1899年にアスピリンはドイツのバイエル社から鎮痛剤として発売された.今年で発売以来120年になり,世界で最も知られた歴史を持つ医薬品である.アスピリンの歴史は古く,紀元前400年ごろ,医学の父と呼ばれる古代ギリシャのヒポクラテスが,熱や痛みを和らげることを目的として柳の樹皮を使用したという記録がある.その後,この柳の作用に関わる物質がサリチル酸として分離されたが,強い胃腸障害の副作用が問題となった.そこで,効果を維持したまま副作用を軽減させる目的で合成されたのがアスピリン(アセチルサリチル酸)であり,1971年にはアスピリンが効く分子メカニズムが明らかになった.
プロスタグランジン(PG)とトロンボキサン(TX)からなるプロスタノイドは,生理的あるいは病理的な刺激に応じて産生・分泌され,標的細胞に作用するオータコイドである.プロスタノイドの産生は次のように行われる.まず,細胞膜リン脂質からホスホリパーゼA2(phospholipase A2:PLA2)によって切り出されたアラキドン酸が,シクロオキシゲナーゼ(cyclooxygenase:COX)によりPGH2に変換され,さらに各々のプロスタノイドに特異的な合成酵素によって生理的に重要なプロスタノイド(PGD2, PGE2, PGF2α, PGI2, TXA2)が生合成される.アスピリンはCOXに作用しプロスタノイドの生合成を阻害することで,熱,腫れ,痛みを伴う炎症を抑える.このメカニズムを解明したのが,英国の薬理学者John Vane博士だった.1982年,Vane博士はプロスタグランジンの発見とアスピリンの抗炎症作用の分子メカニズムを発見した功績でノーベル医学生理学賞を受賞した.
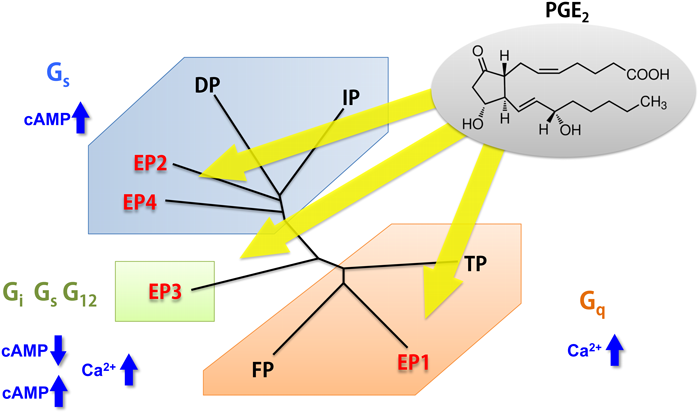
プロスタノイドは,生体内で非常に多彩な作用を示す.これらの作用は,各プロスタノイドに特異的な受容体を介して発揮される.薬理学的研究によりPGD2,PGE2,PGF2α, PGI2,TXA2に特異的な受容体は,各々DP, EP, FP, IP, TPと名づけられ,さらにEPは種々のPGE類似化合物に対する反応性の違いにより4種類のサブタイプ(EP1, EP2, EP3, EP4)に分類された1).また,生化学的研究によりプロスタノイド受容体は,リガンド結合に伴いGタンパク質を活性化することが示唆された.
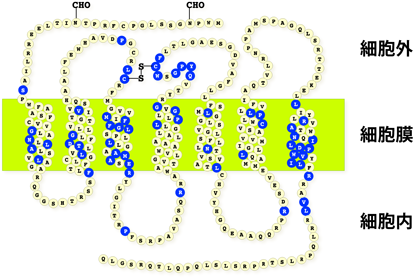
トロンボキサンは血小板の主たるアラキドン酸代謝物であり,血小板の凝集,血管および気管支平滑筋の攣縮という強力な生理活性を有する2).この物質は心筋梗塞など,血栓性疾患や気管支喘息などのアレルギー疾患における重要な脂質メディエーターの一つと考えられている3).しかし,その不安定性のため作用機序の解明は困難であった.1980年代,多くの研究者によりトロンボキサンの安定類縁化合物が多数合成され,これらを用いた薬理学的解析が可能になった.その結果,トロンボキサンが細胞膜上の特異的な受容体を介して作用を発揮することがわかってきた.成宮らは,当時,シオノギ製薬で開発されたトロンボキサン受容体拮抗薬(S-145)をトリチウムラベルし,リガンド結合活性を指標として,ヒトの血小板膜より受容体を可溶化・精製した.さらに,S-145をリガンドアフィニティークロマトグラフィーに利用することで,プロスタノイド受容体としては世界で初めてトロンボキサン受容体を精製することに成功した4).精製タンパク質はSDS-PAGE上,分子量約57,000付近に幅広いバンドとして検出された.これは受容体の糖鎖修飾によるもので,リガンド結合活性を測定していなかったら見落としてしまったであろう.最終精製タンパク質量とリガンド結合活性から,受容体はほぼ単一に精製されていると思われた.成宮らはこの精製した受容体の部分アミノ酸配列を決定し,cDNAクローニングにより,トロンボキサン受容体の一次構造を決定することができ,1991年Nature誌に報告した5).このcDNAは,343アミノ酸からなるタンパク質をコードしており,7回膜貫通構造を持つGタンパク質共役受容体(G protein-coupled receptor:GPCR)に属することが明らかになった(図1).当時,私は修士課程の大学院生で,『実験医学』9月号のミニレビューでそのことを知った.この原著論文を読んだのはその2年後,就職したばかりの会社の図書館だった.プロスタノイド受容体に興味を持った私は,まもなく成宮教授とお会いする機会があり,翌年には会社を辞めて博士課程の大学院生として研究をスタートした.
