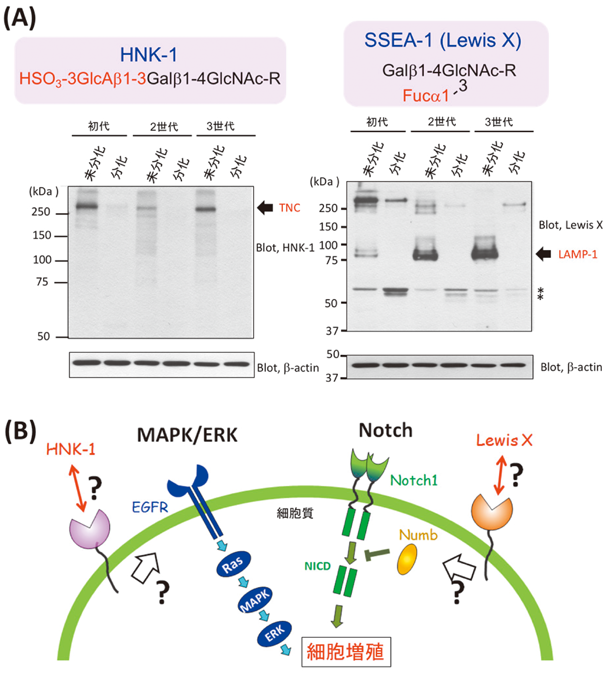
自然界に存在するタンパク質の半数以上は糖鎖修飾を受けた糖タンパク質として存在しており,糖鎖は生体組織形成や生体防御などにおいて不可欠な役割を担っている.糖鎖の生合成は,糖転移酵素や糖加水分解酵素により逐次的に糖残基が付加・脱離されることで行われる.さらには,糖転移酵素の基質である糖ヌクレオチドの存在量によっても糖鎖の発現パターンは大きく異なることが知られている.このように糖鎖の配列情報はゲノムに直接コードされていないため,その構造は予測困難であり,しかも一定しておらず,著しい不均一性を伴っていることが特徴となっている.しかしながら,このように一見無秩序にみえるタンパク質の糖鎖修飾プロセスのうちにも,糖鎖構造の多様性を制御する何らかのメカニズムが存在する可能性が考えられる.たとえば,代表的な血清糖タンパク質である免疫グロブリンG(IgG)のFc領域を修飾している糖鎖は,(生理状態の変化や病態に伴う末端構造の変動はあるものの)常に1対の二本鎖複合型糖鎖を呈している1).また,細胞間接着に関わるラミニン結合性糖鎖は,ジストログリカンという特定のタンパク質の上の特定の部位にのみ結合している2–4).筆者らはこれまでに,神経幹細胞の分化前後において,その発現パターンの異なる糖鎖が,積極的に細胞の分化過程や幹細胞性の維持を制御しているとの予想のもと,糖鎖の発現プロファイリングを行ってきた.その結果,神経幹細胞の分化前後において糖タンパク質糖鎖の発現パターンは大きく異なっており,特に,未分化の神経幹細胞にLewis X[Galβ1-4(Fucα1-3)GlcNAc]およびHNK-1[HSO3-3GlcAβ1-3Galβ1-4GlcNAc-]を有するN型糖鎖が特異的に発現していることを明らかにするとともに,これらのグライコトープは特定のタンパク質にのみ結合していることを見いだしている5–7)(図1).
このようにゲノムに制御されていないはずの糖鎖修飾が,あたかも設計図に従うかのようにタンパク質特異的に起きている例が存在する.しかしながら,こうした制御がいかなる分子機構を通じて実現されているかということは,体系的にこれまでほとんど理解されていない.たとえば,古くから研究されているマンノース6-リン酸化修飾でさえ,多くの分泌タンパク質の中で,なぜリソソームタンパク質のみにマンノース6-リン酸修飾が付与されるのかいまだ明らかになっていない.
本稿では,これまでの筆者らの研究成果を含め,徐々に明らかになりつつあるタンパク質特異的な糖鎖修飾機構について議論したい.
