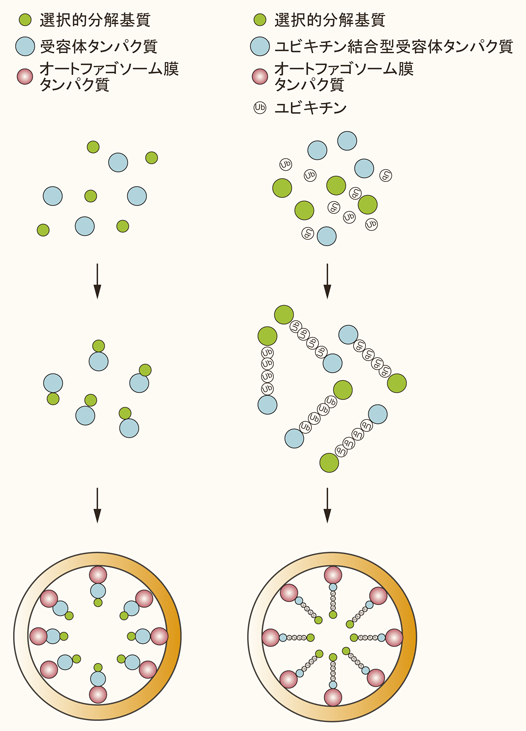
栄養飢餓誘導型のマクロオートファジーでは無作為に細胞質成分を分解すると考えられている一方,ある状況下では特定の積み荷(カーゴ)をオートファゴソームが選択的に認識,隔離,分解する.後者は選択的マクロオートファジーと呼ばれ,特定の可溶性タンパク質,タンパク質凝集体,不要なオルガネラ,病原性細菌を分解することで細胞の恒常性維持に貢献している1).通常のオートファジーと選択的オートファジーにおけるオートファゴソーム膜形成の分子機構は共通であると考えられるが,選択的オートファジーではストレスに応じた「各カーゴの標識」や「受容体タンパク質」により選択性が担保される2).「各カーゴの標識」とは,カーゴのユビキチン化や受容体タンパク質のカーゴへの局在化を意味する.一方,「受容体タンパク質」は,カーゴとオートファゴソーム局在タンパク質LC3ないしはGABARAPファミリーに結合するタンパク質群を指す.受容体タンパク質は,カーゴのユビキチン鎖を認識するユビキチン結合型受容体タンパク質と分解カーゴ上に局在するカーゴ局在型受容体タンパク質の二つに分けられる2)(図1).いずれのタイプの受容体タンパク質もLC3相互作用領域(LC3-interacting region:LIR)あるいはGABARAP相互作用領域(GABARAP-interacting motif:GIM)を有しており,オートファゴソーム膜に局在するLC3ないしはGABARAPファミリーに,あるいは両ファミリーに直接結合する3–7)(図1).LIRの翻訳後修飾による制御も存在する.細胞内侵入細菌やミトコンドリアの受容体タンパク質であるOptineurinやNix/Bnip3Lは,LIRの直前に存在するセリン残基のリン酸化によりLC3との相互作用が増強する8, 9).また,LC3ないしはGABARAPに特異的に結合する受容体タンパク質も同定されており,LC3のホモログに使い分けがあることもわかってきた3–7).しかし,哺乳動物Atg8ホモログをすべて欠損したHeLa細胞においても,脱分極したミトコンドリアをオートファゴソームが隔離する像が観察されている10).このことは,少なくともマイトファジーにおいては受容体タンパク質群とLC3ないしはGABARAPとの相互作用は必須でないことを意味する.受容体タンパク質のいくつかはオートファゴソーム形成に必須な上流因子FIP200との相互作用も知られており11, 12),LC3やGABARAPとの相互作用は補完的な役割を担う可能性も残る.本稿では,マルチモードオートファジーのうちマクロオートファジーによる可溶性タンパク質の選択的分解に焦点をあて,その生理作用について著者らの最近の研究成果を中心に紹介する.