PGE2は全身の多くの臓器において多彩な生理作用を有している.この多彩な生理作用を解析する目的で数多くのPGE類縁化合物が合成され,組織や細胞での反応性が検討された.その結果,先に述べたように,多彩な生理作用には4種類のサブタイプ受容体(EP1, EP2, EP3, EP4)が関与していることが示された.杉本らは,トロンボキサン受容体のcDNAに基づいてホモロジースクリーニングを行い,1992年にはEP3のcDNAをクローニングすることに成功している6).この受容体は,トロンボキサン受容体と同様に,7回膜貫通構造を持つGPCRであり,哺乳類細胞に強制発現させるとGiタンパク質に共役してcAMPの産生を抑えた.PGE類縁化合物に対する結合活性も,これまで報告されていた活性と一致しており,薬理学的にその存在が示唆されていた受容体(EP3)が初めて実体として捉えられた.その後,立て続けに,その他のPG受容体のクローニングが進み,1995年ごろにはすべてのPG受容体の遺伝子が得られた.
EP3の作用はGiタンパク質を介している場合もあるが,子宮平滑筋の収縮などは細胞内カルシウムの増加を来すことも報告されている7).このように,EP3の場合,情報伝達は単純ではなかった.成宮らは,この点にも目をつけ早くから動いていた.ウシ副腎髄質のクロマフィン細胞では,EP3がcAMP産生を抑制し,同時に細胞内カルシウムを増加する.そこで,彼らはウシ副腎髄質のcDNAクローニングにより,EP3の情報伝達の多様性を解明した8).ウシ副腎髄質には選択的スプライシングに由来する少なくとも4種類のEP3アイソフォーム(EP3A, EP3B, EP3C, EP3D)が存在していた.これらはC末端のみアミノ酸配列が異なり,EP3類縁化合物に対する結合親和性は同じであるが,情報伝達,脱感作などが異なった.EP3AはGiタンパク質に共役してcAMPの産生を抑制し,EP3BとEP3CはGsタンパク質に共役してcAMPの産生を促進した.EP3DはGiタンパク質とGsタンパク質に加え,Gqタンパク質とも共役して細胞内カルシウムを増加させた.EP3のリガンド結合能は単一でまったく区別がつかないにもかかわらず,多彩な情報伝達を仲介することは,アイソフォームの発現の調節によって,PGE2の作用が調節されている可能性を示している.同様に,種を超えてマウス,ヒトなどでも多数のアイソフォームが報告されている9–12).ただし,C末端の配列のみが異なるアイソフォームについては,EP3やTPなどPG受容体の中でも非常に限られている13).
PG受容体全般のアミノ酸配列の相同性は20~30%とあまり高くはない.しかし,種を超えた受容体間の相同性は高い.ヒトとマウスのEP1, EP3, EP4, FP, IP, TPを比較すると,その相同性は各々84, 84, 88, 89, 79, 76%である.プロスタノイドやその類縁体化合物の作用に種差があることはよく知られている.たとえば,PGI2の類縁体であるEP-157はヒトやマウス血小板に対してはアゴニストとして作用するが,ブタやラット血小板に対してはアンタゴニストとして作用する14).ウサギ血小板はトロンボキサンの類縁体であるCTA2やPTA2に対する反応がヒト,ネコ,イヌ血小板のものとは異なっている15).また,TPアンタゴニストであるONO-11120のウサギ血小板に対する効力はヒト血小板に対するものより100倍弱いことが知られている16).このような種差は,受容体間のわずかな構造上の違いに起因していると思われる.事実,ラットTPは,ヒトTPに比べTPアゴニストであるI-BOPに対する結合親和性が10倍高いが,この親和性の差は,第1膜貫通領域の二つのアミノ酸残基によって決定されている17).創薬研究において,動物実験は避けては通れず,動物によって薬効が異なると,最終段階のヒトで効果が得られない場合,あるいは開発の途中で十分な効果が得られない場合,開発を中止することになり,莫大な研究費と労力・時間がむだになってしまう.
プロスタノイド受容体は共役する情報伝達系の種類から三つのグループに分類される.EP1, FP, TPはカルシウムイオン動員系に,EP2, EP4, IP, DPはアデニル酸シクラーゼの活性化に,EP3はアデニル酸シクラーゼの抑制に各々共役する.この分類は,平滑筋に対する機能面からも,それぞれ収縮,弛緩および弛緩の抑制を起こす受容体に対応する.これらのグループ内の受容体間の相同性は,他のグループの受容体との相同性より高く,機能的に同一グループの受容体が近縁関係にあることがわかる.実際,異なるグループのEPサブタイプ間の相同性は同一のグループに属する他のプロスタノイド受容体よりも低い.プロスタノイド受容体は,進化上3種類のクラスターに分類されるが,これは上記の三つのグループに一致している.したがって,EPサブタイプは別々のクラスターに属し,一つの物質を共通のリガンドとする受容体でありながら,構造上はかなり異なり,進化の段階で古い時期から派生していたことになる.これは,プロトタイプのEPからまず機能的に異なるEPサブタイプが進化し,さらに機能的には類似しているがリガンドが異なる受容体に分岐したことを示唆している.
