動物細胞のある驚異的な振る舞いが,古くから人々の関心を惹きつけてきた.動物の組織を機械的に潰したり,2価陽イオンを除いたり,タンパク質分解酵素で処理したりすると,細胞が生きたまま解離されるが,この細胞を適切な方法で培養すると,細胞が再び集まり,さらには,異なるタイプの細胞が選別されて,元の組織に自然に戻る.1900年代初頭から半ばにかけて発見された現象で,無脊椎動物の初期胚を使った実験では,体全体が回復することすらある.このような観察から,「細胞は自律的に組織を構築できる」という概念が生まれ,これを可能にする細胞の性質,能力について広範な研究が始まった.細胞による「自己組織化」能力は,現代再生医学を支える生物学的基盤の一つでもある.
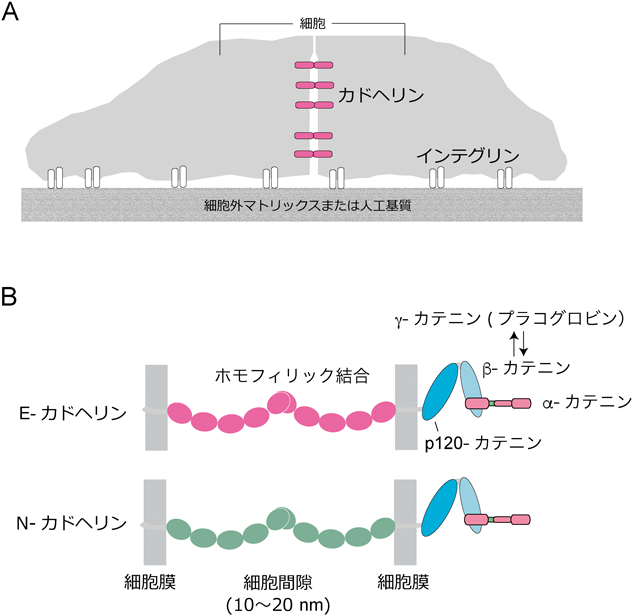
自己組織化研究における当初の関心事は,そもそも細胞はどのようにくっつくのか,そして,どのように選別されるのかであった.細胞選別は多種類の細胞からなる組織形成の基本過程なので,人々の関心を惹き,さまざまなモデルが提案されたが,細胞接着機構自体が不明のままでは決着がつくはずもなかった.混沌の時代が長く続いたが,1980年代に入り二つの重要な細胞接着分子が同定された.インテグリンとカドヘリンである.ともに膜貫通型のタンパク質で,インテグリンは細胞と細胞外マトリックス(ラミニン,コラーゲン,フィブロネクチンなどが主な構成成分)の接着(細胞-基質接着)を担い,カドヘリンは細胞どうしの接着(細胞間接着)のために働く(図1A).それ以降,他の接着分子(クローディン,ネクチンを含む多様なIgスーパーファミリー分子群など)が次々と発見され,細胞接着はタンパク質の働きの問題として捉えることができるようになった.
そして,30~40年の時が流れ,組織構築の基盤となる細胞接着の仕組みはずいぶんわかったと思う.しかし研究というもの,深まれば深まるほど,新しい疑問が次々と生まれ,なかなかゴールには至らない.また,簡単に解けると思った問題が意外に解けていない.私の研究グループはカドヘリンを見つけ,その働きを調べてきたが,いまだに悩ましい日々が続く.未解決問題の一つは,細胞間接着の細胞質側からの制御の仕組みである.カドヘリンにはβ-カテニンが結合し,これにα-カテニンが結合して,カドヘリン—カテニン複合体を作る(図1B).これが核となって,さまざまな分子が集結し,複雑な細胞間接着制御装置が形成される.細胞間の接着は,発生,形態形成,がん転移などの過程において動的に変化し,この変化が細胞行動を左右するので,この装置の働き方の解明が重要であると考え,研究を続けてきた.そろそろ店じまいが近づいているところ,総説執筆の機会をいただいたので,細胞間接着制御装置について,我々がこの問題にどう取り組み,何が未解決なのかについてまとめておくこととした.この研究分野に関心がある方々の参考になれば幸いである.科学は人間活動であることから,エピソードを交えて書かせていただくことし,この分野の全体像を客観的に俯瞰する場合には,筆者が執筆した別の総説を参照いただきたい1).
カドヘリンは膜貫通型タンパク質で,その細胞外領域が細胞表面に突き出ており,同じカドヘリン分子どうしが結合することにより(ホモフィリック結合),向かい合う細胞膜を接着させる(図1B).カドヘリンの機能を説明するとき,これで十分なはずだが問題はそう単純ではなかった.細胞質領域が必要なのだ.その発見の物語から始めたい.
私の研究室では,カドヘリンの発見当時,まず,3種類のカドヘリン・サブタイプ(E, N, P-カドヘリン)を同定していた.細胞外領域のアミノ酸配列にはサブタイプ間で変異があり,同じサブタイプを発現する細胞どうしが選択的に集合することから,細胞選別への関与が確定した2).ところが,細胞質領域はサブタイプ間でよく保存されており,サブタイプ間に共通する重要機能の存在を暗示している.しかし,細胞質領域の問題にアプローチするよい方法がなく放置していた.1987年,大学院生の永渕昭良はE-カドヘリン(主として上皮細胞で発現されるカドヘリン)のcDNAを分離し,これをL細胞(カドヘリンを発現せず相互の接着が弱い)に導入すると細胞接着が誘導されるという,細胞接着分野におけるエポックメイキングな成果をあげた3).cDNAを手中にしたことから,細胞質領域の機能解析も可能となり,永渕はその研究に着手することになる.そして,E-カドヘリンをL細胞に導入するとき,細胞質領域を除去しておくと接着誘導活性がなくなることを発見した4).カドヘリン細胞質領域の重要性を指摘した最初の論文である.細胞質領域が細胞骨格系との相互作用に働くらしいこともこの時示唆している.次のステップでは,細胞質領域には何らかの分子が結合してカドヘリンの機能を支えるのではないかと推測し,実際,二つのタンパク質が細胞質領域に結合,そのうちの一つがC末端側に結合することを見いだした5).そして,このC末端側こそが,カドヘリンの接着機能にとって重要であることも示している.
パスツール研究所のF. Jacob博士の研究室は,マウス初期胚の研究から,E-カドヘリンと同一の分子ウボモルリン(uvomorulin)を発見しており,我々にとって競争相手だった.上記の永渕によるカドヘリン結合タンパク質の発見と時を同じくして,Jacob研究室を経たR. Kemler博士の研究室から,E-カドヘリン/ウボモルリンには3種類の細胞質タンパク質が結合するという論文が出版され,α-,β-,γ-カテニンと命名された6).次のKemler研からの論文では,β-カテニンがE-カドヘリン細胞質領域C末端側に結合すること,α-カテニンはF-アクチンに結合することなど,カドヘリン—カテニン複合体についての基本的性質が確認されていく7).これに先んじて,別の研究室から,細胞間結合に分布する「プラコグロビン」が発見されており8),後に,γ-カテニンとプラコグロビンは同一分子であることがわかる.また,γ-カテニンとβ-カテニンは,E-カドヘリン細胞質領域C末端に,競合して結合することもわかっていく(図1B).こうして,細胞質領域問題は慌ただしく進展し,我々の研究グループはこの分野に先鞭をつけながらも,「カテニン」命名のチャンスを逸する.ちなみに,上記のKemler研論文の筆頭著者は,小沢政之博士で,現在,私の研究室のメンバーとしてお世話になっている.
そして,1991年,β-カテニンがクローニングされ,γ-カテニン/プラコグロビンと同類の分子であることが確定9)した.前後して,ショウジョウバエのセグメントポラリティ遺伝子armadilloがクローニングされ10),この遺伝子産物が,なんと,γ-カテニン/プラコグロビン類似分子であることがわかる(後に,アルマジロ・リピートタンパク質としてまとめられる).突然,接着関連タンパク質と発生情報因子が関連づけられたのである.少し遅れて,1993年,β-カテニンが大腸がん抑制遺伝子産物APC(adenomatous polyposis coli)に結合することが報告される11, 12).それ以来,β-カテニン/Armadilloが転写因子Tcf/Lefと結合し,Wnt/Winglessシグナル系による転写制御のために作用することがわかり大分野へと発展する.刻々と新事実が発見される,実にわくわくする時代だった.
β-カテニンが,細胞接着と遺伝子転写制御というまるで異なる分子現象に関わるという発見には,正直なところ当惑した.何かを見誤っているのではないかという気持が永らく続く.個々のタンパク質の役割は単純明快に違いない,という勝手な思い込みのためだが,どうやら,すっきりしないことの方が生命現象の本質のようだ(β-カテニン,今では,多機能タンパク質とさりげなく呼ばれる).ところで,β-カテニンの転写制御活性に注目する研究が増えるに従い,困ったことに,β-カテニンが接着にも関与していることを無視する傾向が強まった.とりわけ,β-カテニンを除去したときに生じる変化については,両機能を考慮の上,議論しなければならないはずなのに,転写活性機能だけで結論してしまう論文が多数となった.現実には,両機能を分離して解析することは難しい.接着に関しては,β-カテニンの欠失をγ-カテニン/プラコグロビンが補うこともあり複雑である13).ただ,一つの分野が強くなると,他を無視しても通ってしまうという風潮を実感した.しかし,この分野も最盛期を終えて今や冷静となり,β-カテニンの活動を論じるとき,接着問題も視野に入れることが多くなった.実際,最近では,細胞間接着系自体が細胞内シグナルを動かして転写制御にまで至ることが明らかにされつつあり14, 15),今後,β-カテニンの機能的二面性に関しても,なぜ,そういうことになっているのか,理解が深まるかもしれない.
