脂肪組織には,前駆脂肪細胞という間葉系幹細胞由来の脂肪細胞の幹細胞が存在する.この前駆脂肪細胞の過剰な増殖を抑制し,またその枯渇を防いで一定数を維持する局所の微小環境を「ニッチ」というが,そのメカニズムは明らかではない.一般的にどの組織にもその作用を担う実質細胞の幹細胞が存在し,通常は休眠状態にあるが,組織傷害が起こったときに活性化・増殖し,組織修復に関与する.この組織幹細胞を休眠状態におく環境がニッチである1).個体に対して環境変化が生じた場合,ニッチに存在した組織幹細胞は,増殖や実質細胞に分化することにより環境に適応する.たとえば,脂肪組織においては,高脂肪食によるエネルギー過剰の状態に反応し,前駆脂肪細胞が速やかに増殖・分化することで対応し,その結果,肥満を生ずる.ニッチは,前駆脂肪細胞の数(プール)を規定している.ニッチ数が少なければ,前駆脂肪細胞も少なく,脂肪萎縮性糖尿病のような状態を生じる.これにより脂肪細胞のエネルギー貯蔵・代謝機能を損なうため,高度のインスリン抵抗性を生ずる.一方,ニッチ数が十分にあれば,前駆脂肪細胞も十分に存在する.その例として,Schererらが報告したスーパー肥満マウスがある.このマウスは,ob/obマウス(レプチン遺伝子変異により著しい過食と肥満を起こす)とアディポネクチン過剰発現マウスをかけ合わせて得られたマウスであり,高度の肥満を呈する.皮下脂肪型のスーパー肥満を来すが,耐糖能は保たれる2).ニッチ数が十分であれば,耐糖能異常を来さずにエネルギー過剰状態に対応できる.
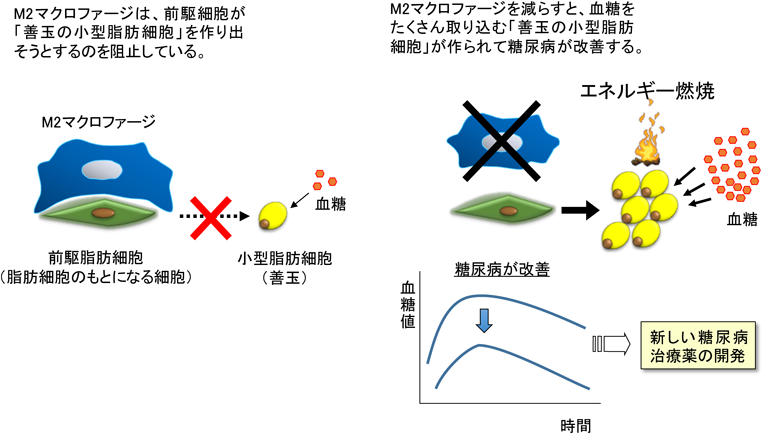
脂肪組織に在住するマクロファージは脂肪細胞の機能を調節する作用を有する.マクロファージはその役割の違いからM1マクロファージとM2マクロファージに分類される.肥満で増加するM1マクロファージからは炎症性サイトカインが分泌され,インスリン抵抗性を誘導する.一方,非肥満時に脂肪組織に存在するM2マクロファージから分泌されるIL-10などのサイトカインは,その抗炎症作用によりインスリン感受性の維持に関与すると考えられてきた3).最近我々は,M2マクロファージが直接,前駆脂肪細胞へ働きかけ,インスリン感受性を制御することを明らかにした.すなわち,M2マクロファージは前駆脂肪細胞のニッチとして,増殖と脂肪細胞への分化を調節し全身の肥満度とインスリン感受性を調節していることを見いだした.本稿では,脂肪組織在住マクロファージによる前駆脂肪細胞の調節を介したインスリン感受性の調節機構について解説する(図1).