3. プロスタノイド受容体欠損マウスの作出とin vivo解析
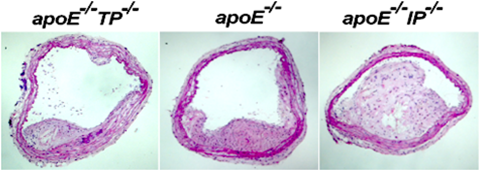
これまで述べたように,分子生物学的な解析手法を用いて,薬理学的に分類されてきたすべてのプロスタノイド受容体の実体が明らかとなった.受容体の構造,機能および生体内分布についても多くの知見が蓄積された.しかし,プロスタノイドの示す多彩な作用がどのプロスタノイドによって発現されているのか,また,プロスタノイドの役割が全体の生理および病態生理において,どの程度の重要性を持つのかは,必ずしも明らかではなかった.そこで,8種類の受容体すべてについて遺伝子欠損マウスが作製された.詳細については,最近の総説などを参考にしてもらいたい18–20).本稿ではその中でも,直接,私が携わった動脈硬化とプロスタノイド受容体についてまとめてみる.1998年から成宮教授の勧めもあり,PGグループのリーダーとしてプロスタノイド受容体欠損マウスのin vivoの解析に携わることになった.所属していた研究室では,プロスタノイド受容体の遺伝子欠損マウスが勢ぞろいし,これらを用いてプロスタノイドの生理・病態生理での役割を解析することが研究の中心となっていた.プロスタノイドの一つ,TXA2は血小板から合成され血小板活性化,血管平滑筋収縮作用を持つ.一方,PGI2は血管内皮より産生され血管平滑筋弛緩作用,血小板凝集抑制作用を持つ.この両者のバランスが,止血や血栓など血管ホメオスタシスの維持に重要であると考えられてきた.しかし,これらプロスタノイドが,慢性の血管病変,特に動脈硬化にどの程度重要な働きをしているかは明らかでなかった.そこで,動脈硬化を自然発症するapoE欠損マウスと,TXA2受容体(TP)欠損マウス,PGI2受容体(IP)欠損マウスの各々をかけ合わせることで,この問題に対する検討を行った(図2).その結果,TP欠損により動脈硬化病変の抑制が,一方,IP欠損により動脈硬化の促進が認められた.これらの解析から,TXA2はTPを介して動脈硬化の促進因子として,一方,PGI2はIPを介して動脈硬化の抑制因子として,生体内で働いていることが明らかとなった21).また,論文掲載から2週間後,Vioxx(COX-2選択的阻害薬)が,血管内皮細胞由来のPGI2の産生を抑えることにより,心筋梗塞や脳梗塞のリスクを上げることが明らかとなり,市場から回収される事件が起こった.我々の論文は,これを遺伝学的に証明した仕事として注目を集めた.プロスタノイドのような生理活性物質の中には,生理・病態生理的条件下において,相反する作用を持つものがある.プロスタノイド受容体欠損マウスは,プロスタノイドの複雑な役割を理解するための重要なツールとなった.
プロスタノイド受容体の一連の遺伝子がクローニングされたことで,プロスタノイド受容体に特異的に保存された部位がいくつか存在していることが明らかとなった.たとえば,第3, 7膜貫通領域と第2細胞外ループは,比較的アミノ酸配列の相同性が高い.これらの領域は種を超えてよく保存されていることから,プロスタノイドの共通構造の認識に関与していると考えられる.また,塩基性アミノ酸であるArg残基が,すべてのプロスタノイド受容体の第7膜貫通領域に保存されている.第7膜貫通領域に塩基性アミノ酸が存在する例としては,ロドプシンのLys残基がレチナールのアルデヒド基とシッフ塩基を形成する例がある22).第7膜貫通領域のArg残基はプロスタノイドに共通したカルボキシ基の結合部位であると考えられている.一方,分子構造からみるとプロスタノイドは,プロスタン酸を基本骨格とし,五員環構造と側鎖の二重結合数によって分類される.すなわち,五員環構造よりD, E, F, Iの4タイプとオキサン環を持つTXに,側鎖の二重結合数より1, 2, 3シリーズに分類される.各プロスタノイドが非常に似通った構造を持つにもかかわらず,各プロスタノイド受容体の相同性が低いことは,まったく対照的である.私は大学で有機化学を学んでいたことも考慮され,1994年から成宮研でプロスタノイドのリガンド結合領域の解析をスタートした.DPとIPのキメラ受容体とアミノ酸変異受容体を用いた解析を行い,五員環構造を認識する部位と側鎖構造を認識する部位が別々に存在することを明らかにした23, 24).五員環構造の認識には,第1~3膜貫通領域が,側鎖構造の認識には第6, 7膜貫通領域がそれぞれ関与していることが明らかとなった.この結果は,マウスのEP3アイソフォーム(EP3α)において,第6膜貫通領域のSer残基が,五員環の15位の水酸基と水素結合しているという報告とよく相関している25).しかし,このようなアプローチには限界があり,2000年以降,GPCRの結晶構造解析により原子分解能レベルで薬物と受容体の結合様式を明らかにしたいと考え始めた.