2. p62/SQSTM1分解:液–液相分離,Nrf2,マクロオートファジー
タンパク質やRNA分子の間に働く物理的な力が,分子を互いに引き離したり引き寄せたりする場合がある.分子の濃度が特定の値に達すると,あるいは修飾により結合価が増えると液–液相分離(liquid–liquid phase-separation)が起こり,類似成分が集まって濃縮し,反応の速度を上げたり,望ましくない分子を隔離したりできるようになる.このような構造体は,液体様の性質を備えていることから液滴(liquid droplet)と呼ばれ,その内部は流動性と生化学反応が維持されている可逆的な構造体である13).細胞内にはストレス顆粒,生殖顆粒や核小体など多数の液滴が存在しており,環境の変化といった外乱の影響によって変化することを阻止する内的な仕組み,つまりロバストネスを制御する.多くのタンパク質が分離もしくは凝縮した液滴ではその内容物の濃度が高くなっており,さまざまな生化学的反応が効率よく進んでいる14).したがって,過剰あるいは異常な液滴は速やかに分散あるいは分解される必要がある.液滴はオルガネラに匹敵する大きさであり,しばしばmembrane-less organelles(MLOs)と呼ばれる15).液滴は融合を繰り返す動的な球状構造体であり,その合理的な典型例としてオートファジー経路による分解がある.事実,液滴であるストレス顆粒や生殖顆粒はオートファジーにより分解される(佐藤らの稿)16, 17)一方,オートファジーの不全は液滴がゲルあるいは不溶化したと考えられるユビキチン陽性の凝集体形成を伴った神経変性疾患や肝腫瘍といったさまざまな疾患を起こす18).
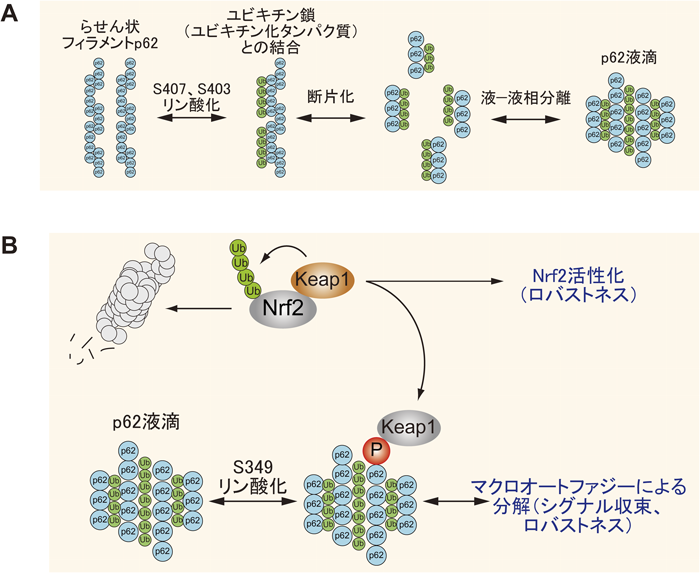
最も代表的なオートファジー選択的分解基質p62/SQSTM1(以降はp62と省略)はユビキチン化タンパク質と結合することで液–液相分離を起こす19, 20)(図2A).通常状態では,p62は,UBAドメインどうしの会合によりユビキチン鎖と相互作用ができない21, 22).また,N末端のPB1ドメインに存在する7番目のリシン残基と69番目のアスパラギン酸の間の静電的相互作用によるhead-to-tailの自己相互作用によりらせん状フィラメントを形成している23)(図2A).プロテオスターシス(タンパク質恒常性維持機構)の異常などに応じてp62のUBAドメイン内の407番目のセリン残基がULK1によりリン酸化されると,UBAドメインどうしの会合が解除されユビキチン鎖と結合可能となる24).その後,ULK1,カゼインキナーゼ2またはTANK結合キナーゼ1により403番目のセリン残基がリン酸化されると,p62とユビキチン鎖との結合が増強する24–27)(図2A).ユビキチン鎖の結合によりp62のらせん状フィラメントは断片化し,液体様の性質を維持したまま細胞質内に濃縮(相分離)する19, 20)(図2A).最終的に,相分離したp62の境界面に沿って隔離膜が形成され,LC3との相互作用依存的にオートファジーにより分解されると考えられる.このLC3との結合に先立って,p62はLIR近傍のセリン残基のリン酸化依存的にFIP200と結合するらしい11).この結合はLC3の結合と相互排他的であることから,FIP200の結合が解除されてからLC3への結合が起こると考えられる.興味深いことに,p62と同様のドメイン構造を持つ受容体タンパク質NBR1の存在下でp62の相分離が促進される.おそらく,p62フィラメント断片が相分離する過程においてNBR1はp62に作用すると考えられる20).
