スフィンゴ脂質は,スフィンゴイド塩基を含んだ脂質の総称である.その構造はスフィンゴイド塩基に脂肪酸が結合したセラミドを基本骨格とする.スフィンゴ脂質は,生体膜の10%程度しか占めない比較的マイナーな膜脂質であるが,コレステロールとともに微小領域(マイクロドメイン)の構成分子として機能する.この脂質マイクロドメインは機能タンパク質を集積し,膜を介するシグナル伝達,細菌やウイルスの感染,細胞内小胞輸送などの過程で重要な役割を果たしている1).
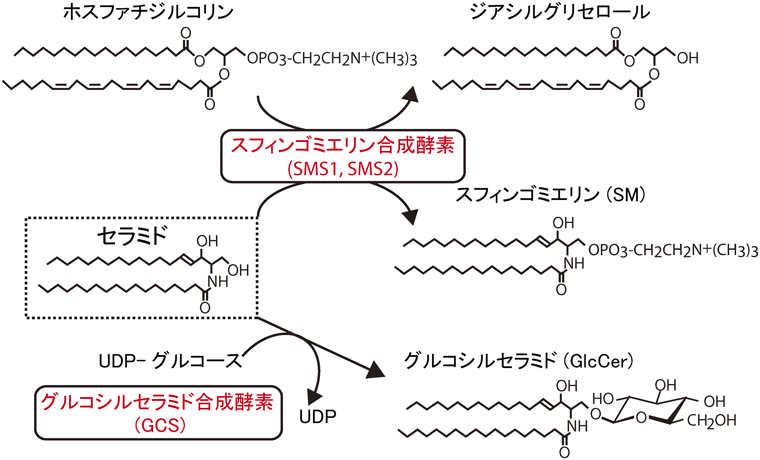
スフィンゴミエリン(SM)は,セラミドにホスホコリンが結合した構造をしており,脂質二重層を形成するスフィンゴ脂質の大部分を占める(図1).SM合成酵素はアミノ酸配列の異なるSMS1およびSMS2の二つのアイソフォームからなり,SMS1はゴルジ体,SMS2はゴルジ体および形質膜に局在する.最近,SM合成酵素が細胞膜上でホモおよびヘテロ複合体を形成することで,脂質マイクロドメインを介したシグナル伝達,SM合成酵素の細胞内局在および安定化,そしてSM合成を調節することがわかってきた.本稿では,SM合成酵素の複合体形成に関する最近の知見を,我々の研究を中心に紹介する.
我々は,SMS2がヒト免疫不全ウイルス(HIV)のエンベロープタンパク質を介した膜融合を促進することをCell-Cell fusion assayを用いて明らかにした2).その解析の過程で,化学架橋剤を用いてSMS2と近接するタンパク質を解析すると,興味深いことにSMS2の二量体と分子量が一致するバンドをウエスタンブロッティングで検出した.さらに,免疫共沈およびBlue-Native PAGEの結果より,SMS2のみならずSMS1も細胞内においてホモ二量体を形成することがわかった.そこで,SM合成酵素のホモ二量体の形成に重要なアミノ酸領域,および,その機能を明らかにすることを目的として以下の実験を行った3).