β-カテニンとその関連分子の同定がにぎやかな中,月田承一郎博士(故人)の研究室(当時,東京都臨床研/岡崎生理研)に移籍した永渕は,α-カテニンのクローニングを着々と進め,これがビンキュリンに類似のタンパク質であることを発見する13).私の方では,α-カテニンにも複数タイプがあるのではないかと想像し,大学院生の平野伸二が,N-カドヘリン(神経系などで発現)と共沈するカテニンの特性を調べてみると,永渕が見つけたα-カテニンとは少し異なることに気づいた.これをαN-カテニンと命名し,先に同定されたα-カテニンをαE-カテニンと再命名した16).発表にあたり,単に新しいα-カテニンが見つかったというだけではインパクトに欠けるので,機能の検証をしたいところだった.
当時,がん研究の分野では,カドヘリンと,がんの悪性化との関係に興味が持たれ始めていた.悪性化したがんでは組織型が乱れ,細胞の分散が起きるからである.私自身も,がん研究者と共同研究を行っていたが,その1人,国立がんセンター研究所の廣橋説雄博士(故人)のグループは,がん由来細胞株のカドヘリン,カテニンの発現を調べる中,肺がん由来のPC9細胞がα-カテニンを発現していないことに気づいていた17).興味深いことに,PC9細胞はE-カドヘリンとβ-カテニンを正常に発現しているにも関わらず,細胞が緩やかにしか集まらない.
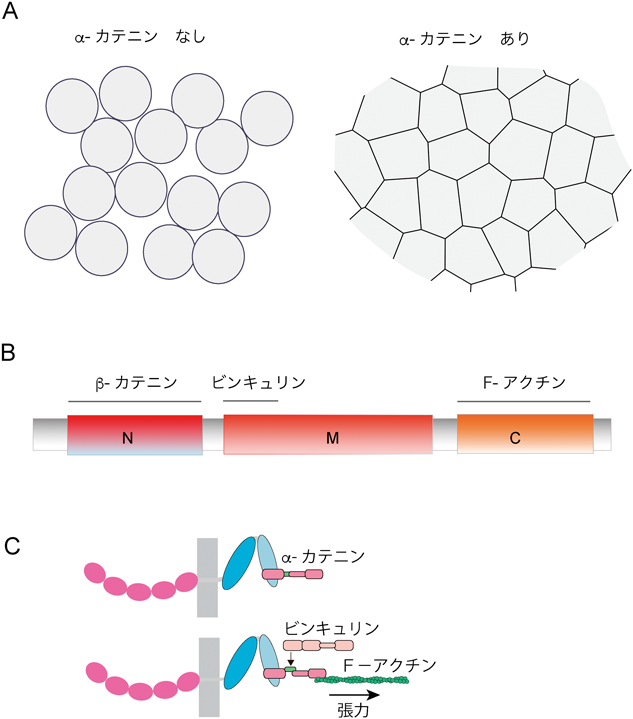
この細胞を使えば,αN-カテニンの機能が検証できると考え,廣橋さんに細胞を送っていただいた.PC9細胞は遺伝子トランスフェクションの効率が悪く,平野は少々苦労したが,αN-カテニンcDNAを導入処理した培養では,再現性よく細胞塊が生じ,カドヘリンが細胞接着のために働くためにはα-カテニンを必要とすることが初めて証明された16).ここで生じた細胞塊は,上皮特有の細胞配列を取り戻しており,たった一つの分子の存否だけで,「上皮か分散か」が決まるという結果は劇的だった(図2A).なお,ここでのβ-カテニンは,カドヘリンとα-カテニンの結合のための媒介役として働いているにすぎない.
ところで,この実験は幸運に恵まれたことが後でわかる.αN-カテニンの体における発現を調べると,分化した神経細胞に特異的で,上皮細胞は発現していない18).しかも,その後,新たに見つかったαE-カテニン欠失細胞株R2/7(大腸がん由来)を使い,これにα-カテニンを導入する実験を行うと,αE-カテニンは期待どおり上皮接着を回復させるものの,αN-カテニンにはまったく効果がない19).両カテニンの働きには細胞タイプ特異性があるらしい.PC9は上皮由来にもかかわらず,何らかの理由でαN-カテニンを受け入れることができたと推測されるが,真相は謎のままである.なお,PC9細胞は,αE-カテニンの導入によっても上皮構造を再構築することは後の実験により確認している20).
5. αE-カテニンの働き方:上皮と線維芽細胞では異なる
それでは,αE-カテニンは,どのようにカドヘリン接着を支えるのだろうか.月田博士とともに京都大学医学研究科に移った永渕らは,その機能の研究に着手していた.前述したように,αE-カテニンはβ-カテニンを介してE-カドヘリンに結合する.永渕らは,β-カテニンをすっとばして,αE-カテニンをE-カドヘリンに直接融合させ,このとき,種々のαE-カテニン変異体を用いることにより接着誘導に必要なドメインを探った.使った細胞は,線維芽細胞様性質を持つL細胞であった.実験の結果は,αE-カテニンのC末端ドメインが重要であることを示す21).C末端ドメインは,アクチンに結合することが知られると同時に,閉鎖帯(後述,図3A参照)の成分ZO-1にも結合するので,これらの分子との相互作用が,カドヘリン/カテニン複合体が接着分子として働くために重要であると結論している22).
ところが,上皮細胞を実験材料として用いると,違う結果が得られる.私の研究室では,大学院生の渡部光子が,上で述べたαE-カテニン欠失細胞株R2/7に種々のαE-カテニン変異体を導入し,上皮構造の回復に必要なドメインの分析を行った23).研究の過程で,αE-カテニンにビンキュリンが結合することを発見したが,別の研究室から類似の報告があり24),先を越されてしまう.しかし,ひるむことなく研究を続け,αE-カテニンとビンキュリンの結合こそが上皮接着構造形成のために必須で,このとき,ビンキュリンのアクチン結合ドメインが関与することを明確に示した.
ここで,αE-カテニンのドメイン構造を説明しておこう(図2B).αE-カテニンは大きく三つのドメインに分割され,N末端ドメインはβ-カテニンに結合,MドメインのN末端側でビンキュリンに結合し,C末端ドメインにはF-アクチンが結合する.L細胞を用いた研究ではC末端ドメインが重要である一方,上皮細胞を用いた研究ではビンキュリン結合ドメインが必要であると結論された.この違いを考察する前に,そもそも上皮と線維芽細胞とでは,接着構造が異なるので,両者の接着構造の特徴について概説しておこう.
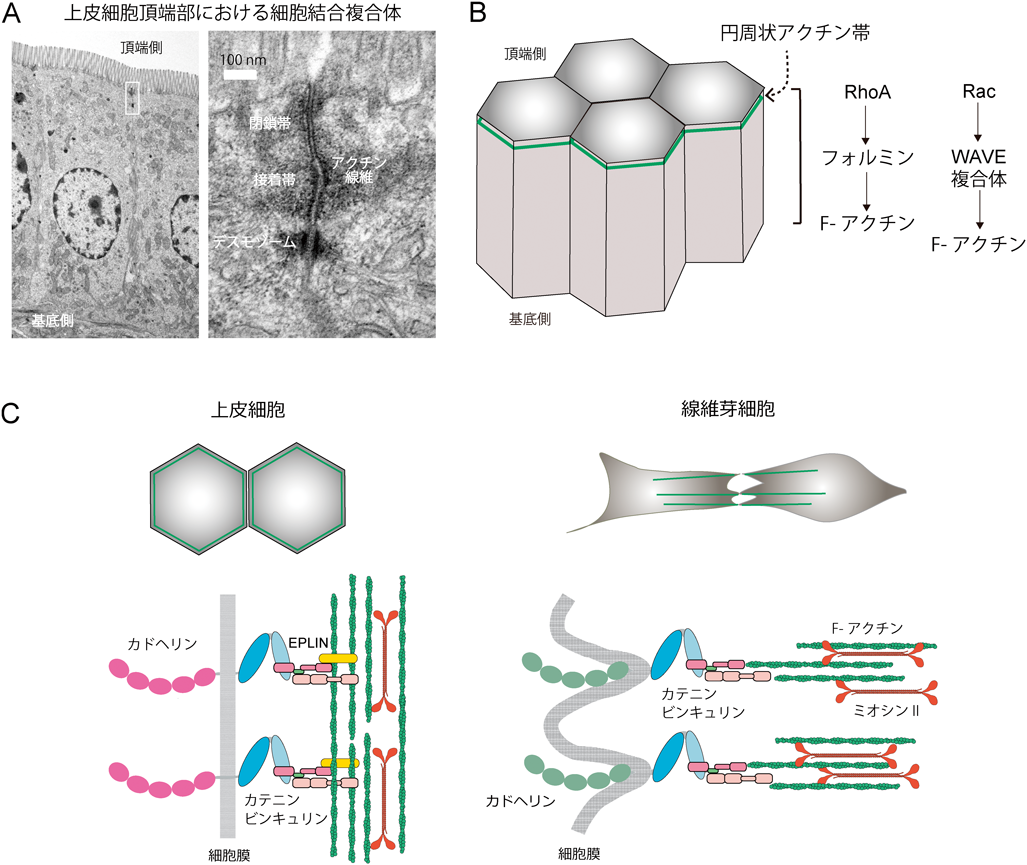
上皮といっても多様な組織型があり,ここでは単層上皮(腸粘膜上皮が代表例)を取り上げる.単層上皮細胞には必ず極性があり,上皮の内腔側を頂端側,細胞外マトリックスに接着する側を基底側と呼ぶ.頂端側には特殊な細胞間結合構造が発達しており,閉鎖帯[zonula occludens;密着結合(tight junction)ともいう]と接着帯(zonula adherens)が順に並んでいる(図3A).これらは,“zonula(小帯)”というように,細胞の頂端部をぐるりと取り巻いている.接着帯の直下には,接着斑(macula adherens;デスモソームともいう)があるが,接着帯とは異なり点刻状の構造である.閉鎖帯・接着帯・接着斑をまとめて「細胞結合複合体(junctional complex)」と呼び,カドヘリンが分布するのは接着帯である.閉鎖帯と接着帯の細胞質側にはアクチン束が集まっていて細胞膜と並行に走行しており,円周状アクチン帯(circumferential actin belt)を形成する(図3B).一方,線維芽細胞は極性があいまいで不定形だ.細胞間結合は帯状とはならずカドヘリンは点刻状に集まる25).このカドヘリン結合に対しアクチン束が細胞膜に直交するかたちでつながる(図3C).このように,上皮と線維芽細胞ではカドヘリンとアクチン束の空間的関係がまったく異なるが,カドヘリンによる接着構造を総称して“接着結合(adherens junction:AJ)”と呼ぶことが多い.αE-カテニンがないと,アクチン束はAJから離れるので,このカテニンが,カドヘリン系とF-アクチンを連結していることは間違いない.