著者らは,プロテオスターシスの異常など選択的マクロオートファジー誘導時にp62の349番目のセリン残基(S349)がリン酸化されるとp62とCullin 3型ユビキチンリガーゼのアダプタータンパク質Keap1とが直接に相互作用すること,その結果,Keap1の標的である酸化ストレス応答のマスター転写因子であるNrf2が安定化して核内に移動し,抗酸化タンパク質の遺伝子発現を統一的に誘導することを見いだした28–31)(図2B).これはさまざまな内乱・外乱にもかかわらず機能を維持できる内的な仕組み,つまり細胞のロバストネスの一種といえる.p62とKeap1複合体はオートファジーカーゴとともにマクロオートファジー依存的に分解され,Nrf2活性化のシグナルをシャットダウンすると考えられる32, 33)(図2B).
p62は最も代表的なマクロオートファジー選択的分解基質であり,マクロオートファジーの障害により著しく蓄積,凝集化するとともにNrf2を活性化する.重要なことに,p62タンパク質は神経変性疾患やアルコール性肝炎,脂肪肝や肝細胞がんなどの肝疾患で確認される封入体の主要構成成分でもある34).しかし,その病態生理的意義はまったく不明であった.著者らは,肝細胞がんで確認されるマロリー小体あるいは硝子体と呼ばれるp62陽性構造体35)に注目し,肝細胞がんにおけるp62介在性Nrf2活性化の意義を調べた.S349がリン酸化されたp62を過剰に蓄積し,p62陽性構造体を有する肝細胞がん株Huh-1では恒常的にNrf2が活性化していた29, 36, 37).一方,ほとんどS349リン酸化p62を持たない別の肝細胞がん株Huh-7, JHH-1, HepG2ではNrf2の活性は低く保たれていた.Huh-7, JHH-1, HepG2にS349リン酸化模倣変異体p62(p62 S349E)を発現させるとNrf2の活性化が確認された一方,そのような効果はHuh-1ではみられなかった37).近年,肺がんや食道がんなど固形腫瘍の症例においてNrf2が恒常的に活性化していることが報告され,それが予後不良因子であることも明らかになった38).恒常的に活性化したNrf2は,解毒系酵素群や抗酸化タンパク質をコードする遺伝子発現を強力に誘導することで酸化ストレスや抗がん剤や放射線治療に対する抵抗性を増強する38).さらに,がん細胞においてグルコースやグルタミンの代謝を変化させて同化反応を促進することによりがん細胞の増殖に貢献する39).野生型p62, p62 S349Eあるいはリン酸化欠失変異体p62(p62 S349A)を発現させたHuh-7のメタボローム解析から,S349Eの発現によりUDP–グルコース,UDP–グルクロン酸,還元型グルタチオンが増加することがわかった37).13C6グルコースを用いたトレーサー解析からp62 S349Eの発現により13C6でラベルされたUDP–グルクロン酸の産生が,13C5グルタミン酸を用いたトレーサー解析からp62 S349Eの発現により13C5でラベルされた還元型グルタチオンの産生が,それぞれ上昇することがわかった37).このことはS349リン酸化p62を持つ肝細胞がんは,グルタチオン合成の促進により抗酸化能を,グルクロン酸抱合およびグルタチオン抱合能の上昇により薬剤耐性を獲得していることを示唆する.事実,Huh-7にp62 S349Eを過剰発現させるだけでソラフェニブやシスプラチンといった抗がん剤に耐性を示すようになった.一方,元来S349リン酸化p62を過剰に持つHuh-1細胞ではそのような効果は確認されなかった37).
