オートファジーの構造生物学第二章Structural biology of autophagy, Chapter II
公益財団法人微生物化学研究会微生物化学研究所Institute of Microbial Chemistry ◇ 〒141–0021 東京都品川区上大崎3–14–23 ◇ 3–14–23 Kamiosaki, Shinagawa-ku, Tokyo 141–0021, Japan
発行日:2019年10月25日Published: October 25, 2019
オートファジーの最大の特徴は二重膜オルガネラであるオートファゴソームの新生を伴う点であり,その過程には6種類のAtg機能グループが関与している.これらの構造生物学研究は,まず2種類,すなわちユビキチン様のAtg8およびAtg12結合反応系について進展し,筆者は2013年に本誌の総説で紹介した.その後の5年間は研究が他の機能グループへと急速に発展し,特にオートファジーの始動を担うAtg1複合体や,オートファゴソーム膜の伸長に直接関与するAtg2の構造研究の進展により,研究はいよいよオートファゴソーム形成機構の核心に迫るフェーズに突入した.本稿では,“オートファジーの構造生物学第二章”と題して,最近5年間のオートファジーの構造生物学研究の進展をまとめるとともに,そこから提起されたオートファゴソーム形成の分子機構を紹介する.
© 2019 公益社団法人日本生化学会© 2019 The Japanese Biochemical Society
マクロオートファジー(以降オートファジー)最大の特徴は,二重膜オルガネラであるオートファゴソームの新生を伴う点である.通常のオルガネラ膜はその機能のために多くの膜タンパク質を含んでいるが,オートファゴソーム膜,とりわけその内膜はほとんど膜タンパク質を含まないことが知られており1),既存オルガネラ膜とは異なる膜が新生されていると考えられる.出芽酵母における遺伝学的解析により,栄養飢餓で誘導されるオートファゴソームの形成には18種類のオートファジー関連(Atg)タンパク質(主要Atgタンパク質)が必須であること,その後の細胞生物学的・生化学的解析により,これらタンパク質群は六つの機能グループに分かれて機能することが明らかとなった(本特集・中戸川の稿)2).しかしながら,主要Atgタンパク質群がオートファゴソーム形成過程において具体的にどのような分子機能を担うのか,そしてその結果どのようにオートファゴソームが新生されるのか,まったくといっていいほどわからない状態がAtgタンパク質の発見以来,20年以上も続いてきた.タンパク質の機能を知る上で,その立体構造はしばし大きな手がかりを与えてくれる.筆者らは2002年から主要Atgタンパク質群の構造生物学研究に着手し,その後多くの構造生物学者が参入して互いに切磋琢磨しながら研究を展開し,2013年までには主要Atgタンパク質のうち約半数を占めるAtg結合反応系について,構造基盤の全容の解明に成功した3, 4).そして筆者は当時の知見をまとめて本誌に「オートファジーの構造生物学」という題の総説を執筆した5).これら構造情報はAtg結合・脱結合反応のメカニズムやオートファジーの選択性発現メカニズムの解明には大いに貢献したが,その一方で核心テーマであるオートファゴソーム形成のメカニズム解明に関してはほとんど無力であった.しかしながら,それから現在に至る約5年間で,オートファジーの始動を担うAtg1複合体(オートファジー始動複合体と呼ばれる)およびオートファゴソームの膜形成で鍵を握ると考えられてきた機能未知タンパク質Atg2について,構造生物学研究が急速に進展し,ようやくオートファゴソーム形成機構の解明につながる知見が得られる段階にまで到達した.本稿では,「オートファジーの構造生物学第二章」と題して,これら最新の構造生物学研究の成果と,そこからわかってきたオートファジーの分子機構について紹介する.
主要Atgタンパク質のうち,最上流で機能するのがオートファジー始動複合体であり,出芽酵母の場合Atg1, Atg13, Atg17, Atg29, Atg31の五つの因子からなる6).栄養飢餓によりオートファジーが誘導されると,液胞近傍の1点にほぼすべての種類の主要Atgタンパク質が集積し,プレオートファゴソーム構造体(pre-autophagosomal structure:PAS)を形成する7).PASへの局在の仕方には階層性があり,最初にPAS局在するのが(あるいはPASを最初に構成するのが)オートファジー始動複合体である8).オートファジー始動複合体がPASの核となり,続いて隔離膜の最初の膜源と考えられているAtg9小胞9)をリクルートし,さらに下流のAtg因子群をリクルートすることでPASが完成し,オートファゴソーム形成が開始すると考えられる(図1).PASの核としての機能以外に,オートファジー始動複合体はAtg1が持つキナーゼ活性を用いて下流Atg因子群をリン酸化する役割も担っており,それもオートファジーの進行に重要である6).
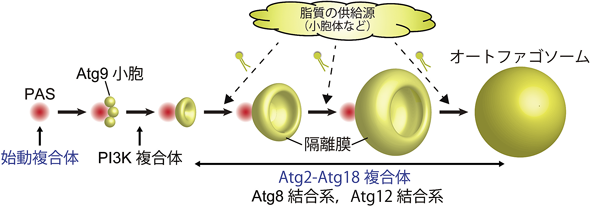
栄養飢餓でオートファジーが誘導されると,始動複合体がPASの核を構築し,続いて膜の最初の材料であるAtg9小胞,ホスファチジルイノシトール3-キナーゼ(PI3K)複合体,そしてさらに下流のAtg2-Atg18複合体やAtg結合系が順次PASへと局在することで,隔離膜の形成が開始する.隔離膜の材料となる脂質は小胞体などの供給源から何らかのメカニズムで隔離膜へと移行する必要がある.