ところで,上皮細胞におけるカドヘリン—カテニン複合体は,接着帯以外の細胞間接触面全体に分布しており,接着帯はむしろ特殊構造である.だが,接着帯こそが単層上皮層の構造的特徴を担っており,αE-カテニンがないときに顕著に壊れるのは接着帯である.そのとき,隣接する閉鎖帯が断片化し,細胞が互いに密着する上皮特有の構造が失われる23).
さて,L細胞と上皮細胞におけるαE-カテニンの関わり方の違いは,どのように説明できるのだろうか.よく考えれば,二つの研究における“接着の見方”が,違うことに気づく.渡部らの研究は,実は,閉鎖帯形成を接着の目安としており,ここで明らかになったのは,αE-カテニン—ビンキュリン複合体が,接着帯(閉鎖帯を支える役割を持つ)の形成に必要であるということだった.一方,接着帯を持たないL細胞の研究の方は,細胞間接着の強度を指標としたことから,接着帯と無関係なαE-カテニンの機能が洗い出され,これにはC末端ドメインが重要という結論に至った可能性が高い.興味深いことは,ビンキュリンが関わる場合でも,そのアクチン結合ドメインが重要であることだ.結局,いずれの場合もF-アクチンとの連携が大切で,F-アクチンの結合相手が細胞タイプによって違うだけのようにもみえる.
6. 細胞結合におけるアクチン線維の制御とその必要性
αE-カテニンを介してAJに結合するアクチン線維はどのような役割を持つのだろうか.この問題を研究するには,細胞結合に局在するアクチン制御因子を探し,その除去の効果を観察すれば何らかの情報が得られよう.Rhoファミリーのタンパク質群はアクチン制御シグナル系の上流に位置しており,その下流が広範に研究されてきた.細胞接着関連では,RhoA, Rac, Cdc42について研究が進み,これらの下流で働くWAVE/WASP—Arp2/3複合体系,フォルミン(formin)ファミリー,ROCK/Rhoキナーゼなどについて多数の論文が出版されている.全貌を解説する余裕はないので,我々の研究例をあげながら問題の要点をまとめよう.
RhoAの発現量をRNAi法で減らすと,上皮細胞間接着の全体が不調となり,その下流が正常な接着維持のために必要であることが示唆される26).RhoAのエフェクターの一つ,アクチン重合開始因子フォルミンファミリーについては複数の研究がある.まず,Dia1(Diaph1 diaphanous related formin 1).これを除去すると,接着帯が壊れるという27).活性化Dia1は細胞間結合に集まり,そこでのカテニンとアクチン線維の局在を増強することから,Dia1によって重合が促進されるアクチン線維が細胞間結合を維持するのだと議論されている(図3B).
上皮細胞の細胞結合複合体の下に広がる接着面(側面間接着)には,アクチン線維が無構造状に分布している.ここでは別のフォルミン,DAAM1が働いていることを理研研究員の西村珠子が明らかにしている26).DAAM1を除去すると,接着帯は影響を受けないまま,側面間接着の細胞膜の可動性が増大する.この細胞膜の可動性はRacエフェクターの一つWAVE複合体の働きによって支えられており,DAAM1による適切なアクチン重合が,WAVE複合体の作用に拮抗する形で接着面の安定化に寄与しているらしい(図3B).また,Cdc42に関しては,大学院生の大谷哲久が,Cdc42GEFの一つTubaが接着帯に分布しながら,細胞接着面全体のアクチン線維の分布を制御し,これが接着帯の形状にも影響することを明らかにしている28).以上のような研究から,細胞結合に分布するアクチンには複合的な役割があり,接着帯ではその維持に,側面接着では運動性の制御に関与している可能性がうかがわれる.ちなみに,側面間接着のF-アクチンについて,大学院生の亀谷祥子によるおもしろい観察がある.ここに分布するアクチン線維によってカドヘリン・カテニン複合体が,細胞下部から頂端部へ向かって輸送されるのだ29).カドヘリンフローと名づけたが,限定的な条件でしか観察されないので生理的意義は不明のままなのが残念である.
ところで,アクチン線維は,なぜ,接着帯では細胞膜に沿って配向し,線維芽細胞では細胞膜に直交するのだろうか.ヒントになる観察が行われている.大学院生の安部健太郎は,αE-カテニンに結合するタンパク質を探索する中で,偶然,EPLIN(epithelial protein lost in neoplasm)を発見した30).EPLINはアクチン線維を束ねる働きを持つタンパク質である.一連の研究の結果から,EPLINはαE-カテニンのC末端領域に結合して,カドヘリン—カテニン複合体にアクチン線維を連結する役割があると推論した.その後に参加した理研研究員の田口勝敏は,上皮細胞におけるEPLINの役割をより詳しく研究し,EPLINを除去すると,アクチン束の配向が変わり,線維芽細胞の場合のように,膜に直交するよう変化することを見つけた31).同時に,細胞全体が激しく変形するようになった.この結果から引き出されたモデルは,EPLINがあればアクチン束が細胞膜に沿ってつなぎ止められ,接着帯が安定化するというものである(図3C).ただ,細胞のタイプによってEPLIN除去に反応しない場合もみつかり(未発表),このモデルのさらなる検証が必要であると感じている.なお,EPLINは,その名の由来のとおりがんで減少するとされ,EPLIN消失による細胞接着の不安定化と,がん進展との関係は興味深い問題である32).
次に,カドヘリン系と結合するアクチン束には,形態形成上,積極的な役割があることを解説しよう.接着帯に沿う円周状アクチン束にはミオシンIIが結合しており,収縮することができる.上皮細胞を上面からみると,接着帯がピンと張っていることがあり,この場合,アクトミオシンが収縮して接着面に張力を与えている可能性が高い.ただし,上皮の接着面はいつも緊張しているわけではなく,湾曲したり,ジグザクと入り組んだりすることもよくある.ミオシンIIの活性化阻害剤(ROCK阻害剤など)を与えても,接着面の形状が影響を受けないことがしばしばで,ミオシンIIが,常時,活性状態にあるわけではないようだ.しかし,動物の形態形成のためには,この接着帯アクトミオシンの収縮が重要な役割を果たす.発生中の胚の上皮は,組織や器官を形づくるために変形するが,一番単純な例が,湾曲,または,折れ曲がりである.以下に我々の研究例を紹介する.