2. 基質タンパク質と糖転移酵素の直接的な相互作用によるタンパク質特異的な糖鎖修飾
タンパク質のリン酸化においては,各種キナーゼがそれぞれ基質となるタンパク質のアミノ酸配列を認識して,タンパク質特異的なリン酸化修飾が行われていることが知られている8).また,タンパク質に対して直接グルコース(Glc),フコース(Fuc),N-アセチルガラクトサミン(GalNAc)残基を付加する酵素も,タンパク質部分を認識することで,配列特異性を有することが明らかになっている9–11).一方,単糖どうしを連結する反応を担う糖転移酵素においても,酵素が基質タンパク質の糖鎖部分のみならず,タンパク質部分を認識することで,タンパク質特異性を発揮している可能性が考えられる.実際に,ポリシアル酸,ラミニン結合性糖鎖,HNK-1糖鎖の生合成に関する酵素は,基質糖タンパク質の糖鎖部分だけでなくタンパク質部分を認識して,特異的な糖鎖修飾を担っていると考えられている.
1)ポリシアル酸
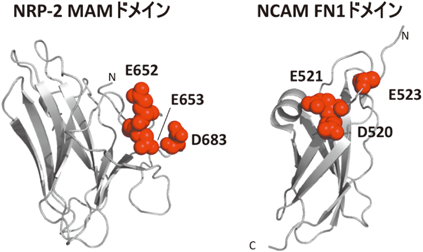
ポリシアル酸は,N型糖鎖やO型糖鎖の還元末端に結合しており,シアル酸がα2–8結合で数残基から100残基以上連なった構造を形成している12).特に,そのかさ高さと負電荷から,細胞–細胞,細胞–細胞外マトリックス間の接着を阻害したり,神経栄養因子や神経伝達物質と特異的に結合することで,神経系の形成や機能発現に関与することが知られている.ポリシアル酸糖鎖は,neural cell adhesion molecule(NCAM), neuropilin-2(NRP-2), synaptic cell adhesion molecule 1, voltage-dependent sodium channelといった数種類の細胞表層のタンパク質に特異的に結合している.NCAMやNRP-2を対象とした研究により,ポリシアル酸の合成に関わるシアル酸転移酵素PST(ST8SiaIV)とキャリアタンパク質が直接的に相互作用することにより,本糖鎖がタンパク質特異性を獲得していることが見いだされている13–15).具体的には,NCAMやNRP-2は,タンパク質の表面に酸性アミノ酸3残基から構成される負電荷のクラスターが存在し(図2),本領域がPSTとの相互作用を介して,ポリシアル酸の伸長を担っていることが示されている.しかしながら,NCAMとNRP-2の負電荷クラスターが存在しうるドメイン構造や近傍の構造に類似性が認められないことから(図2),PSTがいかにして異なる基質タンパク質を認識しているかは不明である.さらにはこうした負電荷クラスターはさまざまな細胞表層タンパク質に認められることに加えて,キャリアタンパク質のポリシアル酸が結合している部位が制限されていることを考慮すると,負電荷クラスター領域とポリシアル酸糖鎖の修飾部位の空間的な配置もタンパク質特異的なポリシアル酸修飾の決定要因であることが予想される.