オートファジー始動複合体の構造的特徴の一つは天然変性領域に富むことであり,Atg1, Atg13, Atg29, Atg31の4因子は配列から天然変性と予測される残基の割合が約3~7割あり,特にAtg13は約7割の残基が天然変性と予測される.一方,Atg17は唯一天然変性領域をほぼ持たないタンパク質である.構造生物学的研究は,天然変性領域を極力除いたドメイン単位,サブ複合体単位で進められてきた(図2).
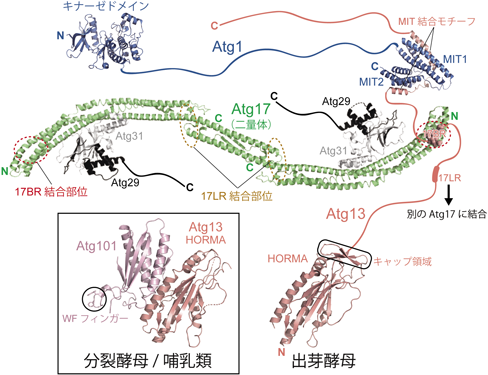
Atg1, Atg13, Atg29の天然変性領域はひもで示してある.リボン図はPyMOLで作成し,以下のPDBを用いた.Atg1のキナーゼドメイン:4WNO, MITドメイン:4P1N, Atg13のHORMA:4J2G, Atg17-Atg29-Atg31複合体:5JHF, Atg101-Atg13複合体:4YK8.
Atg1はN末端側のキナーゼドメインおよびC末端側のタンデムMITドメインと,それらをつなぐ天然変性領域からなる6).MITドメインは一般にタンパク質-タンパク質間の相互作用を担うことで知られており,ヘリックス1本からなるMIT結合モチーフと結合するが10),Atg1の二つのMITも,それぞれがAtg13の天然変性領域に存在するヘリックスと結合する11).Atg13はN末端にHORMAドメインを持ち12),それ以外はほぼ天然変性領域と予測されている.実際に高速原子間力顕微鏡でAtg13を観察すると,球状のHORMAドメイン以外はひも状構造として観察される13).Atg13の天然変性領域にはAtg1のMITとの結合を担う二つのMIT結合モチーフと,Atg17との結合を担う二つの短い領域(17BRおよび17LR)が存在する13).Atg17はヘリックス4本からなり,曲がりながら長く伸びた弓状の構造を持ち,それが二量体を形成することで長さ30 nm程度の特徴的なS字構造をとる14).Atg17のN末端はS字構造の両末端に,C末端は二量体形成部位近傍に位置しており,前者にはAtg13のAtg17-binding region(17BR)が結合するポケットが,後者にはAtg17-linking region(17LR)が結合するポケットが存在する11, 13).二つのポケットは互いに離れているが(約13 nm),それが機能に重要な意味を持つ(後述).Atg29およびAtg31は互いにストランドを出し合って一つのβシートを形成する兄弟タンパク質であり,Atg31のC末端にあるヘリックスでAtg17のS字構造の凹面中心付近に結合し,安定なAtg17-Atg29-Atg31複合体を構築する.
上述した部分構造と相互作用情報を元にオートファジー始動複合体モデルを構築すると,図2のようになる.すなわちひも状構造を持ったAtg13がAtg17-Atg29-Atg31複合体とAtg1を架橋するように結合することで五者複合体を構築する.Atg13は2か所のAtg17結合領域を持っているが,Atg17上の二つの結合部位が互いに離れているため,両方同時に結合することが不可能に思われた.それにもかかわらず,Atg13のどちらのAtg17結合領域も,またAtg17のどちらのAtg13結合部位もPAS形成およびオートファジー活性に必須であった13).精製タンパク質を用いてオートファジー始動複合体を調製し,ゲルろ過クロマトグラフィーおよび分析超遠心による解析を行った結果,始動複合体はAtg13の二つのAtg17結合領域依存的に,高次会合体を形成することが明らかとなった13).すなわちAtg13は同じAtg17分子に2か所で結合するのではなく,二つの結合領域を用いて異なる2分子のAtg17を橋渡しするように結合し,その結果始動複合体の高次会合体形成を促進していると考えられる(図3A).in vitroにおける始動複合体の高次会合体形成活性は,酵母におけるPAS形成活性と強い相関を示したことから13),始動複合体の高次会合体形成がPASの構築自体に重要であると考えられる.