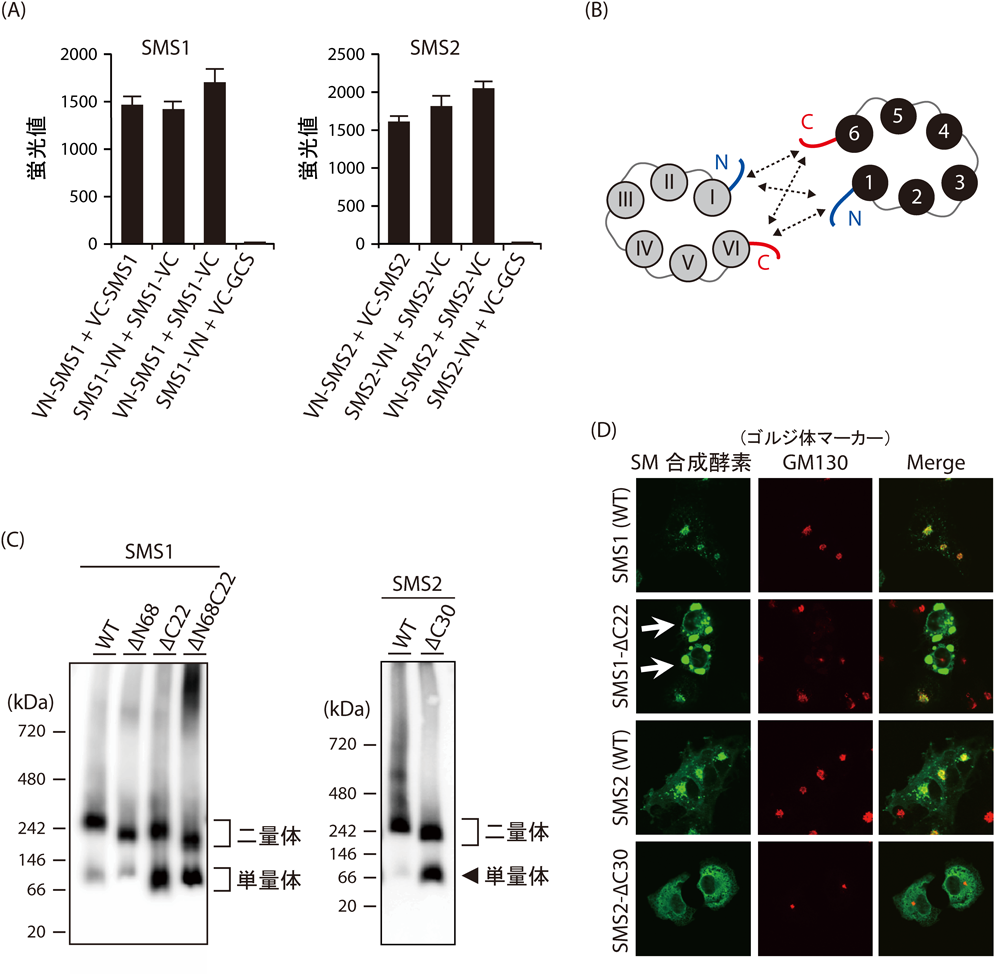
SMS1およびSMS2の各システイン残基をセリン残基に変異したアミノ酸置換変異体を作製し,ホモ二量体形成における近接領域をシステイン残基の化学架橋剤を用いて調べた.その結果,SMS1は50番目のシステイン残基変異体(C50S),SMS2は343, 348番目のシステイン残基変異体(C343S, C348S)において,それぞれの野生型と比較して架橋産物が減少したことから,これらのアミノ酸はホモ二量体を形成する際に近接することが示唆された.N, C末端の配向性の同定から,SMS1のCys50を含むN末端領域,そしてSMS2のCys343, 348を含むC末端領域は,ともにゴルジ体の細胞質側に配向していることがわかった.さらに,SM合成酵素のN, C末端の近接を調べるために,二つの断片に分けた蛍光タンパク質の再構成を指標に分子間相互作用を測定する蛍光タンパク質再構成法を行った.すると,SMS1およびSMS2ホモ二量体においてN末端領域どうし,N末端領域とC末端領域,そしてC末端領域どうしが,それぞれ近接することがわかった(図2A, 2B).
次に,近接領域がホモ二量体の形成に与える影響を調べるために,N, C末端を欠損したSMS1変異体(SMS1-ΔN68, SMS1-ΔC22, SMS1-ΔN68C22)とC末端を欠損したSMS2変異体(SMS2-ΔC30)をCOS-7細胞に発現し,Blue-Native PAGEを行った.その結果,C末端を欠損したSMS1およびSMS2はホモ二量体の割合が減少し,単量体が増加した(図2C).しかしながら,SMS1のN末端欠損体は野生型SMS1と同程度のホモ二量体形成を示し,SMS1のN末端はホモ二量体の形成に関与しないことが示唆された.また,免疫共沈の実験からも同様の結果が得られたことから,SM合成酵素のホモ二量体形成にはC末端を介した相互作用が重要であることがわかった.