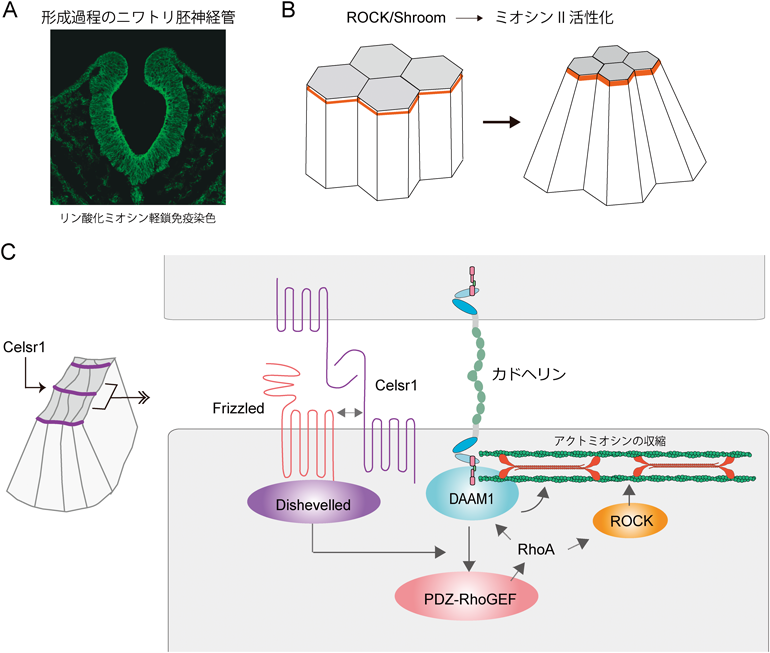
中枢神経系の形成は,外胚葉から生ずる神経上皮層が体内に「陥入」し,神経管を作ることから始まる(図4A).このとき,神経上皮の頂端部(体表面側)が収縮し,個々の細胞の形がくさび状となり,この変化が細胞層を湾曲・陥入させる原動力となる.この収縮こそ接着帯アクトミオシンの収縮によるのだ(図4B).どのように,接着帯アクトミオシンが積極的に収縮するのだろうか.1999年,P. Soriano博士の研究室で,神経管の形成に不全がある遺伝子変異マウスが発見され,その原因遺伝子がShroomと命名された33).Shroomタンパク質は接着帯領域に局在することから,上記の西村珠子がその働き方に興味を持ち研究した結果,ShroomはROCKを接着帯に集め,このROCKがミオシンIIを活性化することが明らかとなった34).接着帯アクトミオシンを活性化する機構が存在したのである.一方,ROCKの接着帯への集積を抑える機構が存在することも,大学院生の石内崇士が明らかにしている.上皮頂端部には,「Par複合体」の成分aPKC(atypical protein kinase C)が局在するが,aPKCはROCKをリン酸化することにより,細胞結合への局在を阻害する35).上皮細胞は必要に応じてROCKの局在を操り,細胞変形に利用しているようだ.
ところで,神経上皮層が湾曲して管を形成するためには,湾曲の方向に極性がなければならない.すなわち,体を横断する軸に沿ってだけ曲がり,体の前後軸方向には曲がらない.西村は,この「平面内極性(planar cell polarity:PCP)」問題を明らかにするため,陥入しつつある神経上皮層を胚の上部から観察してみた.すると,ミオシンIIの活性化は,接着帯全面で起きているわけではなく,体の横断軸に沿った部分に限局される傾向があった36).この接着帯アクトミオシン収縮の方向の偏りを前提とすれば,上皮層の管状変形がうまく説明できる.そして,この偏りはPCP機構に依存するはずであると考え,PCP制御因子の関わりを調べたところ,以前,大学院生の碓井理夫らが発見したFlamingo(カドヘリンスーパーファミリーに属する分子)の脊椎動物ホモログの一つCelsr1が関与することが明らかとなった37).西村の観察によれば,Celsr1も横断軸に向く接着帯に偏在し,ここでDishevelledを介してRhoAを活性化させ,その結果,当該部分に局在するアクトミオシンを選択的に収縮させる(図4C).これで,神経上皮が管を形成するための機構の一端がわかったが,Celsr1がどのようにして特定の接着帯に局在するのかについては今なお不明である.なお,同様な研究は,無脊椎動物(特にショウジョウバエ)を用いて盛んに行われ,細胞間結合の収縮が細胞の再配列,細胞層変形の原動力となるという考えは,広く受け入れられている1).
8. αE-カテニンとアクチン相互作用のメカノバイオロジー
αE-カテニンにF-アクチンが結合するという通説が覆される事件があった.2005年,スタンフォード大学のJ. Nelson研究室は,カドヘリン—カテニン複合体に,F-アクチンは結合しないという報告をする38).in vitro再構成系で行われた実験である.数年間,論議を呼んだが,光トラップ法による生物物理的研究により,αE-カテニン—アクチン線維系に張力さえ働けば,両者はin vitroでも結合するという発見があり,一件落着となる39, 40).個々の観察はたぶん正しく,αE-カテニンとF-アクチンの相互作用には「力」が必要であるという点が重要だ.
αE-カテニンとビンキュリンの結合も力によって制御されていることが,理研の米村重信博士のグループによって明らかにされた41).αE-カテニンに結合するアクチン線維束はミオシンIIによって収縮し,αE-カテニンを引っぱる.この力により,αE-カテニンン上のビンキュリン結合ドメインが開き,ビンキュリンが結合するというわけだ(図2C).このモデルは立体構造学的にも確認され,αE-カテニンの働きを議論する上で重要となっている40).
細胞間の接着部位は,接着し合う二つの細胞が互いに力を及ぼし合う場所で,ここに集積する分子群はその影響下にあり,αE-カテニンの構造がF-アクチンを介した張力によって変化するのはその典型例といえる.同様な力は,細胞-基質接着に対しても働き,インテグリンが働く接着構造(focal adhesion)に集積するビンキュリンについては,クモ糸のタンパク質を分子内に挿入することにより,張力による構造変化が見事に示されている42).機械的な力がどのような生理的効果を及ぼすかについては,メカノバイオロジーとして盛んに研究されている先端分野だが15, 43),αE-カテニンとF-アクチンの相互作用は,力の受容システムとしての役割を果たしている可能性がきわめて高い.次に,αE-カテニンとF-アクチンの間に働く機械力が,実際に,細胞結合に大きな効果を及ぼすことを示した我々の研究例を紹介しよう.
すでに述べたように,がん由来の細胞株には,αE-カテニンを失うなどの接着異常が認められ,がんの悪性化との関係が疑われてきた.しかし,がん細胞株の中には,カドヘリンもカテニンも正常に発現しているにもかかわらず上皮型接着ができない細胞株が見つかる.その一つは,大腸がん腹水から樹立されたColo205で,細胞が分散した状態で増殖する44).カドヘリンとカテニンを発現しているなら,何らかの手法で接着を回復させ,がん細胞の拡散を抑制できるのではないかと期待し,1990年代に研究を始めた.
先行する研究から,カテニンのリン酸化がカドヘリン接着に影響する可能性が示唆されていた.そこで,大学院生の青野真也が,Colo205細胞にプロテインキナーゼ阻害剤の効果を調べる実験に着手し,実際,スタウロスポリンに接着誘導効果があることを見いだした.そこで,スタウロスポリン処理後のカテニンを調べると,αE-カテニン,β-カテニンには変化がなかったが,もう一つのカテニン,p120-カテニンの電気泳動速度が変わり,リン酸化状態が変化している可能性が示唆された.p120-カテニンは,v-Srcの標的の一つとして発見され45),その後,カドヘリン細胞質領域の膜貫通ドメインに近い場所(juxta-membrane domain)に結合する分子であることが明らかにされている(図1A)44).p120-カテニンの関与を検討するため,種々の欠失変異体を作製し,Colo205細胞に導入してみると,N末端側ドメインを欠いた変異体が細胞接着を回復させた.さらに,p120-カテニンが結合できないカドヘリン変異体を過剰発現させても同様な効果が得られた.これらの結果から,p120-カテニンが,N末端側ドメインを介してカドヘリン接着を阻害していると推論された.
この発見を発展させるべく研究を続けたが遅々として進まない.その間,正常細胞におけるp120-カテニンの機能が,他の研究室で明らかにされていく.この分子を線維芽細胞で欠失させるとRhoAの活性が上がるので,正常では,RhoAを抑制する作用があると推測される46).また,p120-カテニンがないと細胞膜上のカドヘリンが不安定になることなども明らかになる47).しかし,これらの観察と,Colo205細胞の接着に影響を与える活性との関係がわからないまま,あるとき,大学院生の一居哲夫がp120-カテニンを神経芽腫細胞株で過剰発現させたところ,微小管上に分布することを偶然発見した48).この観察に興味を覚えた理研研究員Wenxiang Mengが,p120-カテニンは何らかの分子を介して微小管に結合するのではないかと推定し,候補分子を質量分析法により探索して,PLEKHA7という機能未知の分子を見つけた.さらに,PLEKHA7に結合する分子を探し,無名のタンパク質に行き当たる49).非常に興味深いことに,この分子は微小管のマイナス端に結合し,中心体とは独立して,微小管の伸張の場を提供していることがわかった.そして,中国の民話に登場する哪吒(Nezha)と命名した.Nezhaは,槍(=微小管)を持って振り回すからである.この分子は,その後,CAMSAP3と再命名され,微小管マイナス端の新しい制御因子としての地位を確立し,微小管分野に大きく貢献することになる50).