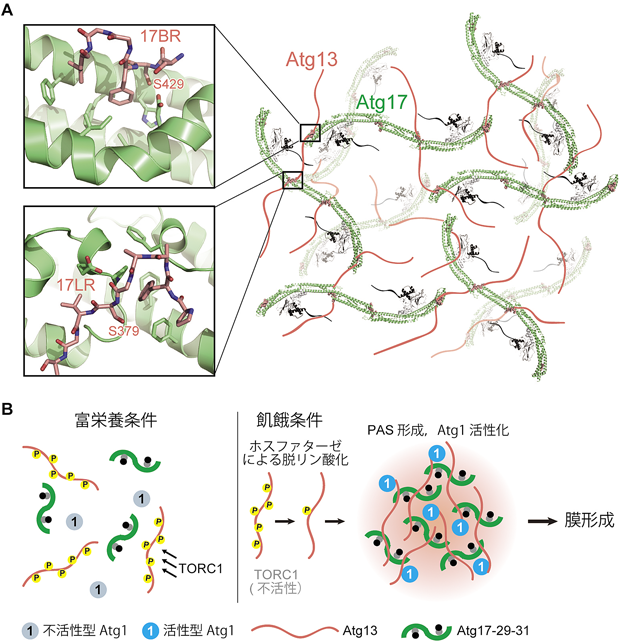
(A) Atg13を介した高次会合体形成モデル.Atg13を赤のひもで示す.図の簡略化のため,始動複合体からAtg1を省いてある.左の差し込み図は上が17BRとAtg17の相互作用,下が17LRとAtg17の相互作用を示す(PDB 5JHF).(B)栄養飢餓によるオートファジー始動モデル.富栄養条件下,Atg13は活性型TORC1により高度なリン酸化を受け,始動複合体を形成できない.Atg1は低活性型の状態で存在する.飢餓条件になるとTORC1の活性が阻害され,ホスファターゼの働きによりAtg13が脱リン酸化する結果,始動複合体の高次会合体形成が進行しPASが形成される.PASではAtg1が活性化され,隔離膜形成が進行する.
オートファジーの始動は栄養センサーであるTORキナーゼ複合体1(TORC1)により制御されている15).TORC1はAtg13を直接リン酸化することでオートファジー始動複合体の形成を阻害し,その結果PASの形成を抑えることでオートファジーを阻害していると考えられてきた.Atg13に関して栄養飢餓依存的に脱リン酸化レベルが低下する残基の網羅的な同定を行った結果,実に50か所を超えるセリン・トレオニン残基が飢餓依存的な脱リン酸化を受けることが明らかとなった13).それらはすべて天然変性領域に存在しており,Atg1結合領域や2か所のAtg17結合領域にも含まれる.Atg13の17BRおよび17LRにはAtg17との結合に直接関わるセリン(429番および379番)が存在しており(図3A,差し込み図),それらをリン酸化模倣変異させるとAtg17との結合が減弱した.セリン429番の変異体に関する詳細な解析の結果,この1残基のリン酸化だけでPAS形成およびオートファジー活性が顕著に阻害を受けることが明らかとなった13).一方,Atg13のAtg1結合領域に関しては,特定の1残基のリン酸化による影響はなく,複数のリン酸化が進むことでAtg1との結合が減弱するようである.
主要Atg因子のほとんどは進化上高度に保存されている.オートファジー始動複合体においてもAtg1およびAtg13は保存されており,Atg17についてもその機能ホモログとしてFIP200が高等生物に存在する6).しかしながら,Atg29およびAtg31の2因子は出芽酵母の近縁種でしか保存されておらず,分裂酵母や哺乳類では代わりにAtg101を構成因子として持つ16).またAtg29-Atg31はAtg17に結合するのに対し,Atg101はAtg17の機能ホモログであるFIP200ではなく,Atg13に結合する.これまでに分裂酵母およびヒトのAtg101に関して,単独あるいはAtg13との複合体として結晶構造が決定された16–20)(図2,差し込み図).
Atg101はAtg13のN末端ドメインと同様,HORMAドメイン構造を持つ.HORMAドメインはMAD2というタンパク質に関して詳細な構造機能解析が行われており,オープン型とクローズ型の2種類の構造をとること,MAD2のオープン型とクローズ型はヘテロ二量体を形成することが知られている21).興味深いことに,Atg101はオープン型,Atg13はクローズ型のHORMA構造を持ち,両者で結合してMAD2と同様のヘテロHORMA二量体を形成する17).出芽酵母のAtg13のHORMAはキャップと呼ばれる挿入領域を持ち,それが構造安定化に寄与しているが,分裂酵母やヒトなどAtg101を保存した種では,Atg13のHORMAはキャップ領域を持たず,単独では不安定な構造を持つ12, 17).Atg101の役割の一つとして,複合体形成を通して不安定なAtg13を安定化する役割があると考えられる.またAtg101のHORMA構造にはトリプトファンとフェニルアラニンを保存した特徴的なループ(WFフィンガー)が挿入されており,それが下流Atg因子のリクルートに関与すること,またC末端領域がオートファジーに必須なホスファチジルイノシトール3-キナーゼ(PI3K)複合体のリクルートに関与することが報告された17, 18).すなわちAtg101は高等生物においてAtg因子群が集積する過程を促進する機能があると考えられる.