3. SMS1およびSMS2のホモ二量体形成はゴルジ体への局在に重要である
SMS1はゴルジ体,SMS2はゴルジ体および形質膜に局在するが,SM合成酵素の二量体が,どこで形成するのかを調べた.細胞をbrefeldin Aで処理しゴルジ体を小胞体へ吸収させ,SMS1およびSMS2のゴルジ体への輸送を阻害してもホモ二量体の形成に影響がなかった.これより,SM合成酵素は小胞体でホモ二量体を形成することが示唆された.続いて,タンパク質のオリゴマー形成は細胞内局在に重要であることが知られているので4),SM合成酵素のホモ二量体形成が細胞内局在に影響するかを調べた.そこで,COS-7細胞にC末端を欠損したSMS1変異体(SMS1-ΔC22)とSMS2変異体(SMS2-ΔC30)を過剰発現させると,野生型と比べてゴルジ体の局在が減少し,小胞体のマーカータンパク質と共局在する割合が増加した(図2D).興味深いことにSMS1のC末端欠損体は,その形態からorganized smooth ER(OSER)を形成していることが考えられた.OSERは小胞体に局在するタンパク質に弱い二量体形成能を持つタンパク質を付加することで観察されることから5),OSER形成はSMS1のC末端欠損による小胞体局在の増加,そしてホモ二量体の形成能の低下に関係することが考えられた.SMS1およびSMS2のC末端欠損体は野生型と同程度の酵素活性を保持することから,C末端欠損体の細胞内局在の変化はタンパク質のミスフォールディングによるものではないことがわかった.また,小胞体に局在する膜タンパク質MBOAT5(LPCAT3)のC末端にSMS1およびSMS2のC末端領域を付加したキメラタンパク質は小胞体に局在したままであることから,SM合成酵素のC末端領域はER exportモチーフを含まないことを確認した.以上の結果より,SMS1およびSMS2のC末端を介したホモ二量体の形成はSM合成酵素のゴルジ体への局在に重要であることが明らかとなった.
最近,PekkinenらによりSMS2が遺伝性骨粗鬆症・骨格異形成の原因遺伝子として同定された6).彼女らは新生児における骨折や骨格の形成異常を特徴とする脊椎骨幹端異形成症という重い症状を示す患者から,一アミノ酸置換のI62SまたはM64Rを引き起こすヘテロ接合性のSMS2変異を同定した.I62SまたはM64R変異体のin vitroでのSM合成酵素活性を測定すると,野生型SMS2と比べて同程度の酵素活性を保持していた.さらに,変異体を保持する患者由来の線維芽細胞を14C-コリンで代謝ラベルすると健常者由来の線維芽細胞と比べてSM合成速度が増加していたが,細胞全体の脂質量ではSM量には顕著な差がなく,トリアシルグリセロール量がわずかに増加していた.興味深いことに,I62SまたはM64R変異体をHeLa細胞に過剰発現させると,本来の局在場所であるゴルジ体や形質膜には局在せずに小胞体に局在した.なぜI62SまたはM64R変異体が小胞体に局在するのかは未解明であるが(PekkinenらはI62とM64がER exportモチーフの可能性を示唆している),SMS2ホモ二量体の形成能の低下の可能性も含めて興味が持たれる.さらに,SMS2が小胞体に局在することが,なぜ骨格形成異常を引き起こすのか,その発症機序の解明が望まれる.
SMとスフィンゴ糖脂質は,共通の基質であるセラミドにホスホコリンか糖のいずれかが結合することによってゴルジ体で産生される(図1).400種以上に及ぶ多種多様な糖鎖構造を示す脊椎動物のスフィンゴ糖脂質の大部分は,セラミドにD-グルコースがβ結合したグルコシルセラミド(GlcCer)を基本骨格とする.哺乳類のGlcCer合成に関わる酵素は現在までにGlcCer合成酵素(GCS)の1種しか見つかっていない.つまり,SM合成酵素とGCSはゴルジ体におけるセラミド代謝の鍵酵素といえる.SMとスフィンゴ糖脂質が形成する脂質マイクロドメインは空間および機能的にも異なることが考えられており7),ゴルジ体でセラミドがSMまたはGlcCerのどちらに合成されるかは,脂質マイクロドメインを介した細胞間・細胞内のさまざまなイベントに影響すると思われる.