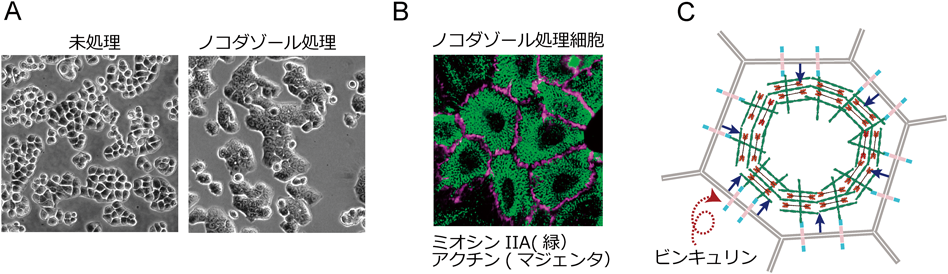
p120-カテニン問題は道をそれてしまったが,Nezha/CAMSAP3の研究に参加した大学院生の長江成典が,HeLa細胞を使ってNezha/CAMSAP3を除去すると,微小管が再編成され,その結果,GEF-H1が活性化されてRhoAの活性が上がることを示唆していた51).GEF-H1は微小管によって活性が抑制されるRhoGEFである52).ここで,p120-カテニンの働き方として,p120-カテニン→Nezha/CAMSAP3→微小管→RhoA抑制という経路が浮上し,がん細胞接着異常におけるp120-カテニンの関与を調べる際,微小管もチェックしたらどうかということになった.このころ,理研研究員となった伊藤祥子が,がん細胞の接着異常問題のテーマを引き継ぐ.伊藤は,Colo205と同様に閉鎖帯/接着帯の形成不全を示すHT29細胞を使い,微小管重合阻害剤ノコダゾールを与えてみた.すると,劇的な効果があり,細胞間接着が速やかに回復した(図5A)53).ノコダゾールはよく効いたが,直感的に選んだ薬剤にすぎず,先入観なしの有効化合物のスクリーニングが必要と考え,理研の創薬・医療技術基盤プログラムに参加した.プログラムのスタッフに10万を超える化合物を検査していただいた結果,驚くべきことに,大多数の有効物質は微小管脱重合活性を示した.こうなると,ノコダゾールを使い,その働きを調べるしかない.
微小管を脱重合させると何が起きるのか,予想どおり,GEF-H1に依存してRhoA活性が上昇した.そして,RhoAが接着の回復のために必要であることも確認できた.しかし,どうして接着が回復するのか,答えが得られないまま時が過ぎる.あるとき,伊藤が,定例の研究室セミナーで,GFP標識したミオシン軽鎖の動画をみせた.なんと,ノコダゾールを与えると,細胞頂端部の皮質にあるミオシンIIAのネットワークが,同心円状に再編成され収縮するのだ(図5B).「これでいける」と思った瞬間である.ここで,微小管脱重合→GEF-H1活性化→RhoA活性上昇→皮質アクトミオシンの収縮,という図式ができあがった.次は,収縮アクトミオシンの役割を明らかにすればよい.ただ不思議なのは,これまで多くの細胞でみてきた事実と異なり,ミオシンIIAの分布は細胞接着部位と重ならないのだ.常識が通用しない現象に遭遇すると説明に窮するが,新発見のチャンスでもある.とりあえず,皮質アクトミオシンの収縮が接着を誘導しているに違いないと考え観察を続けると,確かにミオシンIIAは皮質にしかないが,これに絡まるアクチン繊維は細胞接着部位に達している(図5B).しかも,そのアクチン線維はミオシンの働きにより強く引っぱられていた.
この張力こそが重要だと推定し,その受容機構を探る.ここでビンキュリンの再登場だ.ビンキュリンが張力に依存してαE-カテニンに結合し,αE-カテニン—ビンキュリン複合体が接着帯形成のために必要であることをすでに述べた.未処理のHT29細胞では,軽く接着し合っている細胞間の境界に,E-カドヘリンとαE-カテニンが集まっているが,ビンキュリンはまったく検出されない.しかし,ノコダゾール処理によって接着部位にビンキュリンが集まった.そして,このビンキュリンはミオシン阻害剤処理により消失した.そうなると,次の図式が想定されよう.皮質アクトミオシン収縮→αE-カテニンに連結したアクチン線維の引張り力の増大→ビンキュリンの再分布→接着の回復,である.ここで,ビンキュリンがどのような働きをしているのかについて全貌は解明できていないが,ビンキュリンの集積とともにPDZ-RhoGEFが集積し,その下流では,RhoA→LIMキナーゼ→コフィリンの阻害→アクチン重合というシグナル系が働くことを示すことができた53).
以上,明らかになったシグナル系から想像できるように,RhoAの活性を上昇させることが鍵である.実際,RhoGAPを除去することにより,活性化型RhoAの相対量を増やしてやれば,微小管阻害剤を使わなくても接着誘導できることを確認した.実は,微小管阻害とは,RhoA活性を操作するための一手段にすぎなかったのだ.なお,HT29細胞の接着を「回復」させることができたが,正常な上皮接着とは少々異なる.正常上皮では,繰り返し述べたように,細いアクトミオシン束が接着帯細胞膜に平行に並ぶが,接着回復したHT29細胞では,アクトミオシン束が幅広い帯状を呈する(図5B).このがん細胞は,接着帯を形成するためにより強力な収縮力を必要とすることを物語っている.他の複数の大腸がん由来細胞にもノコダゾール感受性があることが確認され,少なくとも特定のがん細胞群の間で共通する異常らしい.
ところで,以上の研究は,「p120-カテニン→Nezha/CAMSAP3→微小管→RhoA抑制」という仮説が動機づけになっていたが,それなら,HT29細胞では,p120-カテニン,または,Nezha/CAMSAP3の発現や機能に異常があるのだろうか? 残念ながらこの点については検討が不十分なまま研究が終了している.また,理研の創薬・医療技術基盤プログラムへの参加は,医療にも有用な化合物を発見することが本来の目的だったが,ヒットしたのは微小管阻害剤ばかりで,この点に関しては残念な結果に終わった.
カテニン群は,カドヘリンの働きをサポートするための細胞質因子だが,β-カテニンが転写制御因子としても働くように,機能のレパートリーは単純ではない.実際,αE-カテニンについても,カドヘリンとは独立した機能があることが以前から指摘されてきた.カドヘリン—β-カテニン複合体から遊離したαE-カテニンは二量体を形成し,そのC末端ドメインを介してアクチンを制御するという54).一方,我々の最近の研究は,これとはまた別の機能を明らかにしている.
腸神経系は,発生期の腸管上を集団移動する神経冠細胞から生じる.理研研究員Vassil Vassilevは腸神経冠細胞の集団移動の仕組みに興味を持ち,カドヘリンによる細胞間接着が細胞移動に関与するかどうか検討した55).神経冠細胞が発現するN-カドヘリンを除去した場合,遊走に遅れが生じるものの致命的な影響は観察されなかった.しかし,β-カテニン,αE-カテニンを除去した場合には,ともに,著しい遊走遅延が生じた.動画を撮ると,正常な腸管では,神経冠細胞が互いにつながり一方向に移動しているのだが,カテニンがないと細胞がバラバラとなり移動の方向性が失われる.ある意味,期待された結果であって,細胞が集団移動するためには,互いの接着が必要であることが確認された(と思った).ところが,神経冠細胞を「単独で」培養しても,正常細胞は方向性移動をするのに対し,カテニン欠失細胞は方向性に異常を来すのである.つまり,細胞間接着とは無関係な機能がカテニンにあると推測された.同時期,理研研究員Anna Platekが,別の研究プロジェクトのため,U251細胞(グリオーマ細胞株)を使ってαE-カテニン欠失細胞の動画を撮っていた.この場合も細胞の動きがおかしい.共通の現象に違いないと考えて,二つの細胞系をまとめて研究することにした.
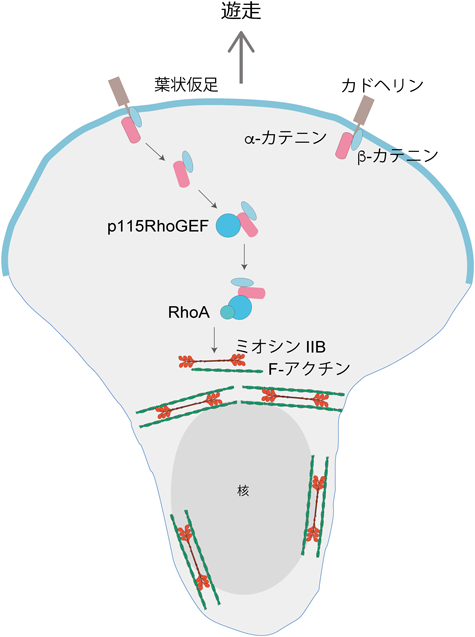
細胞が前進するとき,前後の極性が生じ,先端は運動装置の一つ,葉状仮足(lamellipodium)を形成する.カテニン欠失細胞は葉状仮足を形成するものの,その位置が不安定で,これが直進的な細胞移動を阻害するのではないかと考え,次に,カテニンが単細胞上のどこに分布するのか調べてみた.そうすると,まず,葉状仮足に現れ,次いで核の周辺部に運ばれることがわかった(図6).普通なら,たんに分子のリサイクルをみているに違いないと無視する現象だが,この分子局在の意味を調べることにした.そして,以下の筋書きが明らかになった.まず,β-カテニン,αE-カテニンはカドヘリンとともに,葉状仮足の後方から細胞内に取り込まれ,途中でカドヘリンと解離するらしい.このとき,β-カテニンのチロシン654のリン酸化(pY654)が必要で,リン酸化できない変異β-カテニンを細胞に導入すると,αE-カテニンの細胞内分布も,細胞の直進も撹乱される.それでは,移動したβ-カテニン—αE-カテニン複合体は何をしているのか.