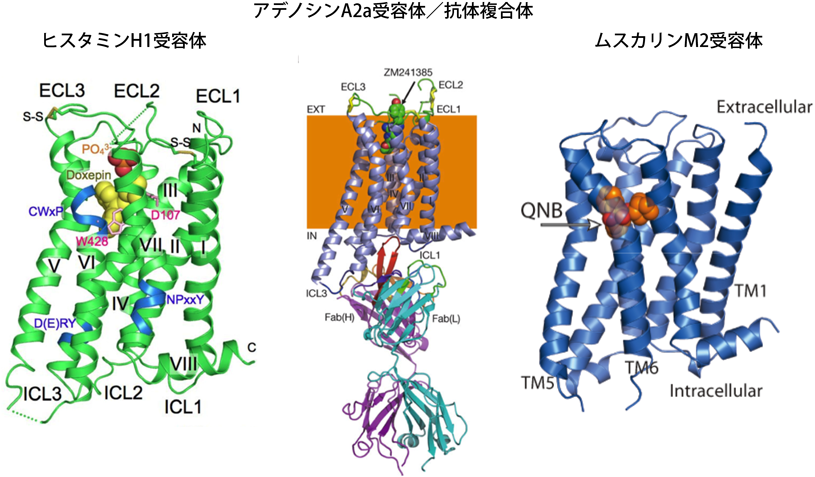
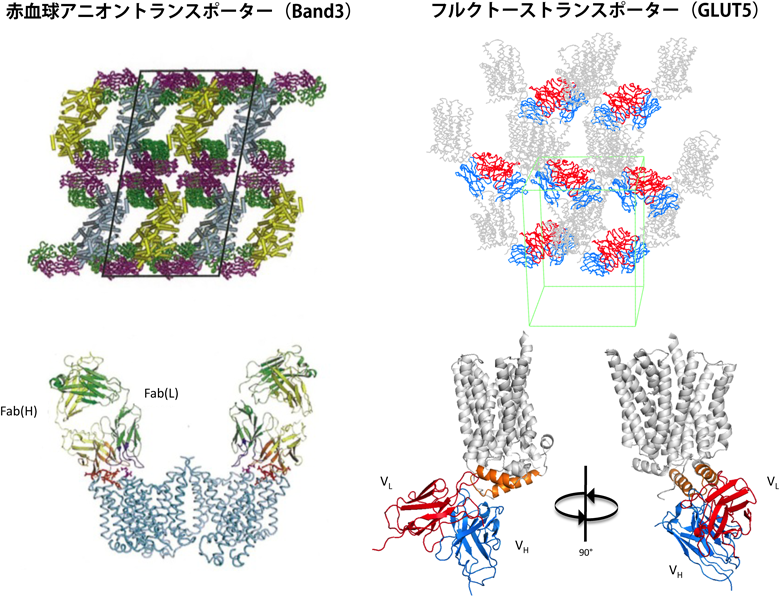
我々はまず,GPCRの高発現株や安定化変異体をハイスループットにスクリーニングするシステムを構築した26).相同組換え活性の高い出芽酵母では,PCRフラグメントを複数準備するだけで,複数の変異を一度に導入することができる.また,GPCRのC末端にGFPを融合させ,蛍光を指標として分子量測定を行うゲル濾過することにより,GPCRの安定性や単分散性を簡便に評価できる.本システムを用いることで,ヒスタミンH1受容体の発現量を約650倍に向上させることができた.安定化したヒスタミンH1受容体とムスカリンM2受容体は,人工的な脂質二重層である脂質キュービック相に結晶化することで,高分解能(3.1 Å, 3.0 Å)で結晶構造を解くことに成功した(図3)27, 28).ヒスタミンH1受容体は安価に大量発現することが可能なメタノール資化酵母を利用し,ムスカリンM2受容体は昆虫細胞を利用して生産した.メタノール資化酵母で生産したヒスタミンH1受容体は,立体構造を決定した世界で初めてのGPCRである.また,アデノシンA2a受容体については,アロステリックなインバースアゴニストとして作用する立体構造認識抗体を作製し,受容体/抗体複合体の結晶構造を解き,アデノシンA2a受容体の不活性化の分子メカニズムを原子分解能レベルで解明した29).現在では,抗体分子は,GPCRだけでなくトランスポーター(Band 3, GLUT5)にも応用できる技術となっている(図4)30, 31).
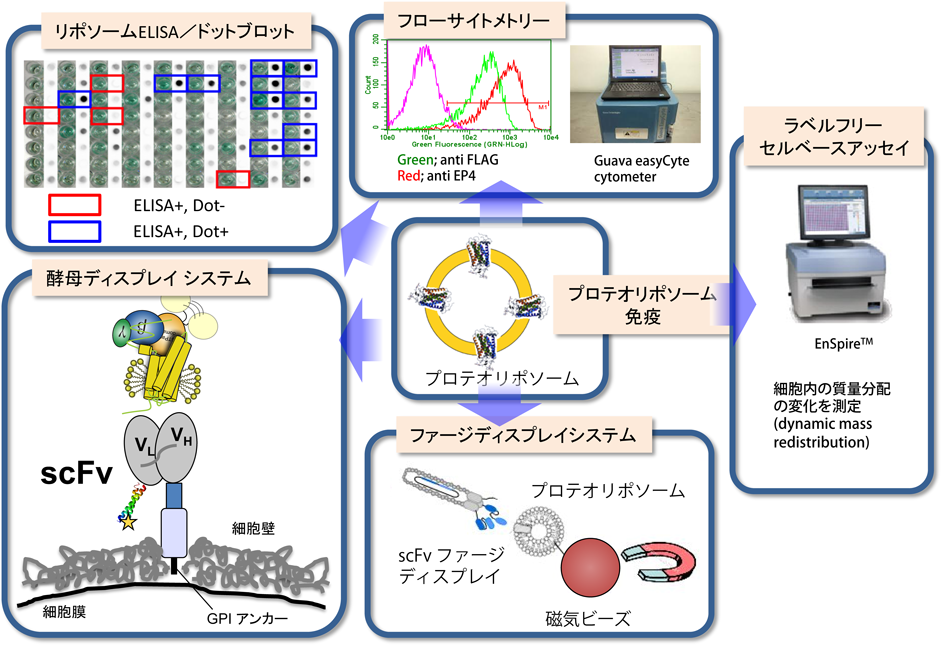
GPCRに対するモノクローナル抗体,特にGPCRの細胞外領域を認識し結合する抗体を用いることで,GPCRの機能解析から構造解析まで行うことが可能になる.GPCRを結晶化するには,界面活性剤のミセル中にGPCRを取り込んだ状態で結晶化する必要がある.この状態ではGPCRの疎水的な表面は特定の構造を持たないミセルで覆われているため,結晶格子の形成に寄与できない.そこで,我々はGPCRの親水性表面の立体構造を特異的に認識して結合し,親水性領域を拡張するとともに,GPCR分子に構造的なゆらぎ(動的構造)が生じないようにある特定のコンホメーションを安定化することによって結晶中の分子パッキングを改善する技術開発に着手した.これまでに,精製標品をプロテオリポソームとして再構築し抗原として利用することで,立体構造を保持した膜タンパク質をマウスに免疫する方法を編み出した.抗体の一次スクリーニングには,リポソームELISA法を利用する.従来のELISA法は,抗原をプラスチックプレートに固相化し,これを認識する抗体を検出する.精製したGPCRを抗原とする場合,界面活性剤存在下での操作となるため,抗原がプラスチックプレートに固相化されないか,あるいは繰り返しの洗浄操作によりGPCRが変性してしまい,立体構造を認識する抗体を得ることができない.そこで,GPCRの安定化および固相化を効率よく行うために,GPCRをビオチン化脂質含有リポソーム中に再構成した上でストレプトアビジンプレートへ固相化し,ELISAを行う方法(リポソームELISA法)を開発した(図5).さらに,立体構造が壊れた変性タンパク質を認識するドットブロット法を組み合わせることにより,ペプチド鎖を認識する抗体とGPCRの立体構造を認識する抗体を区別することが可能となった.GPCR発現細胞とフローサイトメーターを用いることで,GPCRの細胞外領域を認識する抗体が取得できる.EnSpireのラベルフリーセルベースアッセイ機能を組み合わせることで,機能制御抗体のスクリーニングも可能となる.また,京都大学の野村らは,ファージディスプレイ法により,膜タンパク質に対する人工抗体(Fvフラグメント)の作製技術を立ち上げ,親和性の高い抗体分子をスクリーニングするシステムも構築している.