M2マクロファージは,組織の恒常性の維持,IL-10を介した抗炎症作用,TGFβを介した線維化の誘導,寒冷時のベージュ細胞の誘導などの役割が報告されてきた3).脂肪組織では非肥満時には,M2マクロファージが大半を占めており,インスリン感受性の維持に関与すると考えられてきたが,その直接のエビデンスはなかった.
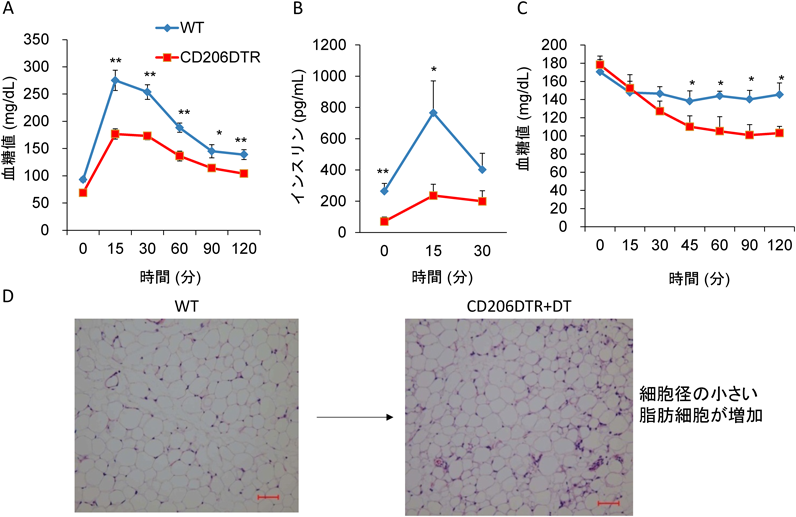
脂肪組織におけるM2マクロファージの役割を明らかにするため,M2マクロファージを任意のタイミングで除去可能なマウスを用いた.M2マクロファージに特異性の高いマーカーであるCd206の下流にジフテリア毒素受容体(diphtheria toxin receptor:DTR)遺伝子を導入したトランスジェニックマウス(CD206 DTRマウス)を作製した.このマウスにジフテリア毒素(diphtheria toxin:DT)を投与すると,脂肪組織中のCD206陽性細胞数を減少させることができる.CD206DTRマウスにDTを投与し,脂肪組織での遺伝子発現をみると,M1マクロファージのマーカーには影響を与えず,M2マクロファージのマーカーが選択的に低下していた.
次に耐糖能について検討した.これまで,M2マクロファージはIL-10などのサイトカイン分泌による抗炎症作用を介して,全身にインスリン感受性を賦与する善玉のマクロファージと報告されている3).M2マクロファージの除去により,インスリン抵抗性が増悪すると予想したが驚くべきことに結果は逆であった.CD206DTRマウスにDTを投与すると,耐糖能は改善し,インスリン投与による血糖の低下が亢進しインスリン感受性を示した.脂肪細胞は小型であり脂肪細胞数自体は増加していた.高脂肪食を負荷した肥満の状態でも同様の実験を行った.肥満状態による脂肪組織での慢性炎症を誘導した状態でも,インスリン抵抗性の改善による耐糖能の改善がみられた.さらに,代謝に好影響を与える遺伝子群や脂肪産生に関連する遺伝子発現が上昇していた.脂肪細胞の大きさについては大型のものが減少し小型脂肪細胞が増加していた.また,組織所見においては,脂肪組織の低酸素領域で炎症性サイトカイン産生の源である王冠様構造(crown-like structure:CLS)も減少した.これらのことから,脂肪組織中のM2マクロファージの抗炎症作用以外のメカニズムにより耐糖能へ好影響を与える機構の存在が示唆された(図2).