細胞が移動するためには,前進しながら後方を引っぱる必要がある.このためには,後方部の収縮が重要で,RhoAに依存したミオシンIIの関与が想定されていた.そこで,活性化RhoAの分布を,RhoA-GTPに結合するマーカーを使って調べると,核の周辺部に集まっていることがわかった.そして,ミオシンIIBを含むアクトミオシン束が,核を包むように後方に移動しており,一方,カテニン除去細胞の場合には,活性化RhoA,および,ミオシンIIBの分布が撹乱されていた.こうなると,カテニンは,RhoAの分布を制御しているのではないかという可能性が浮上し,調べていくと,核周辺部のαE-カテニンはp115RhoGEFと結合しており,p115RhoGEFを除去すると核前方の活性化RhoAが消えるなど,前後極性がおかしくなることもわかった.さらに,αE-カテニンがミオシンIIBと結合することも判明した.まとめると,αE-カテニンは,β-カテニンに助けられて核周辺部に達し,p115RhoGEFと結合して活性化RhoAをこの領域に局在させ,次いで,ミオシンIIBを活性化する(図6).αE-カテニンがないとp115RhoGEF/RhoAの細胞内分布が定まらず,細胞前後軸がうまく形成されない.これが現在の我々のモデルである.あいまいな点が多いが,今後の検証を待ちたい.
αE-カテニンのカドヘリンと独立した機能については,進化的に考えるとおもしろい.接着分子としてのカドヘリン-カテニン複合体は,調べられた限り,多細胞動物すべてに存在する.ところが,その原始型とされる単細胞動物,立襟鞭毛虫は,カドヘリン様分子を持つのにカテニンを持たない.一方,植物として分類される細胞性粘菌は,カドヘリンを持たないにもかかわらず,β-,および,α-カテニンのホモログを発現する56–58).α-カテニン欠失粘菌では,細胞間接着は正常だが,細胞極性に問題があるという59).どうやら,カテニン群は,元来,カドヘリンとは独立した存在で,あるときカドヘリンと出会い,細胞間接着装置を生み出したという空想が成り立ちそうだ54).
中枢神経系におけるシナプス結合は,神経間伝達のための中心的な場だが,教科書では細胞膜と細胞膜の間に何も描かれていないことが多い.シナプス結合の形成機構を明らかにすることは神経生物学にとっても重要なはずで,私は,シナプス結合とAJの構造的な類似性から,ここにもカドヘリンが関与するのではないかと永らく考えていた.しかし,確実な検証のためには電子顕微鏡が必須で,容易には手が出せないでいた.修士課程でαN-カテニンの組織内分布の研究を終えていた内田直磁に,シナプスに分布するかどうか調べたらどうかと持ちかけたところ,同意してくれた.彼は,月田研究室にお世話になって電子顕微鏡技術を習得し,αN-カテニンのみならずN-カドヘリンがシナプスに局在することを見事に示した60).ただし,シナプス全域に分布するわけではなく,シナプス小胞から伝達物質が放出される,いわゆる「活性部位(active zone)」を避け,その両脇を固めるように分布していた.
ところで,これが世界初のシナプス結合分子の同定になるはずだったが,そうはいかなかった.この研究がまとまりかけていたころ,ワシントン大学J. Sanes博士の研究室(現在はハーバード大学)の山形方人さんが,N-カドヘリンが網膜のシナプスに局在することを発表していることに気づいた61).しかし,予備的な報告だったため,内田論文は迷わずNeuron誌に投稿した.ところがリジェクションされてしまう.直ちに,Journal of Cell Biology(JCB)誌に投稿してこちらは無事にアクセプトされた.その間,あろうことか,類似の論文がNeuron誌に掲載された.事後談だが,JCBでレビューをしてくれた某博士が,この成り行きをみていて,Neuron誌のエディターに抗議してくれたという.このエディター氏,気になったとみえ,会うごとに本件を話題にしていた.彼がいうには,私がもっと“persist”すべきだったという.いろいろあったが,カドヘリン/カテニンがシナプスに分布することを内田が得た高質の画像を用いて証明できた.
次は,カドヘリン/αN-カテニンがシナプスで果たす役割を調べる実験だ.手っ取り早い方法は,神経細胞を培養しカドヘリン/αN-カテニンを阻害してやればよい.この系を使った実験には,大学院生の富樫英,安部健太郎が着手した.当時,まだRNAi法は普及しておらず,特定の分子の機能を阻害するには工夫が必要であった.幸い,カドヘリンについては,大学院生の藤森俊彦がドミナントネガティブ変異体を先だって開発しており,また,αN-カテニンについては,助教授の千坂修がノックアウト(KO)マウスを作製してくれた.詳細は省くが,カドヘリン/αN-カテニンを阻害すると,興奮性シナプスを形成するスパインが不安定になり,その結果,シナプスに関わる諸現象が影響を受ける62, 63).シナプス形成も細胞接着現象の一つであって,カドヘリン/αN-カテニンの支配下にあることを示すことができた.なお,この研究を通じて,シナプス活性部位には別の接着分子が存在することが予測されたが,T. Südhof博士の研究室で同定されたneurexinやneuroliginなどが該当するのだろう64).
培養細胞を用いた研究の後に待っているのは,生体における生理機能を明らかにすることだった.しかし,N-カドヘリンKOマウスは発生途上で死に,αN-カテニンのKOマウスは成熟できず,コンディショナルKOを駆使しない限りシナプス機能は研究できない.一方,当時,タイプIIとして分類されるカドヘリンのサブタイプ(カドヘリン6, 8, 11)が神経回路に相関して発現することを見つけ,研究の焦点をこちらに合わせ,KOマウスを作製して表現型を解析した65).これらのマウスは死なないが,表現型は微妙なため,研究は苦戦.それでも,カドヘリン8については,大学院生の鈴木祥宏が,痛覚生理学が専門の古江秀昌博士の協力をいただき,低温感受性の感覚神経と脊髄神経との間のシナプス連絡のために重要であることを示すことができた66).
ところで,αN-カテニンのKOマウスは,プルキンエ細胞の前駆細胞が正常に移動できないなど,シナプス形成以前の発生障害をみせる63).理研テクニカルスタッフだった上村允人が網羅的な表現型解析を行いさまざまな異常を記載しているが67),その中に,前交連神経束の伸張異常というのがあり,プルキンエ前駆細胞の遊走異常とともに,細胞移動に関係した欠陥が認められた.αE-カテニンが細胞の方向性移動に関わるという発見と考え合わせると興味深い.αN-カテニンがカドヘリンと独立した機能を持つかどうかは未検証である.
カドヘリン接着を支える細胞内機構について,私の研究室における発見を中心に解説させていただいた.カドヘリンはその機能を発揮するためにカテニン群を必要とする.
ところで,カテニンは,すべての「カドヘリン」に結合するわけではない.カドヘリンは100以上のメンバーからなるスーパーファミリーを形成しており,すべて,細胞外領域のホモフィリックまたはヘテロフィリックな結合を介して機能する分子だが,細胞質領域は多様化しており,カテニンが結合できるのはE, N-カドヘリンを含む「クラシックカドヘリン」と小分類される分子群だけである.他の分子は,平面内極性,細胞増殖制御,内耳不動毛や微絨毛の配列,細胞間認識などに働くなど,機能が多様化しており,デスモソームカドヘリンを除いて,強力な細胞間結合には関わらない.カテニンの存在がカドヘリンの細胞間接着機能にとっていかに重要かわかるだろう.裏方のカテニンこそが主役といってもよいほどである.
それでは,カテニンの役割とは結局何なのか.上皮などの組織を作るためには,細胞を密に接着させる必要がある.細胞を解離してみるとわかるように,個々の細胞には丸くなろうとする力が働いている.表面張力に加え,表層部骨格系による収縮力などが関与するのだろう.接着分子は,このような細胞を丸くする力に抗して,細胞の膜をファスナーのようにつなぎ止めねばならない.これには,相応の力が必要なはずだ.これまでの観察を総合すれば,この力は,カテニンと,それに結合したアクチン線維が発揮していると推測される.
そして,このような複数成分からなる「装置」を作ったがゆえに,細胞接着は状況に応じて変化でき,一方,部品の欠陥は組織崩壊をもたらす.しかし,この装置がどのように働いているのかについて,残念ながら完全な理解には至っていない.研究における分解的手法はそろそろ限界に近づいており,今後の展開としては,「作ってみる」ことが必要なのではないかと感じている.人工細胞接着装置だ.仕組みの理解は作ってみることにより深まるのではないだろうか.
上で言及したカドヘリンスーパーファミリー分子群については豊富な話題があるのだが,本稿のテーマから離れるため省いた.私の研究室では,碓井,上村らによる平面内極性制御因子“Flamingo”の発見68),田ノ上,石内,塚崎らによるFatカドヘリンの研究69–71),平野,上村,中尾,林らによるδ2-プロトカドヘリンの研究72, 73),などがあり,またの機会に紹介したい.また,カドヘリン結合に隣接した閉鎖帯やデスモソームの構造と機能についても広く研究されており,細胞結合全体を語るには必須なのだが,話が膨大になりすぎるため省いた.
私がまだ細胞表面の問題だけを研究していたころ,内田驍博士(故人;当時,大阪大学細胞工学センター教授)と,カリフォルニア・モントレーの海岸をラッコ見物のために歩きながら,研究の将来について語り合ったことがある.このとき,私は細胞の中の機械仕掛けを探るような研究には興味がないと,つい軽口を叩いてしまい,内田さんから自分は機械仕掛け問題が好きと,皮肉っぽくやりかえされたことがある.その後の私は,いつの間にか,内田さんタイプの研究にどっぷりはまってしまった.言い訳をしたいのだが叶わない.細胞内機構の謎解きは,楽ではないが,限りなくおもしろい.