2)ラミニン結合性糖鎖
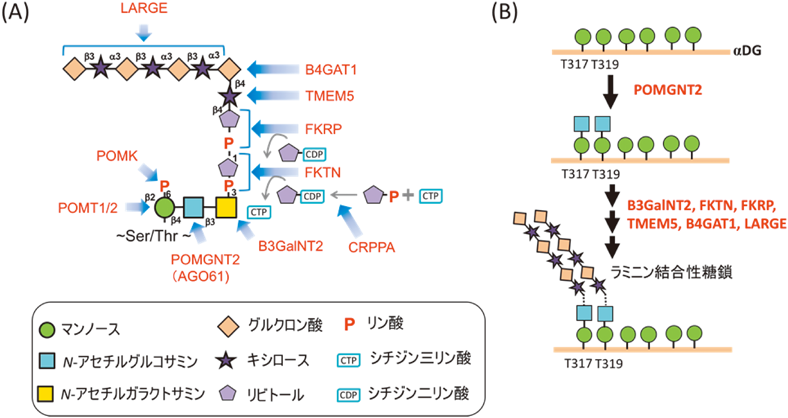
細胞間接着に関わるラミニン結合糖鎖は,その形成不全により筋ジストロフィーが発症することが知られている.ここ10年間で,ラミニン結合性糖鎖の合成に関わる糖転移酵素が次々に同定され,2016年にその全体構造が明らかにされている(図3A)2–4, 16).その構造は,リン酸化3糖構造(ManP-GlcNAc-GalNAc)の先にリビトールリン酸(RboP)がホスホジエステル結合を介してタンデムに結合し,その非還元末端側にキシロース(Xyl)とグルクロン酸(GlcA)からなるリピート配列が続いている.興味深いことに,本糖鎖は,αジストログリカン(αDG)上にしかその存在が見いだされておらず,なかでも,本タンパク質のThr317/Thr319サイトのみに結合していることが知られている17).こうしたタンパク質特異的な糖鎖修飾に関しては,Xyl-GlcAリピート配列の合成を担うバイファンクショナルな酵素であるLARGEがαDGのN末端ドメインと直接相互作用することで,αDG上に特異的にラミニン結合性糖鎖が合成されると考えられている18).さらに,筆者らの研究グループでも,リン酸化3糖構造のGlcNAcの合成を担う酵素であるPOMGNT2(AGO61)がαDG上の特定の位置に結合したマンノース(Man)残基へのN-アセチルグルコサミン(GlcNAc)修飾を担っていることを見いだしている(図3B)19).興味深いことにその修飾箇所は,これまでラミニン結合性糖鎖が結合していると報告されているトレオニン残基(Thr317/Thr319)と一致していた.つまり,POMGNT2がαDG上のラミニン結合性を示す糖鎖の形成部位を決定する重要な酵素であることを示している.このようにラミニン結合性糖鎖は,複数の酵素により基質タンパク質が認識されることで,特定の基質上の特定のアミノ酸残基に糖鎖修飾が生じるような細やかな制御を受けているものと考えられる.
3)HNK-1糖鎖
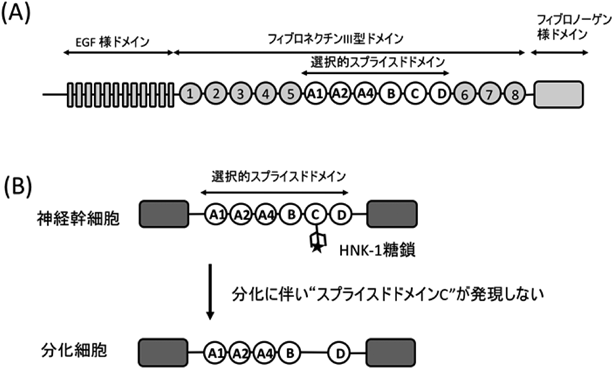
HNK-1糖鎖は,GlcAの3位に硫酸が結合した特徴的な構造を有しており,N型およびO型糖鎖上に存在する.本糖鎖の修飾に関わる酵素として,GlcA転移酵素(GlcAT-P, GlcAT-S)および硫酸転移酵素(HNK-1ST)が同定されており,これら酵素が複合体を形成することで,効率のよい糖鎖の構造形成が行われている20).HNK-1糖鎖も,NCAM, L1, myelin-associated glycoprotein(MAG)といったIgドメインを有する接着分子や細胞外マトリックス分子の一種であるTenascin-R上に特異的に発現し,神経細胞の接着や移動,神経突起の伸長の制御を担っている.筆者らも,神経幹細胞においてHNK-1を有する糖鎖がTenascin-C(TNC)のみに特異的に結合しており,しかもこの糖鎖が,epidermal growth factor(EGF)受容体の発現量を調節することでmitogen-activated protein kinase(MAPK)経路を活性化し,神経幹細胞の幹細胞性の維持に関与していることを見いだしてきた(図1B)5).興味深いことに,このTNCタンパク質上のHNK-1糖鎖は,特定のスプライスドドメイン上にのみ発現しており,分化が進むにつれてそのスプライスドドメインを有するTNCのアイソフォームの発現が減少し,それに伴ってHNK-1の発現が消失していた(図4).つまり,HNK-1糖鎖の生合成に関わる糖転移酵素の発現量を変化させることなく,HNK-1糖鎖の発現を制御していることが明らかとなっている.TNCが有する6個のスプライスドドメインは,すべて同じフィブロネクチンIII型ドメイン構造をしているにもかかわらず,特定のドメインのみにHNK-1糖鎖が発現することから,本糖鎖の修飾に関わるGlcATなどが当該ドメインに特徴的なアミノ酸配列を読み取ることで,ドメイン特異的なHNK-1糖鎖を合成しているものと予想される.
