸性条件では反応が進行するが,中性付近では反応しないというpH依存性を示した4).中性付近ではヘム分解反応の最初の過程であるヘム鉄の還元が遅いことが反応が進行しない原因であることがわかった.これは,ヘムと結合するヒスチジンが周囲のアミノ酸残基と水素結合を形成した結果,ヒスチジンが分極し,酸化型のヘムをより安定化させるためである(図2).このような水素結合はヒト型の酵素には存在しないことから,HutZを含むピロリ菌型の特徴であると考えられる.つまり,HutZはヘムの分解反応の初期過程において,反応を進行させないような構造をしている.コレラ菌は,海や川など増殖に適さない環境に存在する場合,酵素活性を抑制し,エネルギーをむだに消費しない状態で存在しているのに対し,いったんヒトの体内に侵入し,栄養が豊富で増殖が可能な状況に置かれると,酵素を活性化させ急激に増殖する,というように周囲の環境に応じて,水素結合という弱い結合を利用することで,酵素活性を調節していると考えられる.
我々は,コレラ患者の第一選択薬として知られるテトラサイクリンを添加すると,HutZによるヘムの分解反応が著しく遅くなることを発見した5).テトラサイクリンはクロラムフェニコール等の抗生物質と異なり,金属キレート能があり,HutZの阻害効果は金属キレート剤であるデフェロキサミン,フェロジン,クエン酸の添加によっても観測された.素過程の反応速度の比較から,キレート剤は還元された酸素結合体へのプロトン付加を阻害し,効果的な阻害剤として機能することを見いだした.
2)ヘム輸送タンパク質:HutX, HutB
HutZがヘム分解酵素であることを明らかにしたが,基質であるヘムをどのようにして獲得するのかは不明であった.HutZの遺伝子はhutW-hutX-hutZというオペロンを形成している.このうち,HutWは鉄–硫黄クラスター配列を有することからHutZが行うヘム分解反応へ電子を供与するタンパク質と予想されるが,タンパク質の発現に成功していない.一方,HutXはヘム結合タンパク質として報告されていたChuXと相同性が58%と高いことから6),HutXがHutZにヘムを輸送するタンパク質である可能性について検討した.
HutXからHutZへのヘムの輸送を吸収スペクトルにより観測したところ,ヘムと結合したHutXからヘムを結合していないHutZへのヘムの移動はヘムの親和性の高いアポミオグロビンに移動するよりも速かった7).HutXとHutZの相互作用の解離定数は400 µMと大きく,弱い結合であったが,これは解離速度定数が4×10−2 s–1と大きいためであり,HutXはHutZにヘムを輸送した後,直ちにHutXから解離するため,輸送タンパク質として理想的であることがわかった.さらに,結晶構造解析とラマンスペクトルから,HutXはチロシンを配位子とし,ヘムと結合することが示唆された7).
チロシンを利用したヘムの輸送は,膜間のヘム輸送タンパク質であるHutB(図1)でも同様であった8).HutBの全長タンパク質は可溶性の凝集体を形成するが,N末端の22残基を除去すると単量体になったことから,N末端にペリプラズムへの移行シグナル配列を持つ.Tyr65, Tyr198, His164をそれぞれ変異させるとヘムの配位構造が変化した.緑膿菌のヘム輸送タンパク質であるHasAではチロシンがヘムに配位し,そのチロシンにヒスチジンが水素結合することで,チロシンの求核性を高めていることが知られているが9),HutBもHasAと同様の配位構造でヘムを結合し,輸送をしていると考えられる.
コレラ菌には上記のヘムの獲得系だけではなく,グルタミルtRNAを原料に9段階の過程からなるヘムの生合成系も存在する(図1).つまり,獲得系により取り込んだヘムを分解し,鉄を取り出す一方,ヘムを合成する.ヘムの獲得系と生合成系が共存することは,一見むだに思え,その理由は不明である.ビフィズス菌や乳酸菌はヘムの獲得系のみ,大腸菌は生合成系しか持たないのに対し,病原菌の多くは獲得系と生合成系の両方を保有する.つまり,微生物が生存するためだけであれば,どちらか一方の系が存在すればよく,多くの病原菌においてヘムの獲得系と生合成系が共存していることには,病原菌に特有の理由があると考えられる.