謝辞Acknowledgments
ここに記載した研究は,京都大学大学院理学研究科および生命科学研究科に所属した大学院生,職員,そして,理化学研究所の研究員,テクニカルスタッフの皆様によって達成されました.ここに感謝いたします.字数の制約上,すべての関連研究について言及できなかったことはお詫びします.また,本稿に記載した研究は,文部科学省/日本学術振興会からの科学研究費,理化学研究所の内部研究費などによって支えられました.あらためて感謝いたします.
引用文献References
1) Takeichi, M. (2014) Dynamic contacts: Rearranging adherens junctions to drive epithelial remodelling. Nat. Rev. Mol. Cell Biol., 15, 397–410.
2) Takeichi, M. (1988) The cadherins–cell–cell adhesion molecules controlling animal morphogenesis. Development, 102, 639–655.
3) Nagafuchi, A., Shirayoshi, Y., Okazaki, K., Yasuda, K., & Takeichi, M. (1987) Transformation of cell-adhesion properties by exogenously introduced E-cadherin cdna. Nature, 329, 341–343.
4) Nagafuchi, A. & Takeichi, M. (1988) Cell binding function of E-cadherin is regulated by the cytoplasmic domain. EMBO J., 7, 3679–3684.
5) Nagafuchi, A. & Takeichi, M. (1989) Transmembrane control of cadherin-mediated cell-adhesion—a 94 kda protein functionally associated with a specific region of the cytoplasmic domain of e-cadherin. Cell Regul., 1, 37–44.
6) Ozawa, M., Ringwald, M., & Kemler, R. (1990) Uvomorulin catenin complex-formation is regulated by a specific domain in the cytoplasmic region of the cell-adhesion molecule. Proc. Natl. Acad. Sci. USA, 87, 4246–4250.
7) Herrenknecht, K., Ozawa, M., Eckerskorn, C., Lottspeich, F., Lenter, M., & Kemler, R. (1991) The uvomorulin-anchorage protein α-catenin is a vinculin homologue. Proc. Natl. Acad. Sci. USA, 88, 9156–9160.
8) Cowin, P., Kapprell, H.P., Franke, W.W., Tamkun, J., & Hynes, R.O. (1986) Plakoglobin: A protein common to different kinds of intercellular adhering junctions. Cell, 46, 1063–1073.
9) Mccrea, P.D., Turck, C.W., & Gumbiner, B. (1991) A homolog of the armadillo protein in Drosophila (plakoglobin) associated with E-cadherin. Science, 254, 1359–1361.
10) Peifer, M. & Wieschaus, E. (1990) The segment polarity gene armadillo encodes a functionally modular protein that is the Drosophila homolog of human plakoglobin. Cell, 63, 1167–1178.
11) Su, L.K., Vogelstein, B., & Kinzler, K.W. (1993) Association of the APC tumor suppressor protein with catenins. Science, 262, 1734–1737.
12) Rubinfeld, B., Souza, B., Albert, I., Muller, O., Chamberlain, S.H., Masiarz, F.R., Munemitsu, S., & Polakis, P. (1993) Association of the APC gene product with beta-catenin. Science, 262, 1731–1734.
13) Nagafuchi, A., Takeichi, M., & Tsukita, S. (1991) The 102 kd cadherin-associated protein: Similarity to vinculin and posttranscriptional regulation of expression. Cell, 65, 849–857.
14) Hirata, H., Samsonov, M., & Sokabe, M. (2017) Actomyosin contractility provokes contact inhibition in E-cadherin-ligated keratinocytes. Sci. Rep., 7, 46326.
15) Furukawa, K.T., Yamashita, K., Sakurai, N., & Ohno, S. (2017) The epithelial circumferential actin belt regulates Yap/Taz through nucleocytoplasmic shuttling of merlin. Cell Rep., 20, 1435–1447.
16) Hirano, S., Kimoto, N., Shimoyama, Y., Hirohashi, S., & Takeichi, M. (1992) Identification of a neural α-catenin as a key regulator of cadherin function and multicellular organization. Cell, 70, 293–301.
17) Shimoyama, Y., Nagafuchi, A., Fujita, S., Gotoh, M., Takeichi, M., Tsukita, S., & Hirohashi, S. (1992) Cadherin dysfunction in a human cancer cell line: Possible involvement of loss of α-catenin expression in reduced cell-cell adhesiveness. Cancer Res., 52, 5770–5774.
18) Uchida, N., Shimamura, K., Miyatani, S., Copeland, N., Gilbert, D., Jenkins, N., & Takeichi, M. (1994) Mouse αN-catenin: Two isoforms, specific expression in the nervous-system, and chromosomal localization of the gene. Dev. Biol., 163, 75–85.
19) Ishiyama, N., Tanaka, N., Abe, K., Yang, Y.J., Abbas, Y.M., Umitsu, M., Nagar, B., Bueler, S.A., Rubinstein, J.L., Takeichi, M., et al. (2013) An autoinhibited structure of α-catenin and its implications for vinculin recruitment to adherens junctions. J. Biol. Chem., 288, 15913–15925.
20) Watabe, M., Nagafuchi, A., Tsukita, S., & Takeichi, M. (1994) Induction of polarized cell-cell association and retardation of growth by activation of the E-cadherin catenin adhesion system in a dispersed carcinoma line. J. Cell Biol., 127, 247–256.
21) Nagafuchi, A., Ishihara, S., & Tsukita, S. (1994) The roles of catenins in the cadherin-mediated cell adhesion: Functional analysis of E-cadherin–α-catenin fusion molecules. J. Cell Biol., 127, 235–245.
22) Imamura, Y., Itoh, M., Maeno, Y., Tsukita, S., & Nagafuchi, A. (1999) Functional domains of α-catenin required for the strong state of cadherin-based cell adhesion. J. Cell Biol., 144, 1311–1322.
23) Watabe-Uchida, M., Uchida, N., Imamura, Y., Nagafuchi, A., Fujimoto, K., Uemura, T., Vermeulen, S., Van Roy, F., Adamson, E.D., & Takeichi, M. (1998) α-catenin–vinculin interaction functions to organize the apical junctional complex in epithelial cells. J. Cell Biol., 142, 847–857.
24) Weiss, E.E., Kroemker, M., Rudiger, A.H., Jockusch, B.M., & Rudiger, M. (1998) Vinculin is part of the cadherin-catenin junctional complex: Complex formation between alpha-catenin and vinculin. J. Cell Biol., 141, 755–764.
25) Yonemura, S., Itoh, M., Nagafuchi, A., & Tsukita, S. (1995) Cell-to-cell adherens junction formation and actin filament organization: Similarities and differences between non-polarized fibroblasts and polarized epithelial cells. J. Cell Sci., 108, 127–142.
26) Nishimura, T., Ito, S., Saito, H., Hiver, S., Shigetomi, K., Ikenouchi, J., & Takeichi, M. (2016) DAAM1 stabilizes epithelial junctions by restraining WAVE complex-dependent lateral membrane motility. J. Cell Biol., 215, 559–573.
27) Carramusa, L., Ballestrem, C., Zilberman, Y., & Bershadsky, A.D. (2007) Mammalian diaphanous-related formin dia1 controls the organization of E-cadherin-mediated cell-cell junctions. J. Cell Sci., 120, 3870–3882.
28) Otani, T., Ichii, T., Aono, S., & Takeichi, M. (2006) Cdc42 gef Tuba regulates the junctional configuration of simple epithelial cells. J. Cell Biol., 175, 135–146.
29) Kametani, Y. & Takeichi, M. (2007) Basal-to-apical cadherin flow at cell junctions. Nat. Cell Biol., 9, 92–98.
30) Abe, K. & Takeichi, M. (2008) EPLIN mediates linkage of the cadherin-catenin complex to f-actin and stabilizes the circumferential actin belt. Proc. Natl. Acad. Sci. USA, 105, 13–19.
31) Taguchi, K., Ishiuchi, T., & Takeichi, M. (2011) Mechanosensitive EPLIN-dependent remodeling of adherens junctions regulates epithelial reshaping. J. Cell Biol., 194, 643–656.
32) Wu, D. (2017) Epithelial protein lost in neoplasm (EPLIN): Beyond a tumor suppressor. Genes Dis., 4, 100–107.
33) Hildebrand, J.D. & Soriano, P. (1999) Shroom, a PDZ domain-containing actin-binding protein, is required for neural tube morphogenesis in mice. Cell, 99, 485–497.
34) Nishimura, T., Takeichi, M., & Lecuit, T. (2009) Remodeling of the adherens junctions during morphogenesis. Tissue Remodeling and Epithelial Morphogenesis, 89, 33–54.
35) Ishiuchi, T. & Takeichi, M. (2011) Willin and Par3 cooperatively regulate epithelial apical constriction through a PKC-mediated rock phosphorylation. Nat. Cell Biol., 13, 860–866.
36) Nishimura, T., Honda, H., & Takeichi, M. (2012) Planar cell polarity links axes of spatial dynamics in neural-tube closure. Cell, 149, 1084–1097.
37) Usui, T., Shima, Y., Shimada, Y., Hirano, S., Burgess, R.W., Schwarz, T.L., Takeichi, M., & Uemura, T. (1999) Flamingo, a seven-pass transmembrane cadherin, regulates planar cell polarity under the control of Frizzled. Cell, 98, 585–595.
38) Yamada, S., Pokutta, S., Drees, F., Weis, W.I., & Nelson, W.J. (2005) Deconstructing the cadherin-catenin-actin complex. Cell, 123, 889–901.