3. NCoR1分解:細胞質移行,オートファジー,細胞内代謝
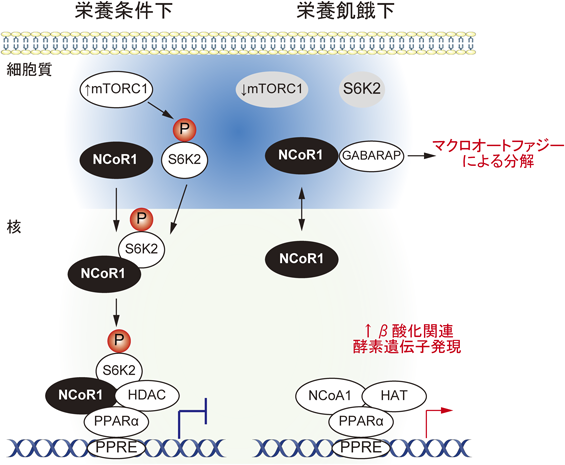
NCoR1(nuclear receptor co-repressor 1)は,核内受容体とヒストン脱アセチル化酵素と相互作用し,ヒストンを脱アセチル化することで核内受容体PPARαなどの転写活性を抑制する40)(図3).栄養条件下では,NCoR1はmTORC1によりリン酸化されたS6K2と複合体を形成して,核内に移行し,PPARαをはじめとした核内受容体の活性化を抑制している41).そのため,PPARαの標的遺伝子であるβ酸化関連酵素をコードする遺伝子群の発現は低い.このとき,mTORC1によりオートファジーの上流因子ULK1もリン酸化され,オートファジーも抑制される42, 43).栄養飢餓などの刺激によりmTORC1が不活性化すると,S6K2の脱リン酸化が起こりNCoR1は核外に移行する(図3).その結果,NCoR1とNCoA1(nuclear receptor co-activator 1)が入れ替わり,NCoA1に結合するヒストンアセチル化酵素により核内受容体の標的遺伝子のエンハンサー領域のアセチル化が促進され,標的遺伝子の発現が誘導される40, 44)(Korzus et al., 1998;Perissi et al., 2010)(図3).著者らは,NCoR1はGABARAPとの結合に必須なGIMを持ち,GABARAPとの結合依存的にマクロオートファジーにより分解されることを見いだした6).オートファジー関連遺伝子ATG7を欠失させた肝がん細胞株HepG2は,核および細胞質においてNCoR1を過剰蓄積し,PPARαの標的遺伝子であるCPT1AやCPT2のエンハンサー領域のヒストン脱アセチル化が亢進,栄養飢餓に応じたPPARαの標的遺伝子群の発現誘導が顕著に抑制されていた6).このことは,飢餓に応じたNCoR1の核外移行の制御だけでは不十分であり,マクロオートファジーによるNCoR1の分解がPPARαの効率のよい活性化に必須なことを意味する.肝臓特異的Atg7あるいはAtg5欠損マウス肝臓を用いたリピドーム解析から,オートファジーの抑制が長鎖アシルカルニチンの蓄積を伴うことがわかった6).さらに,安定同位体の13Cでラベルされたパルミチン酸を取り込ませたマウスのメタボローム解析から,コントロールマウス肝臓に比し肝臓特異的Atg5欠損マウス肝臓では飢餓に応じた13Cラベルされたβ-ヒドロキシ酪酸(ケトン体の一種)がほとんど産生されないことがわかった6).これらのことは,マクロオートファジー欠損肝臓では,飢餓に応じたβ酸化が抑制されていることを意味する.ほぼ同様の結果がマクロオートファジー抑制を伴う肝特異的Vps15欠損マウスでも観察された45).以上をまとめると,マクロオートファジーは栄養飢餓に応じてグリコゲノリシスやリポリシスなど他の経路とともに栄養素を供給するだけでなく,NCoR1の分解を促進することでPPARα標的遺伝子のエンハンサー領域のヒストンのアセチル化を亢進させ,β酸化関連酵素をコードする遺伝子群の発現を誘導する.つまり,マクロオートファジーはβ酸化のための材料である脂肪酸を供給するとともに,それを利用する経路を同時に活性化すると考えられる.
本稿では可溶性タンパク質のマクロオートファジーによる選択的分解の分子機構と生理作用について著者らの結果を中心に概説した.可溶性タンパク質のマクロオートファジー分解は,1分子の基質と結合した受容体タンパク質が,隔離膜に局在するLC3やGABARAPとの相互作用を介して選択的あるいは優先的にオートファゴソームに取り込まれると想定される.たとえば,転写因子CRY1や核内受容体コリプレッサーNCoR1などがこれにあたる6, 7).しかし,オートファゴソームは平均的な大きさのタンパク質であれば10万のタンパク質を一度に取り込めることを考慮すると,この分解系はきわめて効率が悪い.おそらく多くのマクロオートファジーにより選択的に分解される可溶性タンパク質はp62のように液–液相分離するか,液–液相分離するタンパク質のクライアントタンパク質(たとえばp62の場合のクライアントタンパク質NBR1)なのであろう.今後,オートファジーにより選択的に分解される可溶性タンパク質の同定,およびその液–液相分離能,もし液–液相分離するのであればその生理機能の解明が重要であろう.