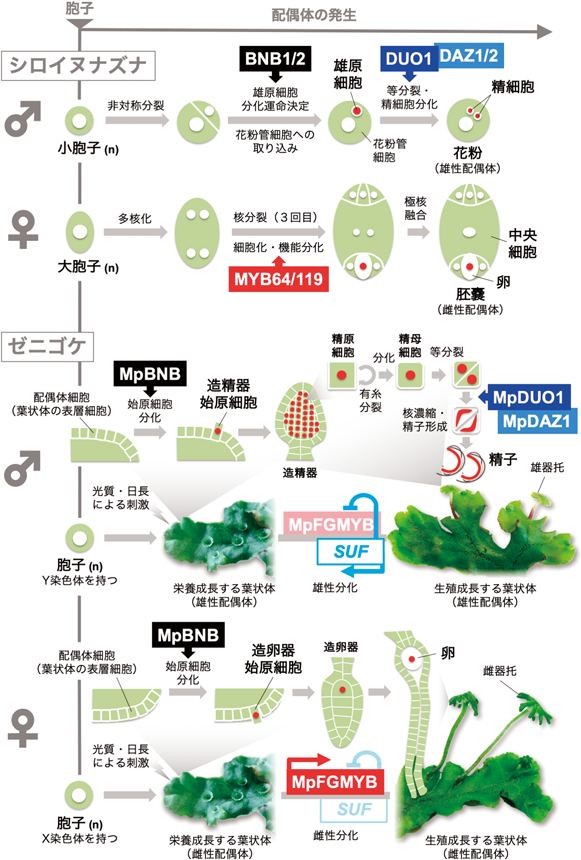
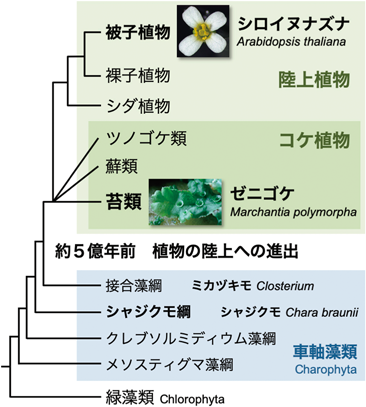
陸上植物の配偶子形成の分子メカニズムとその進化Molecular mechanism and evolution of gametogenesis in land plants
京都大学大学院生命科学研究科Graduate School of Biostudies, Kyoto University ◇ 〒606–8502 京都市左京区北白川追分町 ◇ Kitashirakawa Oiwake-cho, Sakyo-ku, Kyoto 606–8502, Japan
発行日:2019年8月25日Published: August 25, 2019