前述のように,我々はSM合成酵素のホモ二量体の解析で,蛍光タンパク質再構成法を用いてアミノ末端の近接を調べた.その解析過程で異なるゴルジ槽に局在すると考えられているSMS1とGCSの組合わせをネガティブコントロールとして用いたが,予想に反してゴルジ体で蛍光タンパク質の再構成が観察され,SMS1とGCSは近接することが示唆された.Halterらはタグを付加したSMS1とGCSを安定発現するHeLa細胞において電子顕微鏡で細胞内局在を調べたところ,二つのタンパク質はゴルジ体で実質的には異なる分布を示すが部分的に共局在することを報告している8).しかしながら,ゴルジ体で共局在するSMS1とGCSの役割については不明な点が多い.そこで,SMS1とGCSのヘテロ複合体の形成に重要なアミノ酸領域,および,その機能を明らかにすることを目的として以下の実験を行った9).
免疫共沈の結果,GCSはSMS2よりもSMS1と親和性が高いことがわかった.また,SMS1がSMS1(ホモ二量体)とGCS(ヘテロ複合体)のどちらに親和性が高いのかを免疫共沈で比較したところ,SMS1のGCSに対する親和性はSMS1に対する親和性よりも5倍程度弱かった.しかしながら,ゴルジ体に局在する膜タンパク質であるGOSR1はSMS1やGCSと共沈しなかったことを考えると,親和力は弱いながらもSMS1とGCSはヘテロ複合体を形成することが示唆された.
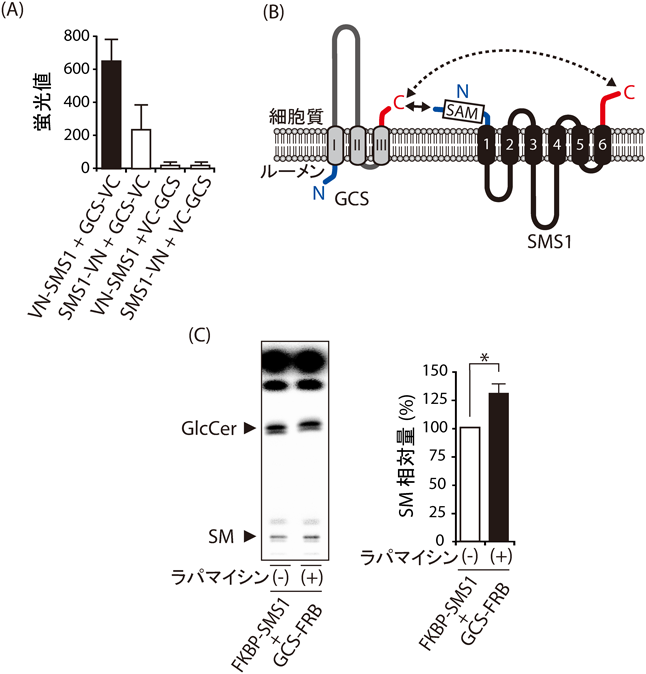
次に,蛍光タンパク質再構成法を用いて,SMS1とGCSのアミノ末端の近接を調べた(図3A).その結果,SMS1-GCSヘテロ複合体において,SMS1のN末端とGCSのC末端は最も近接することがわかった.N, C末端の配向を調べると,SMS1のN, C末端はゴルジ体において細胞質に配向するが,GCSはN末端がゴルジ体のルーメン側,C末端が細胞質側に配向することがわかり,蛍光タンパク再構成法の結果と矛盾しなかった.これらの結果より,SMS1のN末端とGCSのC末端はゴルジ体の細胞質側で近接し,ヘテロ複合体を形成することがわかった(図3B).
SMS1のN末端(4~68 a.a.)には,sterile alpha motif(SAM)領域が存在する.SAM領域はタンパク質相互作用を媒介すると考えられている.ちなみに,SMS1のホモログであるSMSr(SM合成活性はなく,セラミドホスホエタノールアミン合成活性を持つ小胞体に局在するタンパク質)もN末端にSAM領域を持ち,ホモ二量体の形成および細胞内局在に重要であることが知られている10).しかしながら,我々の実験では,SMS1のSAM領域を欠如したSMS1-ΔN68はホモ二量体形成および細胞内局在に変化はなく,SMS1のSAM領域の機能はわかっていなかった.そこで,SMS1のN末端はGCSと近接することから,SMS1のSAM領域がSMS1-GCSヘテロ複合体の形成に関与するのかを免疫共沈で調べた.興味深いことに,SMS1-ΔN68は野生型SMS1と比較してGCSとの親和性が約50%低下した.以上より,SMS1はN末端のSAM領域がGCSとヘテロ複合体の形成に重要であることが示唆された.