39) Buckley, C.D., Tan, J., Anderson, K.L., Hanein, D., Volkmann, N., Weis, W.I., Nelson, W.J., & Dunn, A.R. (2014) Cell adhesion. The minimal cadherin-catenin complex binds to actin filaments under force. Science, 346, 1254211.
40) Ishiyama, N., Sarpal, R., Wood, M.N., Barrick, S.K., Nishikawa, T., Hayashi, H., Kobb, A.B., Flozak, A.S., Yemelyanov, A., Fernandez-Gonzalez, R., et al. (2018) Force-dependent allostery of the α-catenin actin-binding domain controls adherens junction dynamics and functions. Nat. Commun., 9, 5121.
41) Yonemura, S., Wada, Y., Watanabe, T., Nagafuchi, A., & Shibata, M. (2010) α-catenin as a tension transducer that induces adherens junction development. Nat. Cell Biol., 12, 533–542.
42) Grashoff, C., Hoffman, B.D., Brenner, M.D., Zhou, R., Parsons, M., Yang, M.T., Mclean, M.A., Sligar, S.G., Chen, C.S., Ha, T., et al. (2010) Measuring mechanical tension across vinculin reveals regulation of focal adhesion dynamics. Nature, 466, 263–266.
43) Benham-Pyle, B.W., Pruitt, B.L., & Nelson, W.J. (2015) Cell adhesion. Mechanical strain induces E-cadherin-dependent Yap1 and β-catenin activation to drive cell cycle entry. Science, 348, 1024–1027.
44) Aono, S., Nakagawa, S., Reynolds, A.B., & Takeichi, M. (1999) p120(ctn) acts as an inhibitory regulator of cadherin function in colon carcinoma cells. J. Cell Biol., 145, 551–562.
45) Reynolds, A.B., Roesel, D.J., Kanner, S.B., & Parsons, J.T. (1989) Transformation-specific tyrosine phosphorylation of a novel cellular protein in chicken cells expressing oncogenic variants of the avian cellular src gene. Mol. Cell. Biol., 9, 629–638.
46) Wildenberg, G.A., Dohn, M.R., Carnahan, R.H., Davis, M.A., Lobdell, N.A., Settleman, J., & Reynolds, A.B. (2006) p120-catenin and p190RhoGAP regulate cell–cell adhesion by coordinating antagonism between Rac and Rho. Cell, 127, 1027–1039.
47) Kourtidis, A., Ngok, S.P., & Anastasiadis, P.Z. (2013) p120 catenin: An essential regulator of cadherin stability, adhesion-induced signaling, and cancer progression. Prog. Mol. Biol. Transl. Sci., 116, 409–432.
48) Ichii, T. & Takeichi, M. (2007) p120-catenin regulates microtubule dynamics and cell migration in a cadherin-independent manner. Genes Cells, 12, 827–839.
49) Meng, W., Mushika, Y., Ichii, T., & Takeichi, M. (2008) Anchorage of microtubule minus-ends to adherens junctions regulates epithelial cell-cell contacts. Cell, 135, 948–959.
50) Toya, M. & Takeichi, M. (2016) Organization of non-centrosomal microtubules in epithelial cells. Cell Struct. Funct., 41, 127–135.
51) Nagae, S., Meng, W.X., & Takeichi, M. (2013) Non-centrosomal microtubules regulate F-actin organization through the suppression of GEF-H1 activity. Genes Cells, 18, 387–396.
52) Krendel, M., Zenke, F.T., & Bokoch, G.M. (2002) Nucleotide exchange factor GEF-H1 mediates cross-talk between microtubules and the actin cytoskeleton. Nat. Cell Biol., 4, 294–301.
53) Ito, S., Okuda, S., Abe, M., Fujimoto, M., Onuki, T., Nishimura, T., & Takeichi, M. (2017) Induced cortical tension restores functional junctions in adhesion-defective carcinoma cells. Nat. Commun., 8, 1834.
54) Takeichi, M. (2018) Multiple functions of α-catenin beyond cell adhesion regulation. Curr. Opin. Cell Biol., 54, 24–29.
55) Vassilev, V., Platek, A., Hiver, S., Enomoto, H., & Takeichi, M. (2017) Catenins steer cell migration via stabilization of front-rear polarity. Dev. Cell, 43, 463–479.e5.
56) Dickinson, D.J., Nelson, W.J., & Weis, W.I. (2011) A polarized epithelium organized by β- and α-catenin predates cadherin and metazoan origins. Science, 331, 1336–1339.
57) Schippers, K.J. & Nichols, S.A. (2018) Evidence of signaling and adhesion roles for β-catenin in the sponge ephydatia muelleri. Mol. Biol. Evol., 35, 1407–1421.
58) Nichols, S.A., Roberts, B.W., Richter, D.J., Fairclough, S.R., & King, N. (2012) Origin of metazoan cadherin diversity and the antiquity of the classical cadherin/β-catenin complex. Proc. Natl. Acad. Sci. USA, 109, 13046–13051.
59) Dickinson, D.J., Robinson, D.N., Nelson, W.J., & Weis, W.I. (2012) α-catenin and IQGAP regulate myosin localization to control epithelial tube morphogenesis in Dictyostelium. Dev. Cell, 23, 533–546.
60) Uchida, N., Honjo, Y., Johnson, K., Wheelock, M., & Takeichi, M. (1996) The catenin cadherin adhesion system is localized in synaptic junctions bordering transmitter release zones. J. Cell Biol., 135, 767–779.
61) Yamagata, M., Herman, J.P., & Sanes, J.R. (1995) Lamina-specific expression of adhesion molecules in developing chick optic tectum. J. Neurosci., 15, 4556–4571.
62) Abe, K., Chisaka, O., Van Roy, F., & Takeichi, M. (2004) Stability of dendritic spines and synaptic contacts is controlled by αN-catenin. Nat. Neurosci., 7, 357–363.
63) Togashi, H., Abe, K., Mizoguchi, A., Takaoka, K., Chisaka, O., & Takeichi, M. (2002) Cadherin regulates dendritic spine morphogenesis. Neuron, 35, 77–89.
64) Dean, C. & Dresbach, T. (2006) Neuroligins and neurexins: Linking cell adhesion, synapse formation and cognitive function. Trends Neurosci., 29, 21–29.
65) Suzuki, S.C., Inoue, T., Kimura, Y., Tanaka, T., & Takeichi, M. (1997) Neuronal circuits are subdivided by differential expression of type-II classic cadherins in postnatal mouse brains. Mol. Cell. Neurosci., 9, 433–447.
66) Suzuki, S.C., Furue, H., Koga, K., Jiang, N., Nohmi, M., Shimazaki, Y., Katoh-Fukui, Y., Yokoyama, M., Yoshimura, M., & Takeichi, M. (2007) Cadherin-8 is required for the first relay synapses to receive functional inputs from primary sensory afferents for cold sensation. J. Neurosci., 27, 3466–3476.
67) Uemura, M. & Takeichi, M. (2006) αN-catenin deficiency causes defects in axon migration and nuclear organization in restricted regions of the mouse brain. Dev. Dyn., 235, 2559–2566.
68) Shimada, Y., Usui, T., Yanagawa, S., Takeichi, M., & Uemura, T. (2001) Asymmetric colocalization of Flamingo, a seven-pass transmembrane cadherin, and Dishevelled in planar cell polarization. Curr. Biol., 11, 859–863.
69) Tsukasaki, Y., Miyazaki, N., Matsumoto, A., Nagae, S., Yonemura, S., Tanoue, T., Iwasaki, K., & Takeichi, M. (2014) Giant cadherins Fat and Dachsous self-bend to organize properly spaced intercellular junctions. Proc. Natl. Acad. Sci. USA, 111, 16011–16016.
70) Ishiuchi, T., Misaki, K., Yonemura, S., Takeichi, M., & Tanoue, T. (2009) Mammalian Fat and Dachsous cadherins regulate apical membrane organization in the embryonic cerebral cortex. J. Cell Biol., 185, 959–967.
71) Tanoue, T. & Takeichi, M. (2004) Mammalian Fat1 cadherin regulates actin dynamics and cell-cell contact. J. Cell Biol., 165, 517–528.
72) Hayashi, S., Inoue, Y., Kiyonari, H., Abe, T., Misaki, K., Moriguchi, H., Tanaka, Y., & Takeichi, M. (2014) Protocadherin-17 mediates collective axon extension by recruiting actin regulator complexes to interaxonal contacts. Dev. Cell, 30, 673–687.
73) Nakao, S., Uemura, M., Aoki, E., Suzuki, S., Takeichi, M., & Hirano, S. (2005) Distribution of OL-protocadherin in axon fibers in the developing chick nervous system. Brain Res. Mol. Brain Res., 134, 294–308.
著者紹介Author Profile
 竹市 雅俊(たけいち まさとし)
竹市 雅俊(たけいち まさとし)理化学研究所生命機能科学研究センターチームリーダー.理学博士(京都大学).
略歴1943年名古屋市に生る.66年名古屋大学理学部卒業,69年同大学院理学研究科博士課程退学.70年京都大学理学部生物物理学科助手,78年助教授,86年教授,99年同大学院生命科学研究科教授.2002年理化学研究所発生再生科学総合研究センター・センター長,18年より現職.
研究テーマと抱負動物組織の形成機構を理解する.がん化に伴う細胞行動異常の原因究明.微小管マイナス端結合因子による細胞構造と機能の制御.
趣味生きもの観察,最近はとくに野鳥撮影.