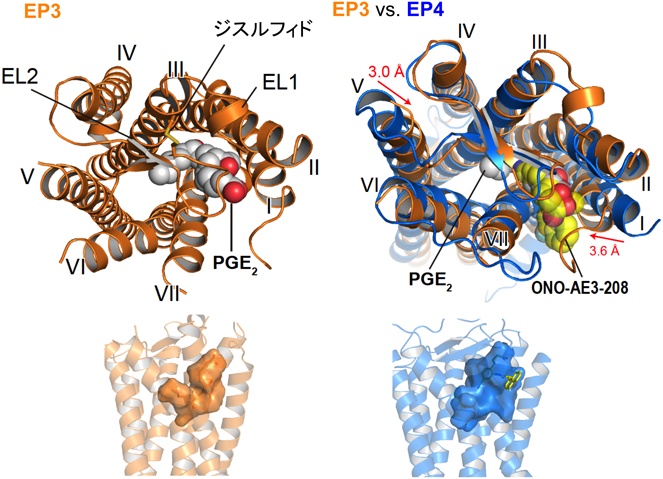
先にも述べたとおり,同じPGE2をアゴニストとするEPサブタイプは,進化の過程で三つのグループに派生し,異なるシグナル伝達系を獲得した(図6).そこで,EPサブタイプの立体構造をX線結晶構造解析により明らかにすることで,シグナル伝達の分子メカニズムを解明し,シグナルを選択的に制御するGPCRの構造生命科学を目指している.私は,抗EP4抗体を取得することに成功した熊本大学の杉本らと共同研究を行ってきた.彼らは,EP4発現細胞をEP4欠損マウスに免疫することにより,PGE2によるcAMP産生を抑えるEP4の細胞外領域を認識する機能性抗体取得に成功している.はじめに,本抗体を熱安定化したEP4に結合させた複合体として,脂質キュービック相での結晶化を試みた.これまで熱安定化したEP4単独では結晶化しなかったが,抗体を結合させることにより,分解能5~6 Åの結晶が得られた.しかし,再現性が悪く,分解能が思うように伸びなかったため,もう一度EP4のコンストラクトから検討することにした.EP4は第3細胞内ループにT4リゾチームを挿入して熱安定化に利用しており,さらに細胞外領域に抗体が結合すると,複合体分子は全体的にサイズが大きくなるため,モノオレインを主成分とした脂質キュービック相での流動性が低下していることが考えられた.そこで,T4リゾチームを第3細胞内ループから外し,第3細胞内ループを短くしたコンストラクトを作製した.このコンストラクトに抗体を結合させた複合体は,単一に精製することができ,単分散性もよかったので結晶化を試みた.いくつかの条件から数日で結晶が現れ,結晶化の再現性も非常によくなった.次に,共同研究者である京都大学の木下と千葉大学の村田が開発した理論的耐熱化予測法を利用して,さらにEP4の熱安定性を向上するアミノ酸残基を探索した.その結果,結晶の分解能を向上させることに成功し,EP4アンタゴニスト(ONO-AE3-208)が結合したEP4と抗体の複合体の立体構造を解明することに成功した32).さらに,ナチュラルアゴニスト(PGE2)が結合したEP3のX線結晶構造解析にも成功することができた33).なお,EP3の立体構造解明には抗体を使用しなかった.
