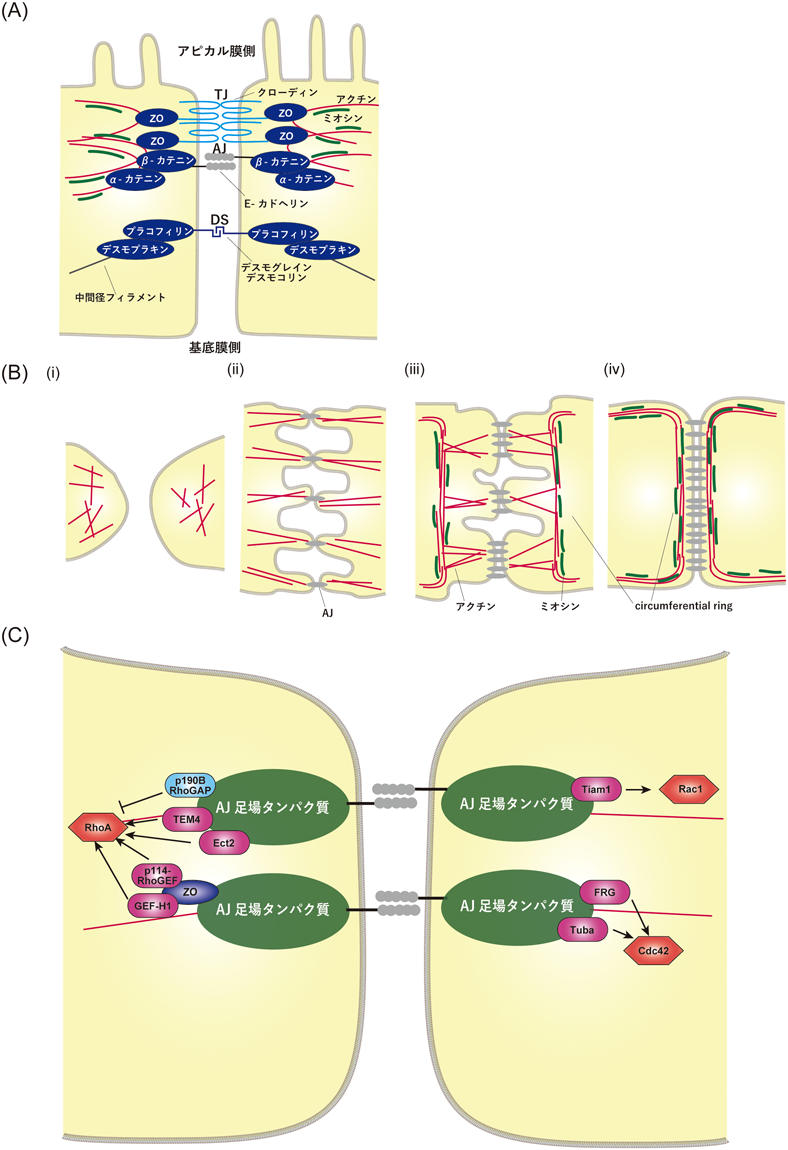
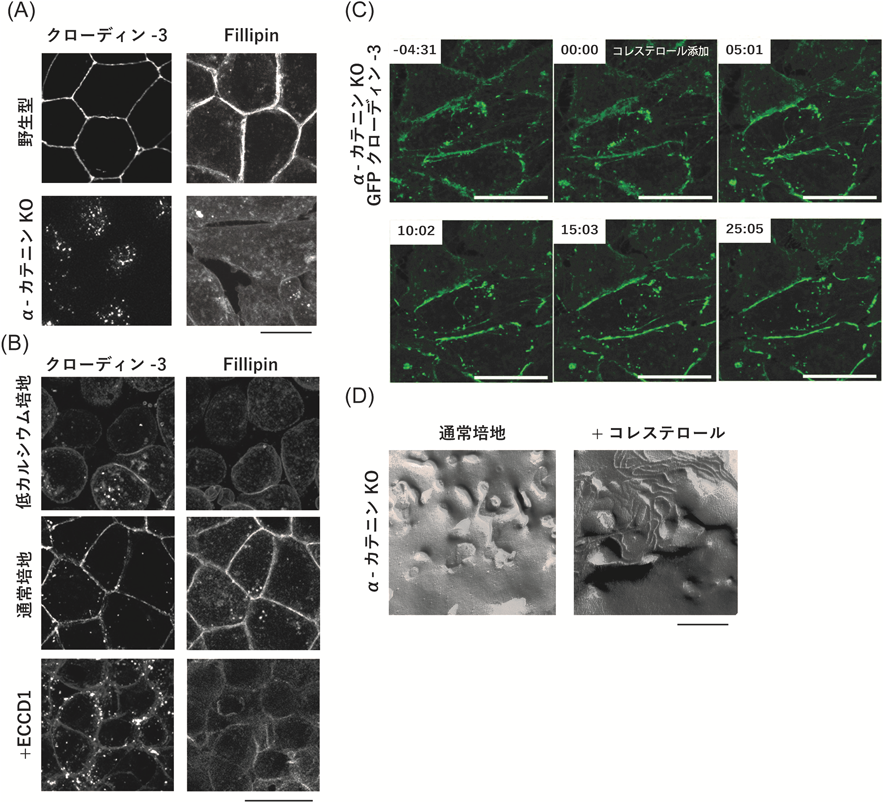
アドへレンスジャンクションを介したタイトジャンクション形成の制御機構How adherens junction formation enables tight junction formation?
九州大学理学研究院Department of Biology, Faculty of Sciences, Kyushu University ◇ 〒819–0395 福岡市西区元岡744 ◇ 774 Moto-oka, Nishi-ku, Fukuoka 819–0395, Japan
発行日:2019年8月25日Published: August 25, 2019