以上述べてきた知見に基づくと,酵母における飢餓時のオートファジーの始動機構は以下のようになると考えられる(図3B).富栄養条件下では高活性状態のTORC1がAtg13を直接リン酸化し,始動複合体の形成を阻害している.飢餓になると速やかにTORC1の活性が阻害され,PP2C型ホスファターゼの働きによりAtg13の脱リン酸化が亢進する15, 22).脱リン酸化したAtg13はAtg1およびAtg17との結合が増強され,オートファジー始動複合体を形成し,さらにAtg17を架橋していくことで高次会合体化を促進し,PASの核を形成する13).そして下流因子がPASに順次リクルートされることで,オートファゴソーム形成が進行する8).このモデルでは,始動複合体の高次会合体がPASの実体ということになるが,ではそれはどのような性質を持った会合体であろうか? 上述のとおり,始動複合体の構成因子は天然変性に富むという特徴があり,形成された高次会合体はリボソームのような形のしっかりした超分子複合体ではなく,やわらかい会合体であると予想される.そしてリン酸化によりその形成がダイナミックに制御されており,特定のタンパク質(Atg因子)を濃縮する場として機能する.これらの特徴は,近年注目されている,液-液相分離により形成されるメンブレンレスオルガネラに酷似している.実際に,我々はPASが液体の性質を持つこと,始動複合体がin vitroにおいて液-液相分離することを確認しており(論文投稿中),PASはメンブレンレスオルガネラの一種であることが強く示唆された.そのようなやわらかい会合体が,隔離膜形成においてどのような分子機能を担うのか,今後明らかにする必要のある重要な課題である.
オートファジー始動複合体によるPASの構築ののち,他の主要Atg因子群がPASに濃縮され,隔離膜の形成が進行する.Atg2は主要Atg因子中最も大きいタンパク質であり,出芽酵母の場合1592アミノ酸からなる.1993年に報告された酵母オートファジーの不能変異株スクリーニングで同定された最古参Atg因子のうちの一つであるが23),その配列は既知のタンパク質,ドメインやモチーフと相同性を持たず,構造および分子機能の情報は長らくまったくの謎であった.Atg2はAtg18と結合して,オートファゴソーム形成に関与する六つの機能グループのうちの一つを形成する2).Atg18はホスファチジルイノシトール3-リン酸(PI3P)結合タンパク質であり,PI3P依存的にAtg2-Atg18複合体はPASに移行する24).まずAtg18の構造生物学が進展し,2012年にそのホモログの結晶構造が複数グループから報告され,PI3Pを認識する構造基盤が明らかとなったが25–27),Atg2の構造研究はその後もしばらく難航した.2013年になり,Atg2-Atg18複合体は伸長中の隔離膜先端に局在すること,そこで小胞体-隔離膜間のコンタクトサイトを形成することが報告され,隔離膜伸長に直接関与することが強く示唆された28, 29).Atg2のN末端およびC末端の短い領域は,Vps13と弱い配列相同性を示すが,Vps13もまたミトコンドリア-液胞間などのコンタクトサイトを形成することから30),これらタンパク質には脂質膜どうしを繋留する共通した構造を持つことが予想された.そして2017年から,Atg2の構造生物学研究が夜明けを迎え,その分子機能の謎が明らかとなり始めた31).
哺乳類にはAtg2ホモログが二つ(ATG2Aおよび2B),Atg18ホモログが四つ(WIPI1~4)存在する.ATG2A-WIPI4およびATG2B-WIPI4複合体に関して,ネガティブ染色による電子顕微鏡解析が行われ,低分解能ではあるが全体の形状が初めて明らかとなった(図4A)32, 33).ATG2は長さ20 nm程度の棍棒状の形をとっており,その一端の側面にWIPI4が結合する.これまでにいくつかのAtg18ホモログの結晶構造が報告されており,WIPI4もそれらと同様,ドーナツ状の構造をとる.高等生物のAtg2ホモログにはH/FYモチーフが保存されており,それがAtg18ホモログとの相互作用を担う32).さらに酵母Atg2も類似の構造をとることが確認されたことから,この特徴的形状はAtg2ファミリーに保存されていると考えられる.ATG2Aは曲率の大きい(サイズの小さい)リポソームに強い結合を示し,ATG2Aが棍棒構造の両側を用いて二つのリポソームを橋渡しする形で結合するようすが電子顕微鏡を用いて捉えられた33).このことは,ATG2Aが脂質膜どうしを繋留する分子機能を持つことを強く示唆したが,実際に試験管内の解析でATG2Aがリポソームどうしを繋留することが確認された33).この繋留活性に関しては酵母Atg2について詳細なドメイン解析が行われ,Atg2はN末端側およびC末端に近い領域の2か所に脂質膜結合領域を持っていること,そのどちらもリポソームの繋留活性および酵母におけるオートファジー活性に必須であることが示された34)(詳しくは中戸川の稿参照).すなわちAtg2ファミリータンパク質は,棍棒状の全体形状を持ち,脂質膜どうしを繋留する分子機能を持つことが明らかとなった.