5. SMS1-GCSヘテロ複合体はSM合成を正に制御する
次に,SMS1とGCSのヘテロ複合体の形成が,SM合成に与える影響を調べた.内在性のSMS1, GCSによる影響を防ぐために,CRISPR/Cas9システムを用いてHEK293T細胞よりSMS1-GCSダブルノックアウト(DKO)細胞を作製した.FKBP(FK506-binding protein)-SMS1キメラタンパク質とFRB(FKBP-rapamycin binding protein)-GCSキメラタンパク質を発現するSMS1-GCS DKO細胞に対し,ラパマイシン処理でヘテロ複合体の形成を誘導すると,ラパマイシン未処理と比べて,14C-ステアリン酸代謝ラベルによるSM合成速度が約30%増加した(図3C).また,グリシンとセリンの繰り返し配列であるGSリンカーでつなげたSMS1とGCSのキメラタンパク質でも,野生型SMS1よりもSM合成速度が増加したことから,SMS1-GCS複合体はSM合成を正に制御することが明らかになった.
小胞体で合成されたセラミドは,セラミド輸送タンパク質(CERT)によりトランスゴルジ槽に運ばれ,SMへと合成される11).SMS1とGCSは部分的にトランスゴルジ槽で共局在するにもかかわらず,なぜCERTの運ぶセラミドはGlcCer合成に用いられずSM合成に使われるのかは謎である.一つ目の説として,細胞内のSMS1発現量がGCS発現量よりも著しく多いことが考えられる.我々の実験で,SMS1とGCSの間にself-cleaving 2Aペプチド配列を挿入し,等量のSMS1とGCSを細胞内で発現させると,セラミドはSM合成よりもGlcCer合成に多く使われた.しかしながら,実際の哺乳動物細胞ではGlcCerよりもSMの合成に多くのセラミドが使われることを考えると,内在性レベルのGCS発現量はSMS1よりも顕著に少ないことが予想される.つまり,細胞内のSMS1量がGCS量よりも著しく多いため,CERTが運ぶセラミドはSM合成に使われるのかもしれない.
二つ目の説として,IshibashiとHirabayashiらは別の視点からCERTの運ぶセラミドの謎をとらえている12).CERTは,主にトランスゴルジ槽に分布するリン酸化イノシトール脂質であるホスファチジルイノシトール4-リン酸(PI4P)に特異的に結合することでトランスゴルジ槽にセラミドを輸送する11).興味深いことに,PI4Pは糖供与体であるUDP-グルコースのGCSへの取り込みを阻害することで,GlcCer合成を抑制することが明らかになった.CERTがトランスゴルジ槽へ運ぶセラミドの近辺にはPI4Pが豊富に存在することが予想され,このためCERTの運ぶセラミドはGlcCer合成には使われないという説である.CERTとSMS1は部分的にゴルジ体で共局在することから13),SMS1はPI4Pが豊富な領域に局在していることが考えられる.この説の場合,SMS1がGCSとヘテロ複合体を形成することで,PI4Pが豊富な領域にGCSをトラップする可能性があり,SMS1-GCSヘテロ複合体がPI4PによるGlcCer合成の抑制に,どの程度関与しているかに興味が持たれる.
三つ目の説として,SMS1-GCSヘテロ複合体によるセラミド代謝の制御機構がある.しかしながら,なぜSMS1とGCSが複合体を形成することがSM合成を促進するのか不明であり,今後の課題である.CERTが運ぶセラミドの謎の解明には,内在性のSMS1, GCSの細胞内での発現量,そして,その局在を正確に把握する必要がある.