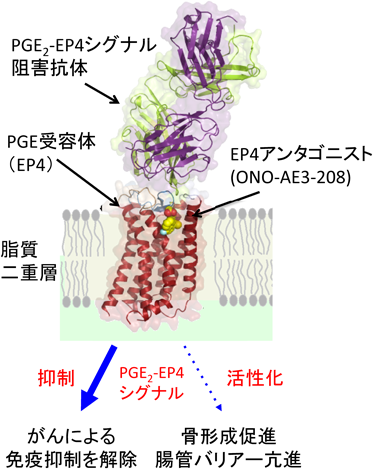
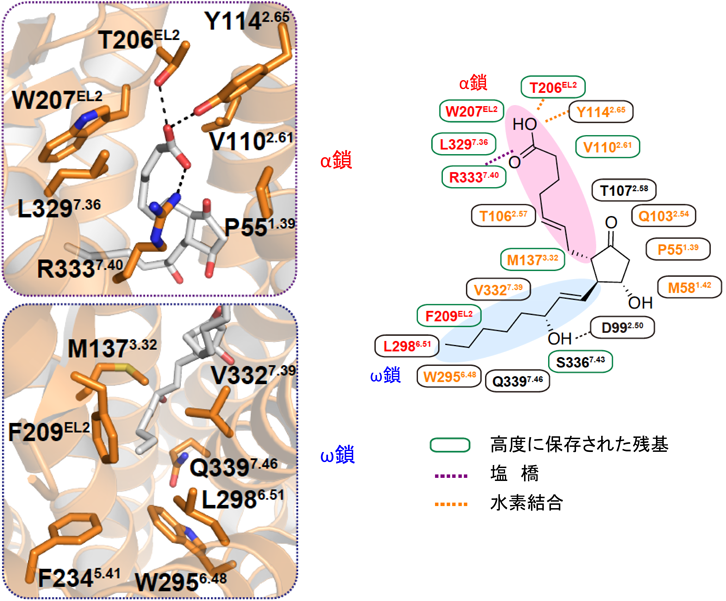
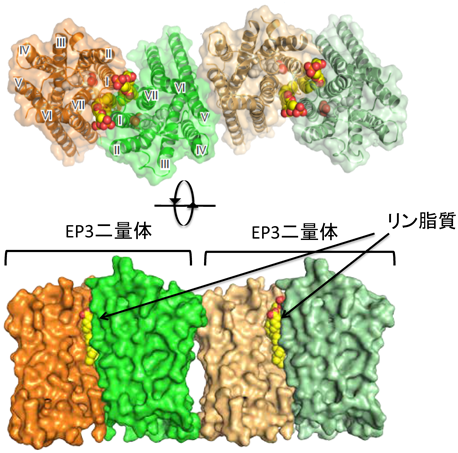
EP3とEP4の全体構造は,これまでに報告されているGPCRと同様に7本の膜を貫通したらせん構造を持っていた.第2細胞外ループがβヘアピン構造をとり,オルソステリック部位に蓋をするように覆っていた(図7).そのため,EP4アンタゴニストは細胞の外側からダイレクトに受容体に結合するのではなく,いったん,細胞膜の中に入り,脂質二重層との境界面に結合することが示唆された.PGE2-EP4シグナルを阻害する抗体は,EP4の細胞外領域(アロステリック部位)に結合し,PGE2の結合を阻害していることがわかった(図8).脂質受容体をターゲットにした抗体医薬の開発につながることを期待している.EP3とEP4の構造を比較すると内因性アゴニストであるPGE2は,EP4アンタゴニストよりもさらに受容体の奥に入り込み,完全に閉じ込められていることがわかる(図7).PGE2のα鎖のカルボキシ基は,第7膜貫通領域のArg残基(R333)と塩橋を形成し,疎水性を保持するω鎖は受容体の奥まで伸び,活性化に重要な第6膜貫通領域のTrp残基(W295)まで達していた(図9).また,EP3の細胞内領域では,第6膜貫通領域が大きく外側に開いており,Gタンパク質が共役するスペースが空いていた.さらに,EP3はホモ二量体を形成しており,細胞膜の成分であるリン脂質がホモ二量体の分子間で形成した二量体を安定化していると思われた(図10).今後,新しいタイプの薬剤設計につながる重要な構造情報として注目している.
アスピリンはプロスタグランジン合成酵素のシクロオキシゲナーゼを阻害することで,すべてのPGの生合成を抑えてしまう.しかし,前述したTXA2とPGI2のように,PGの中には相反する作用を示すものが含まれており,すべてのPGを抑えてしまうことは必ずしも得策ではない.PGは受容体を介してさまざまな作用を呈する.たとえば,アゴニストが結合しEP4の賦活作用が発揮されると骨形成,腸管バリアーが亢進し,アンタゴニストが結合しEP4の機能が阻害されるとがんによる免疫抑制を解除する(図8).近年,免疫チェックポイント阻害薬とEP4アンタゴニストの併用効果が注目されている.ナチュラルアゴニスト(PGE2)とEP3,アンタゴニストとEP4の結合様式が明らかになったことで,活性型および不活性型EP受容体の立体構造を基に,より有効性が高く副作用の少ない治療薬の開発が期待される.
すべてのプロスタノイド受容体の第7膜貫通領域で保存されているArg残基が,選択的なGタンパク質の連関に関与しているという報告がある34).PGE2アナログの中には,Sulprostoneのようにα鎖のカルボキシ基が修飾され電荷を失っていても活性を示す非荷電性アゴニストが存在する.PGE2をはじめ負に荷電したカルボキシ基を持つ荷電性アゴニストは,ウシEP3アイソフォーム(EP3D)を介して,GiおよびGs活性を示す.一方,非荷電性アゴニストはGi活性についてはPGE2と同等の活性を示すが,Gs活性をほとんど示さない.この結果から,Arg残基の正電荷とアゴニストのカルボキシ基の負電荷との相互作用がGタンパク質の種類によって異なるという仮説がたてられた.そこで,これらを裏づけるため,第7膜貫通領域のArg残基をGlu残基に置換し,ここに電荷を持たない受容体を作った.ここの変異受容体は,PGE2結合能を持ち,Gi活性を示した.ところが,荷電性アゴニストでみられたGs活性を示さなくなっていた.これらの実験からEP3においては,Arg残基の正電荷とアゴニストのカルボキシ基の負電荷との相互作用は,Giを活性化するためには必ずしも必要ではなく,Gs活性を示すためには重要であることが示唆された.EP3には選択的スプライシングにより,C末端の配列だけが異なるアイソフォームが存在し,シグナル伝達に違いが認められている35).今回,PGE2の結合する活性型EP3がホモ二量体を形成していた(図10).二量体の界面に第1, 7膜貫通領域やカルボキシル末端が存在することから,EP3のオリゴマー化により共役するGタンパク質の種類が変化する可能性もある.また,一般的にフレキシブルな領域と考えられているアイソフォーム間で配列の異なるC末端が,オリゴマー化やGタンパク質との共役により,新たな立体構造を形成する可能性もある.これまでに20種類以上のGPCRの構造が明らかになったが,すべてが単量体をターゲットにしており,GPCRのオリゴマーをターゲットにした構造解析は皆無である.立体構造を基盤としたオリゴマーGPCRのシグナル伝達機構を明らかにすることで,生理的・病態生理的条件下におけるGPCRの新しい役割解明につながるかもしれない.