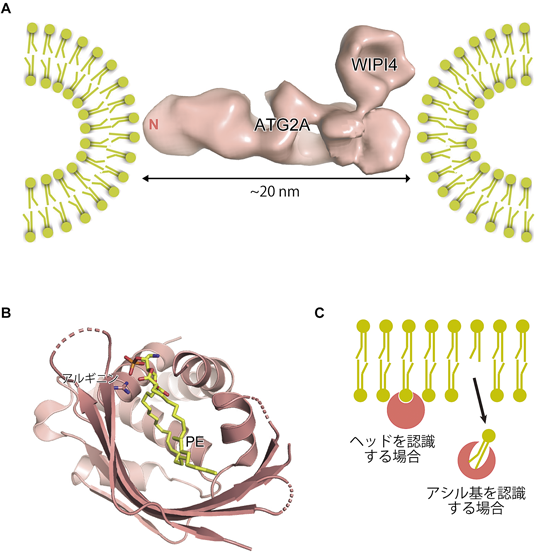
(A)ヒトATG2A-WIPI4複合体の電子顕微鏡像(EMD-8899).ATG2Aは棍棒状の構造をとり,そのN末端領域と反対側の面でそれぞれ膜に結合することで,脂質膜の繋留を担う.(B)分裂酵母Atg2のN末端領域がホスファチジルエタノールアミン(PE)を収容した状態の結晶構造(PDB 6A9J).PEおよびPEのリン酸基を認識するアルギニン側鎖をスティックモデルで示す.(C)リン脂質結合タンパク質の2通りの結合機構.
電子顕微鏡による解析が進展する中,X線結晶構造解析法による構造決定の試みも行われた.Atg2全体の結晶化の報告はこれまでないが,我々は分裂酵母Atg2のN末端領域(約240残基)に関して結晶化に成功し,分解能3.2 Åで構造決定した35).N末端領域はヘリックスに富む領域とねじれたβシート領域からなり,それらがまとまって一つの球状ドメインを形成する.その構造の最大の特徴は内部に巨大な空洞を持つ点であり,空洞内壁はほぼ疎水性アミノ酸で形成されている.この構造的特徴は,N末端領域が脂質結合能を有することを想起させたことから,続いてリン脂質の一つであるホスファチジルエタノールアミン(PE)との共結晶化を行い,複合体の構造を分解能2.7 Åで決定した(図4B).結晶中,Atg2のN末端ドメインに対しPEは1:1で複合体を形成し,アシル基部分を空洞内部へと結合させていた.PEのヘッド(エタノールアミン部分)はタンパク質外部に露出し,タンパク質との相互作用がほぼみられない一方,リン酸基部分はアルギニン残基と相互作用を形成していた.複合体構造から,Atg2のN末端領域は①リン脂質を脂質膜から引き抜いてアシル基部分を空洞に収容すること,②リン脂質のヘッド部分への特異性は持たないことが予想される.蛍光標識させたさまざまなリン脂質を含むリポソームを調製し,試験管内でAtg2と混合した結果,Atg2はどのリン脂質も類似の効率でリポソームから引き抜くことが確認された35).
N末端領域によるPEの認識が,ヘッド部分でなくアシル基部分であるということは,大きな意味を持つ.すなわちAtg2はPEが脂質二重層膜に組み込まれた状態のままでは結合できず,PEを膜から引き抜いて結合する(図4C).Atg2は小胞体膜と隔離膜の間の繋留を担うが,脂質を膜から引き抜く活性はその繋留の目的のためには意味がない.すなわちAtg2には繋留以外の重要な活性があることが示唆された.配列やトポロジー上の相同性はないが,Atg2と類似の様式でリン脂質のアシル基部分を認識するタンパク質ドメインとしてSMPが知られている.SMPドメインはミトコンドリアと小胞体間のコンタクトサイトを形成するタンパク質にみられ,脂質を膜間で転送する活性を持つ36).我々はAtg2にも同様の脂質転送活性があるのではないかと考え,試験管内で活性を調べたところ,Atg2にはリポソーム間で脂質を転送する活性があることが明らかとなった35).結晶構造が決定されたN末端領域だけでは活性が低いのに対し,全長Atg2でははるかに高い活性を示したことから,他の領域も脂質転送に関与すると考えられる.Atg2は曲率の大きい(すなわちサイズの小さい)リポソームどうしを繋留する活性がある一方,サイズの大きいリポソームどうしは繋留できない.Atg2の脂質転送活性もまたサイズの小さいリポソーム間でのみ観察され,さらに繋留活性を抑制する変異を導入したAtg2は,小さいリポソーム間でも脂質転送を示さなくなる.すなわちAtg2は自身で繋留したリポソーム間の脂質転送を担っており,繋留活性と脂質転送活性が強くリンクしていると考えらえる.ヒトのATG2Aに関しても脂質転送活性があることが報告されたことから37, 38),この活性は進化上保存されていると考えられる.Atg2のN末端領域はVps13のN末端領域と弱い配列相同性を持つが,Vps13のN末端領域もまたAtg2と同様の巨大な空洞を持ち,さらにVps13はAtg2と同様,脂質転送活性を持つ39).Vps13に膜の繋留活性があるかどうかはわかっていないが,Atg2のN末端領域をVps13のN末端領域と交換したキメラタンパク質もオートファジーを担えたことから35),両者の機能は類似していると考えられる.