3. 基質タンパク質の細胞内輸送の違いによるタンパク質特異的な糖鎖修飾
上記で述べたように,ある種の糖転移酵素については基質のタンパク質部分が直接相互作用することで,タンパク質特異的な糖鎖修飾が行われているものと考えられる.一方,糖タンパク質の分泌経路においては,小胞体で合成された糖タンパク質がゴルジ体を通る過程でさまざまな糖加水分解酵素および糖転移酵素と遭遇し,糖鎖の構造が形成されていく.このような過程で基質タンパク質と糖転移酵素の出会う機会が制御され,特定の糖鎖修飾が行われている可能性がある.
たとえば,傍絞輪部における軸索グリア接合に関与しているcontactin-1は,相互作用分子であるparanodinと細胞内で複合体を形成する場合と形成しない場合とで異なる分泌経路を経由した後,細胞表層に発現することが知られている21).すなわち,paranodinと複合体を形成したcontactin-1は高マンノース型糖鎖を発現しているが,複合体を形成しない場合には複合型糖鎖を発現していることが示されている.このようにcontactin-1はその細胞内の分泌経路に応じて,異なる糖鎖修飾を受ける.
一方で,ショウジョウバエにおいては,糖タンパク質の種類によって,輸送されるゴルジ体が異なることが示されており22),糖鎖修飾の種類によって専用のゴルジ体が形成されていることが考えられている.筆者もごく最近,これまでトランスゴルジに局在することが知られている数種類の糖転移酵素が実はゴルジ体内で異なる局域に存在していることを見いだしつつある(未発表データ).これらの知見に鑑みると,基質タンパク質は自身に組み込まれている特異的なアミノ配列によりその細胞内輸送が規定されており,これにより基質タンパク質が特定の糖転移酵素の局在するゴルジ体の領域に送り込まれて特異的な糖鎖修飾を受けるという機構が存在する可能性も想定される.
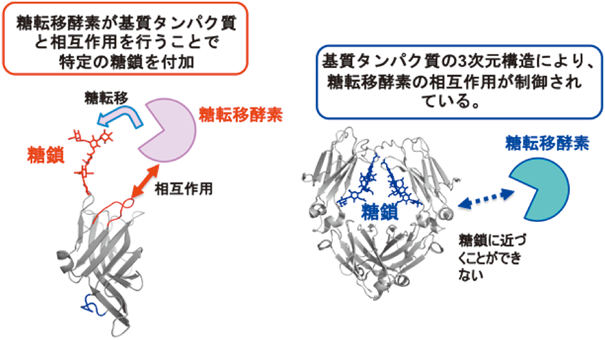
4. 基質タンパク質の3次元立体構造による糖鎖修飾の特異性制御
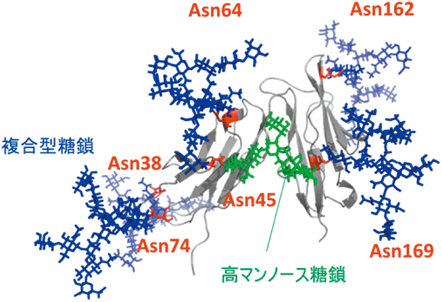
糖鎖はタンパク質上に発現しているため,糖鎖転移酵素が糖鎖を伸長する場合,基質タンパク質の構造が影響を与えることは想像にかたくない.筆者らはFcγ受容体IIIの部位特異的な糖鎖分析から,分子中の6か所のN型糖鎖修飾部位のうちの特定の1か所が高マンノース型糖鎖を発現していることを見いだしている(図5)23, 24).高マンノース型糖鎖はN型糖鎖形成過程の初期構造であるため,本糖鎖は糖加水分解酵素や糖転移酵素によるプロセッシングを受けていないことが考えられる.つまり,糖タンパク質の3次元構造が規定する特徴によって,タンパク質に発現している糖鎖の結合部位ごとに,糖加水分解酵素や糖転移酵素との立体障害の違いが生まれ,タンパク質上の部位特異的な糖鎖構造が生じうる可能性が想定される.実際に,E-cadherin上でN-acetylglucosaminyl transferase(GnT)-V依存的なGlcNAc修飾を受けるN型糖鎖の発現部位は,E-cadherinの立体的特徴によりGnT-Vが糖鎖に接近できるか否かで規定されていることが報告されている25).