SMS2はSMS1と異なり,ゴルジ体のみならず形質膜に局在する.ちなみに,SMS1とSMS2の局在の違いを規定している要因がパルミトイル化の有無であり,パルミトイル化がSMS2の形質膜への局在に重要であることをTaniらが明らかにしている14).SMS2は形質膜で脂質マイクロドメインに分画することから,細胞間のシグナル伝達に関わるタンパク質と相互作用することが報告されている15).Mitsutakeらは,SMS2ノックアウトマウスでは高脂肪食を摂取しても,野生型マウスと比較して,肥満,インスリン抵抗性,脂肪肝のいずれもが有意に抑えられるのを見いだした15).その解析過程でSMS2が,カベオラの構成タンパク質であるカベオリン-1,および脂肪酸の取り込みに関与するスカベンジャー受容体CD36/FATと相互作用することを免疫共沈で明らかにした.そして,SMS2-CD36/FATヘテロ複合体はCD36/FATの機能を正に制御し,脂肪酸の取り込みを促進することで脂肪肝の形成に関わることを明らかにした.
さらに,KimとHirabayashiらはSMS2およびGPRC5Bノックアウトマウスの表現型の類似から,SMS2とGPRC5Bの相互作用を推測し,パルミチン酸の刺激に応じてSMS2とGPRC5Bがヘテロ複合体の形成を促進することを見いだした16).さらに,GPRC5BとSMS2が相互作用すると,リン酸化酵素FynによりSMS2のリン酸化が促進されることで,SMS2のユビキチン化を阻害し,結果としてSMS2の細胞内における安定性につながることがわかった.そして,SMS2の安定化は細胞内でのジアシルグリセロール量を増加し,インスリンの作用が阻害されること明らかにした.これらの結果は,SMS2はヘテロ複合体を形成することで,脂質マイクロドメイン上でのシグナル伝達やタンパク質の安定化など多様な機能を発揮することを示している.
SM合成酵素の機能は,セラミドを代謝し,SMとジアシルグリセロールの生成であることは間違いない.しかしながら,脂質代謝酵素として機能するだけにとどまらず,さまざまなパートナーと複合体を形成することで多彩な機能を発揮するという酵素を超えた役割があると我々は考え始めている.スフィンゴ脂質の代謝に関与する酵素のクローニングはほとんど終了したが,個々のスフィンゴ脂質代謝酵素の制御に関してはいまだ多くの不明な点が存在する.今後は,内在性レベルのSM合成酵素の発現量を考慮し,SM合成酵素の機能を制御する複合体形成をさらに解析していきたい.
謝辞Acknowledgments
本稿で紹介した研究成果は,和田郁夫教授,荒井斉祐助教(福島県立医科大学医学部),光武進准教授(佐賀大学農学部),谷川尚准教授(城西大学薬学部)との共同研究であることを記し,深く御礼を申し上げます.そして,本研究が行われました帝京大学薬学部の和久敬蔵名誉教授,山下純教授,佐々木洋子講師,そして,いつも叱咤激励してくださる若手(?)教員の仲間たちに感謝いたします.
引用文献References
1) Rietveld, A. & Simons, K. (1998) The differential miscibility of lipids as the basis for the formation of functional membrane rafts. Biochim. Biophys. Acta, 1376, 467–479.
2) Hayashi, Y., Nemoto-Sasaki, Y., Tanikawa, T., Oka, S., Tsuchiya, K., Zama, K., Mitsutake, S., Sugiura, T., & Yamashita, A. (2014) Sphingomyelin synthase 2, but not sphingomyelin synthase 1, is involved in HIV-1 envelope-mediated membrane fusion. J. Biol. Chem., 289, 30842–30856.
3) Hayashi, Y., Nemoto-Sasaki, Y., Matsumoto, N., Tanikawa, T., Oka, S., Tanaka, Y., Arai, S., Wada, I., Sugiura, T., & Yamashita, A. (2017) Carboxyl-terminal tail-mediated homodimerizations of sphingomyelin synthases are responsible for efficient export from the endoplasmic reticulum. J. Biol. Chem., 292, 1122–1141.
4) Martzoukou, O., Karachaliou, M., Yalelis, V., Leung, J., Byrne, B., Amillis, S., & Diallinas, G. (2015) Oligomerization of the UapA purine transporter is critical for ER-Exit, plasma membrane localization and turnover. J. Mol. Biol., 427, 2679–2696.