オートファジー始動複合体がPASを構築したのち,数個のAtg9小胞がPASにリクルートされることで隔離膜の最初の膜源になると考えられるが9),Atg9小胞が供給する脂質はオートファゴソーム構築に必要な脂質量の1%にも満たない.したがって隔離膜が伸長してオートファゴソームになるためには,材料となる脂質を別に供給する必要がある(図1).脂質の供給源としてさまざまなオルガネラが提唱されているが,中でも小胞体が最有力候補と考えられている40).小胞体から隔離膜への脂質供給機構としては,これまでに①小胞体と隔離膜が直接連結するモデル41, 42)と②小胞輸送を介するモデル43)が提唱されてきた.今回,Atg2が脂質転送活性を有することが明らかになったことで,③小胞体と隔離膜をAtg2が橋渡しし,Atg2を介して脂質が輸送されるモデルが新たに考えられる(図5).小胞体を含む通常のオルガネラは,その機能のためにさまざまな膜タンパク質を膜成分として含んでいる.一方,上述したようにオートファゴソームの膜は膜タンパク質が顕著に少ないことが古くから知られていた1).オートファゴソームは分解に特化した一過性のオルガネラであり,形成後はリソソーム/液胞との融合がほぼ唯一の機能であることから,多種類の膜タンパク質を必要としていないのであろう.上述した3通りの脂質供給機構のうち,①と②は脂質とともに膜タンパク質も隔離膜へと流入するのに対し,③のAtg2を介したモデルでは,脂質のみの輸送が可能であると考えられる.すなわちオートファゴソーム膜の特徴を考えると,主要な脂質供給機構はAtg2を介した経路であることが示唆される.実際,出芽酵母を用いた解析において,膜繋留活性と脂質転送活性の両方を失ったAtg2では隔離膜伸長を担うことができないことが明らかとなった35).ただし膜繋留活性が①,②のメカニズムにも関係している可能性があることから,脂質転送活性のみを特異的に消失させる変異を同定し,その場合の隔離膜伸長への影響を調べることが重要である.また最近になって,COPII小胞がオートファゴソーム膜に組み込まれていることを示す証拠が見つかっていることから44)(中戸川の稿参照),小胞輸送の経路も確かに働いているようであり,3通りの脂質輸送メカニズムが並行して働いている可能性も十分にあると思われる.オートファゴソーム形成機構のさらなる理解のためには,一つのオートファゴソームを新生するのにどのメカニズムがどの程度の量の脂質供給に寄与しているのか,脂質供給源のオルガネラの問題も含めて定量的な解析を行うことが求められる.
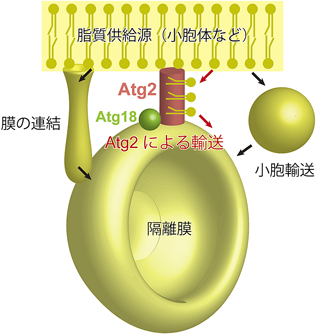
脂質供給源(小胞体など)から隔離膜への脂質供給機構は,これまで膜の連結や小胞輸送などが考えられてきたが,新たに脂質転移酵素Atg2を介した供給機構の存在が明らかとなった.
2013年に「オートファジーの構造生物学」という題の総説を本誌に執筆したが5),その「おわりに」を読み返すと,“オートファジーにおける膜動態は絶望的に複雑であり……具体的な分子機能が一向に見えてこない”と暗中模索の状況が書かれている.その後,当時は予想さえできなかった急速な研究の進展があり,難航不落と思われたオートファゴソーム形成の謎がいよいよ明らかになる,その入口に立つことができたと実感している.最後の難関である膜タンパク質Atg9の構造生物学を達成し,謎の扉が完全に開かれる日が近いうちに来ることを期待している.
本稿で紹介した内容の多くは,筆者のグループの研究成果を中心にまとめたものです.一緒に研究を進めてくれた研究室メンバーに感謝いたします.またこれまで長きにわたり共同研究をしていただき,多大なご指導,ご助言を賜りました大隅良典先生,大隅研究室の方々,そして共同研究者の方々に感謝申し上げます.最後に,私が本研究テーマを始めるきっかけを与えていただき,研究を長きにわたり支えていただきました故・稲垣冬彦先生に心から感謝申し上げます.
1) Baba, M., Osumi, M., & Ohsumi, Y. (1995) Analysis of the membrane structures involved in autophagy in yeast by freeze-replica method. Cell Struct. Funct., 20, 465–471.
2) Mizushima, N., Yoshimori, T., & Ohsumi, Y. (2011) The role of Atg proteins in autophagosome formation. Annu. Rev. Cell Dev. Biol., 27, 107–132.
3) Noda, N.N. & Inagaki, F. (2015) Mechanisms of Autophagy. Annu. Rev. Biophys., 44, 101–122.
4) Noda, N.N., Ohsumi, Y., & Inagaki, F. (2009) ATG systems from the protein structural point of view. Chem. Rev., 109, 1587–1598.
5) 野田展生(2013)オートファジーの構造生物学,生化学,85, 762–774.
6) Noda, N.N. & Fujioka, Y. (2015) Atg1 family kinases in autophagy initiation. Cell. Mol. Life Sci., 72, 3083–3096.
7) Suzuki, K., Kirisako, T., Kamada, Y., Mizushima, N., Noda, T., & Ohsumi, Y. (2001) The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation. EMBO J., 20, 5971–5981.
8) Suzuki, K., Kubota, Y., Sekito, T., & Ohsumi, Y. (2007) Hierarchy of Atg proteins in pre-autophagosomal structure organization. Genes Cells, 12, 209–218.