このM2マクロファージを除去した後の耐糖能への好影響を与える存在を明らかにするために,DTを投与したCD206DTRマウスの脂肪組織中の細胞の変化を調べた.脂肪組織中の細胞数が増加し,細胞増殖のマーカーであるKi67,ブロモデオキシウリジン(BrdU)に染色される細胞数も増加していた.Cd90, Cd105などの間葉系幹細胞のマーカーの発現上昇とともに,前駆脂肪細胞のマーカーであるPdgfra, Sca1, Cd24, Pref1などの遺伝子発現が上昇していた.フローサイトメトリーでもPDGFRαとSca1共陽性の細胞が増加していた.これらから,増殖している細胞は前駆脂肪細胞の可能性が高いと考えられた.このことを検証するため,マウスに5-エチニル-2′-デオキシウリジン(EdU)を投与し,増殖している細胞の運命を追跡した.EdU投与96時間後において,EdU陽性細胞はPerilipin陽性の成熟した脂肪細胞に分化していた.このことから増殖している細胞は,前駆脂肪細胞であることが明らかになった.
M2マクロファージが前駆脂肪細胞の増殖と分化を抑制していると考えられ,そのメカニズムを明らかにしたいと考えた.M2マクロファージはIL-10のみならずTGFβも高発現している.TGFβは,前駆脂肪細胞のみならず,メラノサイトや血液幹細胞の増殖や分化を抑制している4).そこで,M2マクロファージが前駆脂肪細胞の増殖分化を抑制するメカニズムにTGFβシグナルが関与しているかを検討した.
脂肪組織を間質血管画分(stromal vascular fraction:SVF)から,前駆脂肪細胞,マクロファージに分画し,Tgfb1とその受容体の発現を検討した.M2マクロファージには,Cd206とともにTgfb1が高発現していた.一方,前駆脂肪細胞にはTgfβr1, Tgfβr2ともに高発現していた.さらに共焦点顕微鏡による解析で,CD206陽性細胞にTGFβ1が同時に発現していることを確認した.M2マクロファージを除去すると,TGFβシグナルの下流に位置する遺伝子であるp15, p16, p27, p57の発現が低下した.免疫染色では,M2マクロファージの除去により,TGFβ1, p-SMAD2/3, p27のタンパク質発現が低下した.M2マクロファージ特異的にTgfb1を欠損させるため,CD206-CreERT2マウスとTgfβ1 flox/floxマウスをかけ合わせ,M2マクロファージ特異的TGFβ1欠損マウスを作製した.タモキシフェン投与により,M2マクロファージ特異的にTgfb1を欠損させると,脂肪細胞は小型化し,PDGFRα/Sca1共陽性の前駆脂肪細胞が増加した.これらより,CD206陽性M2マクロファージは,TGFβシグナルを介して前駆脂肪細胞へ働きかけている可能性が示唆された.
TGFβシグナルの影響をより直接的に評価するため,in vitroにおいてもM2マクロファージによる前駆脂肪細胞の増殖と分化の抑制が起きるかを検討した.骨髄単球由来マクロファージ(BMDM)をIL-4とPGE2で処理し,CD206陽性TGFβ1陽性のM2様マクロファージを作製した.これと鼠径部脂肪組織由来の前駆脂肪細胞(adipose tissue-derived stem cell:ASC)を共培養した.するとASCから成熟脂肪細胞への分化が抑制された.また,ASCとM2様マクロファージの共培養中に,TGFβ受容体I/II阻害薬(LY2109761)を添加すると,ASCの増殖と脂肪細胞への分化が回復した.この共培養系に,抗TGFβ1, 2, 3抗体を加えることでも,増殖マーカーの亢進や細胞数の増加を認めた.これらのことからCD206/TGFβシグナルが前駆脂肪細胞の増殖やそれに続く脂肪細胞への分化を調節していることが明らかになった.