2017年のノーベル化学賞は,「クライオ電子顕微鏡法の開発」に授与された.X線結晶構造解析,核磁気共鳴に加え,第三の構造解析法として注目を集めている.生体高分子をわざわざ結晶化する必要がないため,高純度で均一なサンプルを調製する必要がない.必要なサンプル量が格段と少なくなり,極低温・凍結状態で測定することで,生体環境に近い状態の試料から3 Å程度の解像度の構造を得ることができる.アミノ酸や核酸の配列をもとにした原子モデルを組み立てることが可能になっている.また,電子線で一つ一つの分子をみているので,結晶化を必要としない.さらに,大きな複合体の観察が可能となり,必ずしもすべての分子の形が同じである必要がないため,今後,フレキシブルな領域を含み,結晶化がきわめて難しいGPCRやオリゴマー化したGPCRの構造解析に応用されることを期待している.
引用文献References
1) Coleman, R.A., Kennedy, I., Humphrey, P.P.A., Bunce, K., & Lumler, P.(1990) in Comprehensuve Medicinal Chemistry (Emmett, J.C. eds), Vol. 3, pp. 643–714, Pergamon Press, Oxford, UK.
2) Samuelsson, B., Goldyne, M., Granström, E., Hamberg, M., Hammarström, S., & Malmsten, C. (1978) Prostaglandins and thromboxanes. Annu. Rev. Biochem., 47, 997–1029.
3) Ogletree, M.L. (1987) Overview of physiological and pathophysiological effects of thromboxanes A2. Fed. Proc., 45, 133–138.
4) Ushikubi, F., Nakajima, M., Hirata, M., Okuma, M., Fujiwara, M., & Naurmiya, S. (1989) Purification of the thromboxane A2/prostaglandin H2 receptor from human blood platelets. J. Biol. Chem., 264, 16496–16501.
5) Hirata, M., Hayashi, Y., Ushikubi, F., Yokota, Y., Kageyama, R., Nakanishi, S., & Narumiya, S. (1991) Cloning and expression of cDNA for a human thromboxane A2 receptor. Nature, 349, 617–620.
6) Sugimoto, Y., Namba, T., Honda, A., Hayashi, Y., Negishi, M., Ichikawa, A., & Narumiya, S. (1992) Cloning and expression of a cDNA for mouse prostaglandin E receptor EP3 subtype. J. Biol. Chem., 267, 6463–6466.
7) Goureau, O., Tanfin, Z., Marc, S., & Harbon, S. (1992) Diverse prostaglandin receptors activate distinct signal transduction pathways in rat myometrium. Am. J. Physiol., 263, C257–C265.
8) Namba, T., Sugimoto, Y., Negishi, M., Irie, A., Ushikubi, F., Kakizuka, A., Ito, S., Ichikawa, A., & Narumiya, S. (1993) Alternative splicing of C-terminal tail of prostaglandin E receptor subtype EP3 determines G-protein specificity. Nature, 365, 166–170.
9) Sugimoto, Y., Negishi, M., Hayashi, Y., Namba, T., Honda, A., Watabe, A., Hirata, M., Narumiya, S., & Ichikawa, A. (1993) Two isoforms of the EP3 receptor with different carboxyl-terminal domains. Identical ligand binding properties and different coupling properties with Gi proteins. J. Biol. Chem., 268, 2712–2718.
10) Negishi, M., Sugimoto, Y., Irie, A., Narumiya, S., & Ichikawa, A. (1993) Two isoforms of prostaglandin E receptor EP3 subtype. Different COOH-terminal domains determine sensitivity to agonist-induced desensitization. J. Biol. Chem., 268, 9517–9521.
11) Irie, A., Sugimoto, Y., Namba, T., Harazono, A., Honda, A., Watabe, A., Negishi, M., Narumiya, S., & Ichikawa, A. (1993) Third isoform of the prostaglandin-E-receptor EP3 subtype with different C-terminal tail coupling to both stimulation and inhibition of adenylate cyclase. Eur. J. Biochem., 217, 313–318.
12) Adam, M., Biuem, Y., Rushmore, T.H., Müller, G., Bastien, L., McKee, K.T., Metters, K.M., & Abramovitz, M. (1994) Cloning and expression of three isoforms of the human EP3 prostanoid receptor. FEBS Lett., 338, 170–174.
13) Hirata, T., Ushikubi, F., Kakizuka, A., Okuma, M., & Narumiya, S. (1996) Two thromboxane A2 receptor isoforms in human platelets. Opposite coupling to adenylyl cyclase with different sensitivity to Arg60 to Leu mutation. J. Clin. Invest., 97, 949–956.
14) Armstrong, R.A., Lawrence, R.A., Jones, R.J., Wilson, N.H., & Collier, A. (1989) Functional and ligand binding studies suggest heterogeneity of platelet prostacyclin receptors. Br. J. Pharmacol., 97, 657–668.
15) Burke, S.E., Lefer, A.M., Nicolaou, K.C., Smith, G.M., & Smith, J.B. (1983) Responsiveness of platelets and coronary arteries from different species to synthetic thromboxane and prostaglandin endoperoxide analogues. Br. J. Pharmacol., 78, 287–292.
16) Narumiya, S., Okuma, M., & Ushikubi, F. (1986) Binding of a radioiodinated 13-azapinane thromboxane antagonist to platelets: correlation with antiaggregatory activity in different species. Br. J. Pharmacol., 88, 323–331.
17) Dorn, G.W. 2nd, Davis, M.G., & D’Angelo, D.D. (1997) Structural determinants for agonist binding affinity to thromboxane/prostaglandin endoperoxide (TP) receptors. Analysis of chimeric rat/human TP receptors. J. Biol. Chem., 272, 12399–12405.
18) 杉本幸彦(2015)プロスタグランジンによる神経機能の調節機構.日本薬理学雑誌,145, 237–242.