9) Yamamoto, H., Kakuta, S., Watanabe, T.M., Kitamura, A., Sekito, T., Kondo-Kakuta, C., Ichikawa, R., Kinjo, M., & Ohsumi, Y. (2012) Atg9 vesicles are an important membrane source during early steps of autophagosome formation. J. Cell Biol., 198, 219–233.
10) Hurley, J.H. & Yang, D. (2008) MIT domainia. Dev. Cell, 14, 6–8.
11) Fujioka, Y., Suzuki, S.W., Yamamoto, H., Kondo-Kakuta, C., Kimura, Y., Hirano, H., Akada, R., Inagaki, F., Ohsumi, Y., & Noda, N.N. (2014) Structural basis of starvation-induced assembly of the autophagy initiation complex. Nat. Struct. Mol. Biol., 21, 513–521.
12) Jao, C.C., Ragusa, M.J., Stanley, R.E., & Hurley, J.H. (2013) A HORMA domain in Atg13 mediates PI 3-kinase recruitment in autophagy. Proc. Natl. Acad. Sci. USA, 110, 5486–5491.
13) Yamamoto, H., Fujioka, Y., Suzuki, S.W., Noshiro, D., Suzuki, H., Kondo-Kakuta, C., Kimura, Y., Hirano, H., Ando, T., Noda, N.N., et al. (2016) The Intrinsically Disordered Protein Atg13 Mediates Supramolecular Assembly of Autophagy Initiation Complexes. Dev. Cell, 38, 86–99.
14) Ragusa, M.J., Stanley, R.E., & Hurley, J.H. (2012) Architecture of the Atg17 complex as a scaffold for autophagosome biogenesis. Cell, 151, 1501–1512.
15) Kamada, Y., Funakoshi, T., Shintani, T., Nagano, K., Ohsumi, M., & Ohsumi, Y. (2000) Tor-mediated induction of autophagy via an Apg1 protein kinase complex. J. Cell Biol., 150, 1507–1513.
16) Noda, N.N. & Mizushima, N. (2016) Atg101: Not Just an Accessory Subunit in the Autophagy-initiation Complex. Cell Struct. Funct., 41, 13–20.
17) Suzuki, H., Kaizuka, T., Mizushima, N., & Noda, N.N. (2015) Structure of the Atg101-Atg13 complex reveals essential roles of Atg101 in autophagy initiation. Nat. Struct. Mol. Biol., 22, 572–580.
18) Kim, B.W., Jin, Y., Kim, J., Kim, J.H., Jung, J., Kang, S., Kim, I.Y., Kim, J., Cheong, H., & Song, H.K. (2018) The C-terminal region of ATG101 bridges ULK1 and PtdIns3K complex in autophagy initiation. Autophagy, 14, 2104–2116.
19) Qi, S., Kim, D.J., Stjepanovic, G., & Hurley, J.H. (2015) Structure of the Human Atg13-Atg101 HORMA Heterodimer: an Interaction Hub within the ULK1 Complex. Structure, 23, 1848–1857.
20) Michel, M., Schwarten, M., Decker, C., Nagel-Steger, L., Willbold, D., & Weiergraber, O.H. (2015) The mammalian autophagy initiator complex contains 2 HORMA domain proteins. Autophagy, 11, 2300–2308.
21) Luo, X. & Yu, H. (2008) Protein metamorphosis: the two-state behavior of Mad2. Structure, 16, 1616–1625.
22) Memisoglu, G., Eapen, V.V., Yang, Y., Klionsky, D.J., & Haber, J.E. (2019) PP2C phosphatases promote autophagy by dephosphorylation of the Atg1 complex. Proc. Natl. Acad. Sci. USA, 116, 1613–1620.
23) Tsukada, M. & Ohsumi, Y. (1993) Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett., 333, 169–174.
24) Obara, K., Sekito, T., Niimi, K., & Ohsumi, Y. (2008) The Atg18-Atg2 complex is recruited to autophagic membranes via phosphatidylinositol 3-phosphate and exerts an essential function. J. Biol. Chem., 283, 23972–23980.
25) Watanabe, Y., Kobayashi, T., Yamamoto, H., Hoshida, H., Akada, R., Inagaki, F., Ohsumi, Y., & Noda, N.N. (2012) Structure-based analyses reveal distinct binding sites for Atg2 and phosphoinositides in Atg18. J. Biol. Chem., 287, 31681–31690.
26) Baskaran, S., Ragusa, M.J., Boura, E., & Hurley, J.H. (2012) Two-site recognition of phosphatidylinositol 3-phosphate by PROPPINs in autophagy. Mol. Cell, 47, 339–348.
27) Krick, R., Busse, R.A., Scacioc, A., Stephan, M., Janshoff, A., Thumm, M., & Kuhnel, K. (2012) Structural and functional characterization of the two phosphoinositide binding sites of PROPPINs, a beta-propeller protein family. Proc. Natl. Acad. Sci. USA, 109, E2042–E2049.
28) Suzuki, K., Akioka, M., Kondo-Kakuta, C., Yamamoto, H., & Ohsumi, Y. (2013) Fine mapping of autophagy-related proteins during autophagosome formation in Saccharomyces cerevisiae. J. Cell Sci., 126, 2534–2544.