脂肪組織のM2マクロファージは,TGFβシグナルを介して前駆脂肪細胞の増殖と分化を抑制し,体脂肪量やインスリン感受性を調節している.幹細胞は,組織維持のために自己複製と分化のバランスをとりながら細胞分裂を行う.細胞分裂を繰り返すとゲノムの複製時に生じるDNA損傷が蓄積し細胞死を招き,幹細胞数が減少する1).M2マクロファージは前駆脂肪細胞の不要な細胞分裂を防ぐことで幹細胞機能の劣化を抑制している.つまり,M2マクロファージが,前駆脂肪細胞のニッチを形成していると考えられる5).
脂肪組織には,内臓脂肪や皮下脂肪といったエネルギーを貯蔵する白色脂肪細胞の他に,熱産生により体温を保つベージュ脂肪細胞が存在する.ベージュ脂肪細胞は,常温では白色脂肪細胞であるが,寒冷曝露により褐色化し熱産生を行う機能を持つようになる6).寒冷な状態ではエネルギーを燃やして体温を維持するために,ベージュ脂肪細胞が増加する.脂肪細胞のベージュ化が促進されると基礎代謝が高まり,抗肥満作用,2型糖尿病の予防効果が期待される.
このことからM2マクロファージを除去すると,ベージュ前駆脂肪細胞の増殖が亢進される可能性も考えられる.
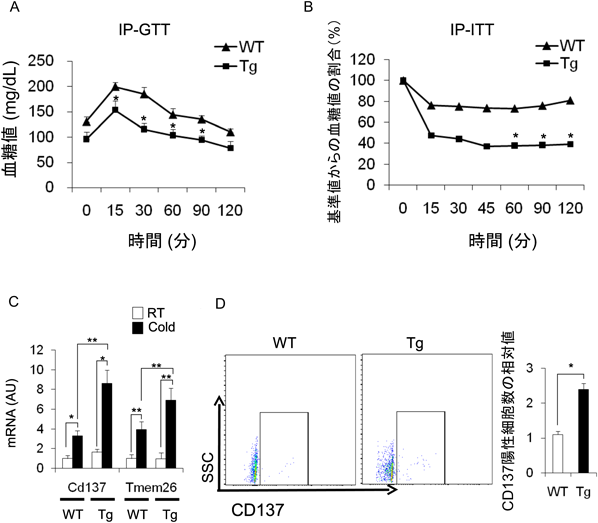
そこで,寒冷刺激によって白色脂肪細胞がベージュ化する際のM2マクロファージの影響を検討した(図3).CD206DTRマウスにDTを投与しM2マクロファージを除去した.対照を野生型として両群マウスに寒冷刺激を加えた.M2マクロファージを除去すると,Ucp1, Cpt-1bなどの褐色化の指標である遺伝子の発現が上昇した.免疫染色ではUCP1陽性細胞がより増加しており,褐変が促進されていた.耐糖能およびインスリン感受性が亢進しており,M2マクロファージの減少で褐変を促進し基礎代謝が高まったためと考えられる.
さらに,M2マクロファージのベージュ前駆脂肪細胞の増殖への影響を検討した.M2マクロファージを除去し寒冷刺激を与えると,脂肪組織においてKi67陽性細胞数が増加し,これに一致して細胞周期関連遺伝子の発現が亢進していた.Cd137やTmem26といったベージュ前駆脂肪細胞のマーカーが上昇していた.フローサイトメトリーではCD137陽性細胞が増加していた.また,蛍光免疫染色で,CD137かつUCP1共陽性細胞が増加し,CD137かつKi67細胞も増加していることから,ベージュ前駆脂肪細胞が増加していることが明らかになった7).
脂肪組織在住M2マクロファージを選択的に除去することで,寒冷時に適応するために活性化するベージュ前駆脂肪細胞が増殖し,基礎代謝が高まって血糖値が改善した.M2マクロファージが,脂肪組織のベージュ化を調節しているという新規の役割が明らかになった.内臓脂肪や皮下脂肪と異なり,基礎代謝を高めるため肥満予防作用があるベージュ脂肪細胞を増やす新規の治療法につながることが期待される.