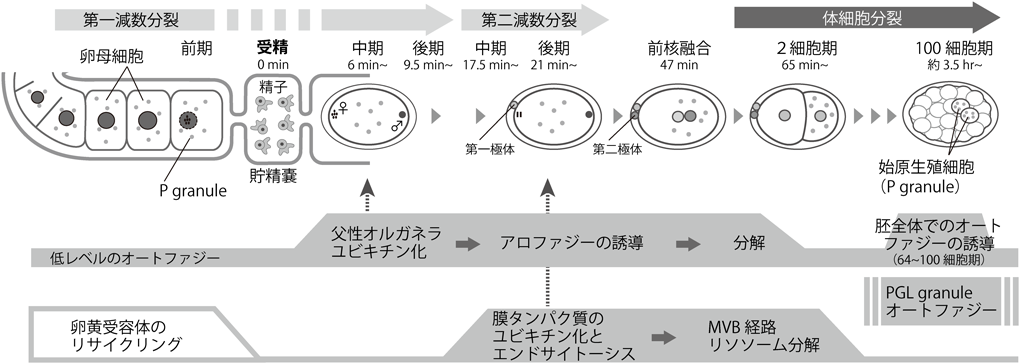
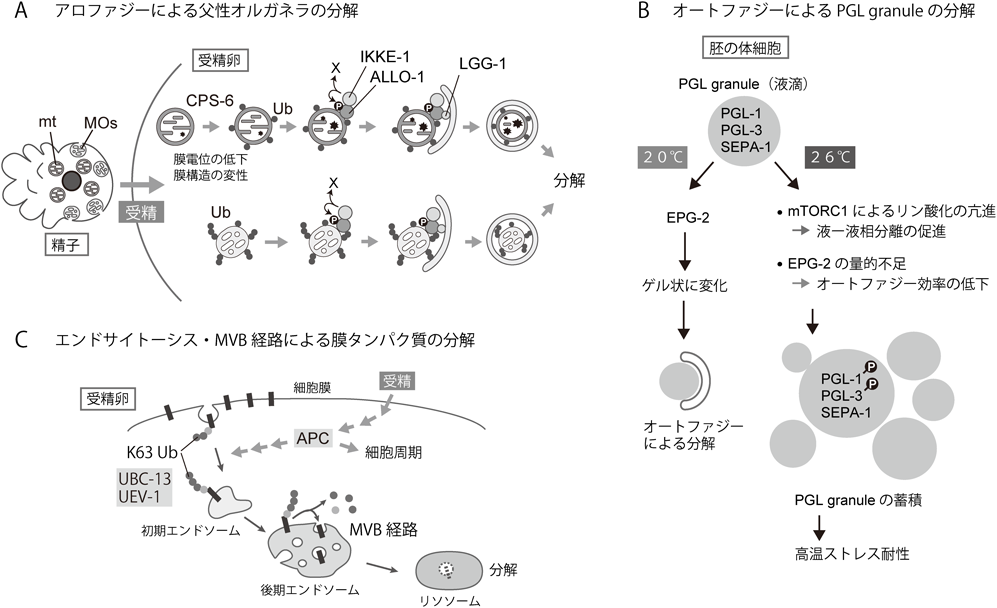
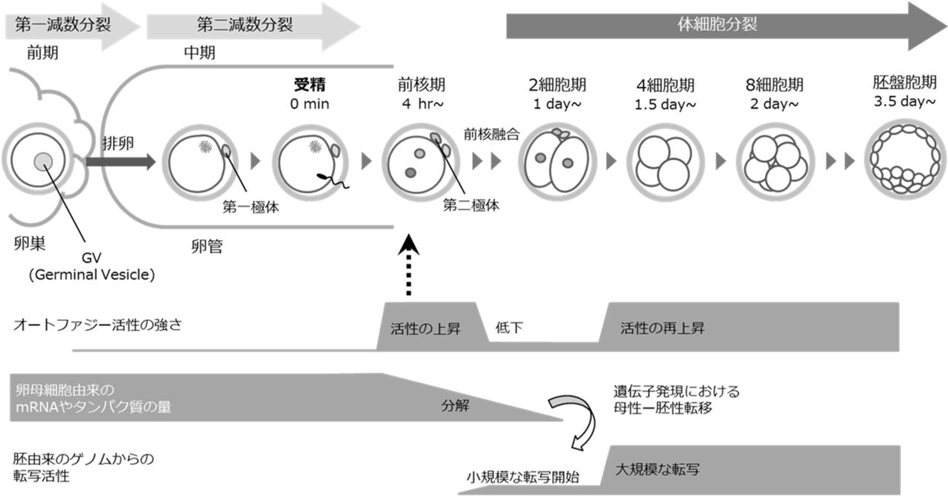
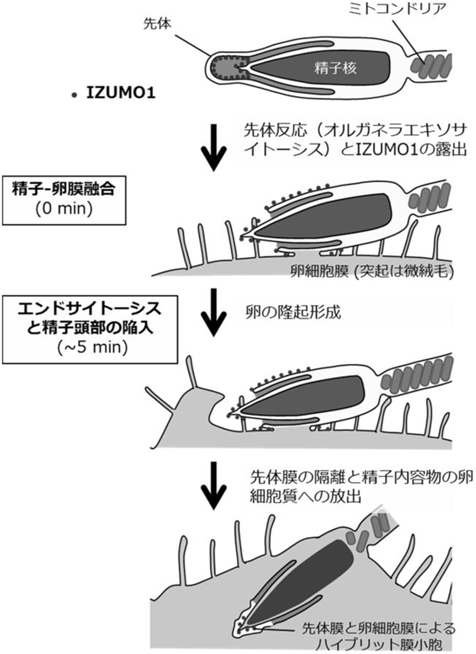
初期発生におけるリソソーム分解の生理機能と分子メカニズムHow lysosomal degradation systems function in early embryos
1 群馬大学生体調節研究所生体膜機能分野Laboratory of Molecular Membrane Biology, Institute for Molecular and Cellular Regulation, Gunma University ◇ 群馬県前橋市昭和町3–39–15 ◇ 3–39–15 Showa, Maebashi, Gunma, Japan
2 群馬大学生体調節研究所細胞構造分野Laboratory of Molecular Traffic, Institute for Molecular and Cellular Regulation, Gunma University ◇ 群馬県前橋市昭和町3–39–15 ◇ 3–39–15 Showa, Maebashi, Gunma, Japan
発行日:2019年10月25日Published: October 25, 2019