1)ヘムによるヘム合成の制御1:ポルフォビリノーゲンデアミナーゼ
ヒトのヘム合成系の場合,ヘム生合成の初期段階であるアミノレブリン酸(ALA)の合成を触媒するアミノレブリン酸合成酵素(ALAS)にヘムが結合することで,ALAの合成が抑制される10).しかし,コレラ菌などのグラム陰性菌の多くはALAの合成過程が異なり,ALASが存在しない.そのため,ヘムが過剰に存在する際に,ヒトなどの哺乳類と異なる制御機構が存在すると考えられるが,知られていない.我々はヘム合成の中間体であるポルフィリノーゲン(PBG)から1-ヒドロキシメチルビラン(1-HMB)の合成を触媒するポルフォビリノーゲンデアミナーゼ(PBGD)(図1)にヘムが結合し,酵素活性が抑制されることを発見した11).
PBGDには,ヘムが結合する共通モチーフは存在しないが,1等量のヘムが結合し,その解離定数が0.33 µMであった.ヘムが存在するとPBGD活性は約25%減少する一方,ミカエリス定数はヘムの有無でほとんど差がなかったことから,ヘムは基質結合部位以外に結合し,活性を阻害すると考えられた.
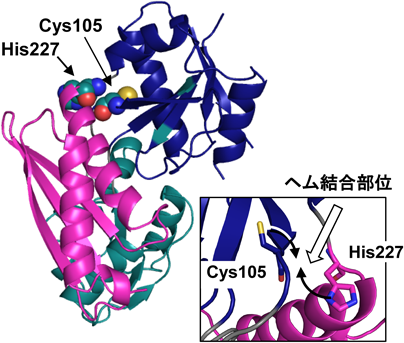
吸収スペクトルと変異体の解析からシステインとヒスチジンがヘムの配位子であると予想された.ヘムと結合していないPBGDの結晶構造によるとCys105の近傍にHis227が存在することから(図3),Cys105とHis227がヘムに配位すると考えた.Cys105とHis227は近くに存在するが,側鎖は異なる方向を向いている.Cys105はドメイン2, His227はドメイン3に存在しているため,ヘムにCys105とHis227が同時に結合するとドメイン2–3間の動きが抑制され,酵素活性が減少すると考えられる.
ヒトのPBGDはコレラ菌由来のPBGDと45%という高いアミノ酸配列の同一性を示すが,ヘムの親和性はコレラ菌由来PBGDの100分の1以下と小さく,ヘムの結合による活性の減少も観測されなかった.ヒトの場合,ALASがヘムによりネガティブフィードバックを受けることから,PBGDがヘムにより,制御される必要がないことと対応していると考えられる.
2)ヘムによるヘム合成の制御2:CyaY
CyaYはヘムの合成酵素であるフェロキラターゼや鉄–硫黄クラスターの生合成系への鉄輸送タンパク質として知られる(図1).我々は,CyaYがヘムと結合し,フェロキラターゼによるヘムの合成を抑制することを見いだした12).ヘムの結合部位はTyr67とCys78で,鉄の結合部位と離れていることから,鉄とヘムが同時に結合することができる.CyaYはヘムと結合すると単量体から三量体に変化し,鉄の親和性は減少した.CyaYを鉄源としてフェロキラターゼの鉄挿入反応を観測すると,鉄イオンを鉄源として利用する場合より,反応速度が20%程度遅くなった.これは,ヘムの結合によりCyaYが三量体を形成し,フェロキラターゼへのヘムの輸送に際し,立体障害が大きくなり,輸送が阻害されたためと考えられる.このことからCyaYは過剰に存在するヘムを感知し.鉄の輸送を介してヘムの合成を抑制する可能性が示された.