5) Snapp, E.L., Hegde, R.S., Francolini, M., Lombardo, F., Colombo, S., Pedrazzini, E., Borgese, N., & Lippincott-Schwartz, J. (2003) Formation of stacked ER cisternae by low affinity protein interactions. J. Cell Biol., 163, 257–269.
6) Pekkinen, M., Terhal, P.A., Botto, L.D., Henning, P., Mäkitie, R.E., Roschger, P., Jain, A., Kol, M., Kjellberg, M.A., Paschalis, E.P., et al. (2019) Osteoporosis and skeletal dysplasia caused by pathogenic variants in SGMS2. JCI Insight, 4, 126180.
7) Kiyokawa, E., Baba, T., Otsuka, N., Makino, A., Ohno, S., & Kobayashi, T. (2005) Spatial and functional heterogeneity of sphingolipid-rich membrane domains. J. Biol. Chem., 280, 24072–24084.
8) Halter, D., Neumann, S., van Dijk, S.M., Wolthoorn, J., de Maziere, A.M., Vieira, O.V., Mattjus, P., Klumperman, J., van Meer, G., & Sprong, H. (2007) Pre- and post-Golgi translocation of glucosylceramide in glycosphingolipid synthesis. J. Cell Biol., 179, 101–115.
9) Hayashi, Y., Nemoto-Sasaki, Y., Matsumoto, N., Hama, K., Tanikawa, T., Oka, S., Saeki, T., Kumasaka, T., Koizumi, T., Arai, S., et al. (2018) Complex formation of sphingomyelin synthase 1 with glucosylceramide synthase increases sphingomyelin and decreases glucosylceramide levels. J. Biol. Chem., 293, 17505–17522.
10) Cabukusta, B., Kol, M., Kneller, L., Hilderink, A., Bickert, A., Mina, J.G., Korneev, S., & Holthuis, J.C. (2017) ER residency of the ceramide phosphoethanolamine synthase SMSr relies on homotypic oligomerization mediated by its SAM domain. Sci. Rep., 7, 41290.
11) Hanada, K., Kumagai, K., Yasuda, S., Miura, Y., Kawano, M., Fukasawa, M., & Nishijima, M. (2003) Molecular machinery for non-vesicular trafficking of ceramide. Nature, 426, 803–809.
12) Ihibashi, Y., Ito, M., & Hirabayashi, Y. (2018) Regulation of glucosylceramide synthesis by Golgi-localized phosphoinositide. Biochem. Biophys. Res. Commun., 499, 1011–1018.
13) Spessott, W., Uliana, A., & Maccioni, H.J. (2010) Cog2 null mutant CHO cells show defective sphingomyelin synthesis. J. Biol. Chem., 285, 41472–41482.
14) Tani, M. & Kuge, O. (2009) Sphingomyelin synthase 2 is palmitoylated at the COOH-terminal tail, which is involved in its localization in plasma membranes. Biochem. Biophys. Res. Commun., 381, 328–332.
15) Mitsutake, S., Zama, K., Yokota, H., Yoshida, T., Tanaka, M., Mitsui, M., Ikawa, M., Okabe, M., Tanaka, Y., Yamashita, T., et al. (2011) Dynamic modification of sphingomyelin in lipid microdomains controls development of obesity, fatty liver, and type 2 diabetes. J. Biol. Chem., 286, 28544–28555.
16) Kim, Y.J., Greimel, P., & Hirabayashi, Y. (2018) GPRC5B-mediated sphingomyelin synthase 2 phosphorylation plays a critical role in insulin resistance. iScience, 8, 250–266.
著者紹介Author Profile
 林 康広(はやし やすひろ)
林 康広(はやし やすひろ)帝京大学薬学部助教.博士(農学).
略歴1979年山口県防府市に生る.2001年九州大学農学部卒業.03年同大学院生物資源環境科学府修士課程修了.06年同大学院生物資源環境科学府博士後期課程修了.07年同大学CREST研究員.08年アメリカ国立衛生研究所visiting fellow.10年より現職.
研究テーマ脂質とウィルス感染,脂質代謝酵素の複合体形成.
抱負脂質代謝酵素の機能を制御する新奇分子を探索し,脂質代謝経路の全貌を明らかにしたい.
ウェブサイトhttps://researchmap.jp/1549688131217/
趣味野球.