19) 杉本幸彦(2017)雌性生殖生理におけるプロスタグランジン受容体の役割.別冊BIO Clinica, 6, 16–21.
20) 稲住知明,杉本幸彦(2018)プロスタグランジンE2による炎症,免疫調節機構.臨床免疫・アレルギー科,70, 216–220.
21) Kobayashi, T., Tahara, Y., Matsumoto, M., Iguchi, M., Sano, H., Murayama, T., Arai, H., Oida, H., Yurugi-Kobayashi, T., Yamashita, J.K., et al. (2004) Roles of thromboxane A2 and prostacyclin in the development of atherosclerosis in apoE-deficient mice. J. Clin. Invest., 114, 784–794.
22) Findlay, J.B. & Pappin, D.J. (1986) The opsin family of proteins. Biochem. J., 238, 625–642.
23) Kobayashi, T., Kiriyama, M., Hirata, M., Hirata, T., Ushikubi, F., & Narumiya, S. (1997) Identification of domains conferring ligand binding specificity to the prostanoid receptor. Studies on chimeric prostacyclin/prostaglandin D receptors. J. Biol. Chem., 272, 15154–15160.
24) Kobayashi, T., Ushikubi, F., & Narumiya, S. (2000) Amino acid residues conferring ligand binding properties of prostaglandin I and prostaglandin D receptors. Identification by site-directed mutagenesis. J. Biol. Chem., 275, 24294–24303.
25) Huang, C. & Tai, H.H. (1996) Ser-268 plays an important role in ligand binding of prostaglandin E2 receptor EP3alpha subtype. Arch. Biochem. Biophys., 327, 161–166.
26) Shiroishi, M., Tsujimoto, H., Makyio, H., Asada, H., Yurugi-Kobayashi, T., Murata, T., Nomura, N., Haga, T., Iwata, S., & Kobayashi, T. (2012) Platform for the rapid construction and evaluation of GPCRs for crystallography in Saccharomyces cerevisiae. Microb. Cell Fact., 11, 78.
27) Shimamura, T., Shiroishi, M., Weyand, S., Tsujimoto, H., Winter, G., Katritch, V., Abagyan, R., Cherezov, V., Liu, W., Han, G.W., et al. (2011) Structure of the human histamine H1 receptor complex with doxepin. Nature, 475, 65–70.
28) Haga, K., Kruse, A.C., Asada, H., Yurugi-Kobayashi, T., Shiroishi, M., Zhang, C., Weis, W.I., Okada, T., Kobilka, B.K., Haga, T., et al. (2012) Structure of the human M2 muscarinic acetylcholine receptor bound to an antagonist. Nature, 482, 547–551.
29) Hino, T., Arakawa, T., Iwanari, H., Yurugi-Kobayahi, T., Ikeda-Suno, C., Nakada-Nakura, Y., Kusano-Arai, O., Weyand, S., Shimamura, T., Nomura, N., et al. (2012) G-protein-coupled receptor inactivation by an allosteric inverse-agonist antibody. Nature, 482, 237–240.
30) Arakawa, T., Kobayashi-Yurugi, T., Alguel, Y., Iwanari, H., Hatae, H., Iwata, M., Abe, Y., Hino, T., Ikeda-Suno, C., Kuma, H., et al. (2015) Crystal structure of the anion exchanger domain of human erythrocyte band 3. Science, 350, 680–684.
31) Nomura, N., Verdon, G., Kang, H.J., Shimamura, T., Nomura, Y., Sonoda, Y., Hussien, S.A., Qureshi, A.A., Coincon, M., Sato, Y., et al. (2015) Structure and mechanism of the mammalian fructose transporter GLUT5. Nature, 526, 397–401.
32) Toyoda, Y., Morimoto, K., Suno, R., Horita, S., Yamashita, K., Hirata, K., Sekiguchi, Y., Yasuda, S., Shiroishi, M., Shimizu, T., et al. (2019) Ligand binding to human prostaglandin E receptor EP4 at the lipid-bilayer interface. Nat. Chem. Biol., 15, 18–26.
33) Morimoto, K., Suno, R., Hotta, Y., Yamashita, K., Hirata, K., Yamamoto, M., Narumiya, S., Iwata, S., & Kobayashi, T. (2019) Crystal structure of the endogenous agonist-bound prostanoid receptor EP3. Nat. Chem. Biol., 15, 8–10.
34) Negishi, M., Irie, A., Sugimoto, Y., Namba, T., & Ichikawa, A. (1995) Selective coupling of prostaglandin E receptor EP3D to Gi and Gs through interaction of alpha-carboxylic acid of agonist and arginine residue of seventh transmembrane domain. J. Biol. Chem., 270, 16122–16127.
35) Hasegawa, H., Negishi, M., & Ichikawa, A. (1996) Two isoforms of the prostaglandin E receptor EP3 subtype different in agonist-independent constitutive activity. J. Biol. Chem., 271, 1857–1860.
著者紹介Author Profile
 清水(小林) 拓也(こばやし-しみず たくや)
清水(小林) 拓也(こばやし-しみず たくや)関西医科大学医学部医化学講座教授.博士(医学).
略歴1967年七尾で生まれ金沢で育つ.91年金沢大学薬学部卒業.93年同大学院薬学研究科修士課程を修了.コスモ石油(株)で1年間勤務し,94年から京都大学医学部成宮周教授の下,プロスタノイド受容体の研究をスタート.2018年より現職.
研究テーマと抱負「くすり」を作りたいと思い,薬学部に進学し有機合成化学を学ぶ.くすりが作用する生体分子についてもっと知りたくなり,医学部で薬理学の研究をスタート.くすりのターゲットのしくみを知るために構造生物学の技術を導入するも,まだまだ遠き道のりかな.
ウェブサイトhttp://www.kmu.ac.jp/faculty/medical/category2/unit5.html
趣味家族と一緒に過ごすこと.