ヘムは多くのタンパク質の活性中心に存在し,すべての生物において必須の分子である一方,その高い反応性のため,過剰に存在すると活性酸素などを産生し,有害である.そのため,過剰に存在しないようにその合成量と取り込み量が制御されおり,菌体内のヘム量の制御機構に摂動を与えることで病原菌にダメージを与えることができると考えられる.ヘムの生合成や分解は微生物とヒトで共通している機構が多いことから,ヘムをターゲットとした薬剤はヒトのタンパク質にも作用し,副作用を起こす可能性が高い.しかし,本稿で紹介したように病原菌に特有な性質の分子機構が明らかになりつつあり,そのような性質を利用することにより,これまで存在しない新規の創薬が開発されることが期待される.たとえば,CyaYは鉄の輸送タンパク質であるが,ヘムと結合し,鉄の親和性を変化させ,フェロキラターゼによるヘムの合成を抑制する(図1)12).CyaYのヒトの相同体であるフラタキシンにはヘムが結合しないことから,CyaYを標的とすることで,病原菌のみを対象とした創薬の開発につながるのではないかと考えられる.
引用文献References
1) Wyckoff, E.E., Mey, A.R., & Payne, S.M. (2007) Iron acquisition in Vibrio cholerae. Biometals, 20, 405–416.
2) Wilks, A. & Ikeda-Saito, M. (2014) Heme utilization by pathogenic bacteria: Not all pathways lead to biliverdin. Acc. Chem. Res., 47, 2291–2298.
3) Uchida, T., Sekine, Y., Dojun, N., Lewis-Ballester, A., Ishigami, I., Matsui, T., Yeh, S.-R., & Ishimori, K. (2017) Reaction intermediates in the heme degradation reaction by HutZ from Vibrio cholerae. Dalton Trans., 46, 8104–8109.
4) Uchida, T., Sekine, Y., Matsui, T., Ikeda-Saito, M., & Ishimori, K. (2012) A heme degradation enzyme, HutZ, from Vibrio cholerae. Chem. Commun. (Camb.), 48, 6741–6743.
5) Dojun, N., Sekine, Y., Ishimori, K., & Uchida, T. (2017) Iron chelators inhibit the heme-degradation reaction by HutZ from Vibrio cholerae. Dalton Trans., 46, 5147–5150.
6) Suits, M.D.L., Lang, J., Pal, G.P., Couture, M., & Jia, Z. (2009) Structure and heme binding properties of Escherichia coli O157 : H7 ChuX. Protein Sci., 18, 845–849.
7) Sekine, Y., Tanzawa, T., Tanaka, Y., Ishimori, K., & Uchida, T. (2016) Cytoplasmic heme-binding protein (HutX) from Vibrio cholerae is an intracellular heme transport protein for the heme-degrading enzyme, HutZ. Biochemistry, 55, 884–893.
8) Uchida, T., Funamizu, T., Ogura, M., & Ishimori, K. (2017) Heme iron coordination structure of heme transport protein HutB from Vibrio cholerae. Bull. Chem. Soc. Jpn., 90, 924–930.
9) Arnoux, P., Haser, R., Izadi, N., Lecroisey, A., Delepierre, M., Wandersman, C., & Czjzek, M. (1999) The crystal structure of HasA, a hemophore secreted by Serratia marcescens. Nat. Struct. Biol., 6, 516–520.
10) Scholnick, P.L., Hammaker, L.E., & Marver, H.S. (1972) Soluble δ-aminolevulinic acid synthetase of rat liver. II. Studies related to the mechanism of enzyme action and hemin inhibition. J. Biol. Chem., 247, 4132–4137.
11) Uchida, T., Funamizu, T., Chen, M., Tanaka, Y., & Ishimori, K. (2018) Heme binding to porphobilinogen deaminase from Vibrio cholerae decelerates the formation of 1-hydroxymethylbilane. ACS Chem. Biol., 13, 750–760.
12) Uchida, T., Kobayashi, N., Muneta, S., & Ishimori, K. (2017) The iron chaperone protein CyaY from Vibrio cholerae is a heme-binding protein. Biochemistry, 56, 2425–2434.