29) Graef, M., Friedman, J.R., Graham, C., Babu, M., & Nunnari, J. (2013) ER exit sites are physical and functional core autophagosome biogenesis components. Mol. Biol. Cell, 24, 2918–2931.
30) Lang, A.B., John Peter, A.T., Walter, P., & Kornmann, B. (2015) ER-mitochondrial junctions can be bypassed by dominant mutations in the endosomal protein Vps13. J. Cell Biol., 210, 883–890.
31) Osawa, T. & Noda, N.N. (2019) Atg2: A novel phospholipid transfer protein that mediates de novo autophagosome biogenesis. Protein Sci., 28, 1005–1012.
32) Zheng, J.X., Li, Y., Ding, Y.H., Liu, J.J., Zhang, M.J., Dong, M.Q., Wang, H.W., & Yu, L. (2017) Architecture of the ATG2B-WDR45 complex and an aromatic Y/HF motif crucial for complex formation. Autophagy, 13, 1870–1883.
33) Chowdhury, S., Otomo, C., Leitner, A., Ohashi, K., Aebersold, R., Lander, G.C., & Otomo, T. (2018) Insights into autophagosome biogenesis from structural and biochemical analyses of the ATG2A-WIPI4 complex. Proc. Natl. Acad. Sci. USA, 115, E9792–E9801.
34) Kotani, T., Kirisako, H., Koizumi, M., Ohsumi, Y., & Nakatogawa, H. (2018) The Atg2-Atg18 complex tethers pre-autophagosomal membranes to the endoplasmic reticulum for autophagosome formation. Proc. Natl. Acad. Sci. USA, 115, 10363–10368.
35) Osawa, T., Kotani, T., Kawaoka, T., Hirata, E., Suzuki, K., Nakatogawa, H., Ohsumi, Y., & Noda, N.N. (2019) Atg2 mediates direct lipid transfer between membranes for autophagosome formation. Nat. Struct. Mol. Biol., 26, 281–288.
36) Reinisch, K.M. & De Camilli, P. (2016) SMP-domain proteins at membrane contact sites: Structure and function. Biochim. Biophys. Acta, 1861(8 Pt B), 924–927.
37) Valverde, D.P., Yu, S., Boggavarapu, V., Kumar, N., Lees, J.A., Walz, T., Reinisch, K.M., & Melia, T.J. (2019) ATG2 transports lipids to promote autophagosome biogenesis. J. Cell Biol., 218, 1787–1798.
38) Maeda, S., Otomo, C., & Otomo, T. (2019) The autophagic membrane tether ATG2A transfers lipids between membranes. Elife, 8, e45777.
39) Kumar, N., Leonzino, M., Hancock-Cerutti, W., Horenkamp, F.A., Li, P., Lees, J.A., Wheeler, H., Reinisch, K.M., & De Camilli, P. (2018) VPS13A and VPS13C are lipid transport proteins differentially localized at ER contact sites. J. Cell Biol., 217, 3625–3639.
40) Axe, E.L., Walker, S.A., Manifava, M., Chandra, P., Roderick, H.L., Habermann, A., Griffiths, G., & Ktistakis, N.T. (2008) Autophagosome formation from membrane compartments enriched in phosphatidylinositol 3-phosphate and dynamically connected to the endoplasmic reticulum. J. Cell Biol., 182, 685–701.
41) Hayashi-Nishino, M., Fujita, N., Noda, T., Yamaguchi, A., Yoshimori, T., & Yamamoto, A. (2009) A subdomain of the endoplasmic reticulum forms a cradle for autophagosome formation. Nat. Cell Biol., 11, 1433–1437.
42) Yla-Anttila, P., Vihinen, H., Jokitalo, E., & Eskelinen, E.L. (2009) 3D tomography reveals connections between the phagophore and endoplasmic reticulum. Autophagy, 5, 1180–1185.
43) Ge, L., Zhang, M., Kenny, S.J., Liu, D., Maeda, M., Saito, K., Mathur, A., Xu, K., & Schekman, R. (2017) Remodeling of ER-exit sites initiates a membrane supply pathway for autophagosome biogenesis. EMBO Rep., 18, 1586–1603.
44) Shima, T., Kirisako, H., & Nakatogawa, H. (2019) COPII vesicles contribute to autophagosomal membranes. J. Cell Biol., 218, 1503–1510.

公益財団法人微生物化学研究会微生物化学研究所部長.博士(薬学).
1996年東京大学薬学部卒業.2001年同大学院薬学系研究科博士課程修了.同年より北海道大学大学院薬学研究院にてポスドク,助教,講師を歴任.11年より現所属・主席研究員,17年より現職.
研究テーマと抱負研究テーマはオートファジーの分子機構の解明.構造生物学が専門であるが,真の意味で“生物学”と言える研究をすること,“構造のない”タンパク質の構造の謎に迫ることが現在の目標.
ウェブサイトhttp://www.bikaken.or.jp
趣味オートキャンプ,スキー,登山.

This page was created on 2019-08-23T11:31:43.121+09:00
This page was last modified on 2019-10-08T09:40:49.000+09:00
このサイトは(株)国際文献社によって運用されています。