免疫グロブリンG(IgG)は,タンパク質分子の内部に1対のN型糖鎖を包含するユニークな四次構造を有しているが,これらの糖鎖はシアル酸(Sia),ガラクトース(Gal),Fuc残基の有無による不均一性を示すものの,興味深いことにそれらの構造はもっぱら二本鎖複合型糖鎖に限られている.タンパク質の内部に糖鎖が発現しているため,分岐構造の形成に関与するGnT-IV, GnT-V, GnT-VIなどが糖鎖にアクセスできない可能性があるが,分子の内部構造に位置するGlcNAcがフコース転移酵素8(FUT8)によってフコシル化されるという事実は,大変興味深い.一方,筆者らは,これまでにIgYの糖鎖の構造解析を通じて,細胞内における四次構造形成のタイミングが糖鎖修飾に関連している可能性を見いだしてきた.
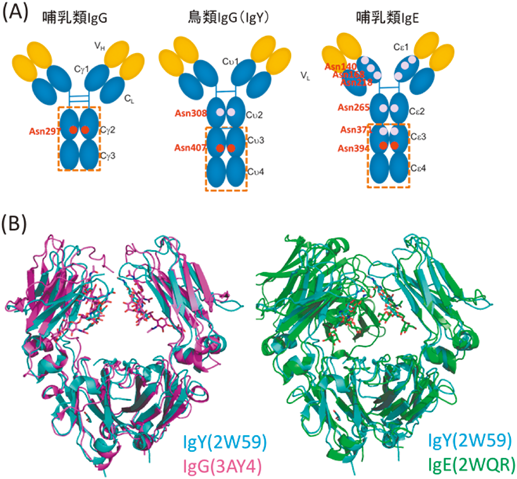
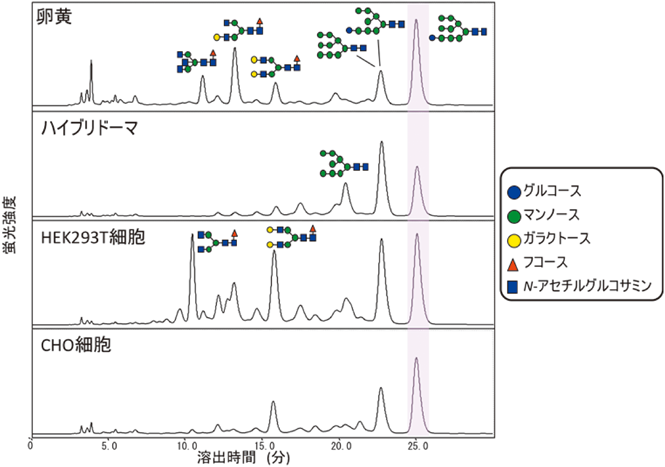
卵黄に豊富に含まれるIgYは免疫生化学試薬として利用されており,近年では医療応用の可能性も期待されている.IgYのFc領域のAsn407にはN型糖鎖結合部位が存在し,この部位は哺乳類のIgクラスに保存された糖鎖修飾部位にあたる(図6A).こうした類似性にもかかわらず,ニワトリ血清および卵黄由来のIgYのAsn407は,哺乳類の分泌された糖タンパク質にはほとんどみられないモノグルコシル化された糖鎖が結合している26).筆者らは,可変領域のアミノ酸配列が共通なIgYを異なる産生細胞(ハイブリドーマ,CHO, HEK293T)に発現させ,これらリコンビナントIgYのN型糖鎖プロファイリングを比較解析した.その結果,興味深いことに,これらすべてのリコンビナントIgYは,共通してモノグルコシル化糖鎖を多く発現していた(図7)26).一方で,IgYと相同な四次構造を有するIgEも,Igクラスに保存されたAsn394にN型糖鎖を有するが(図6),それらはモノグルコシル化糖鎖ではく,もっぱら二本鎖複合型糖鎖である27).
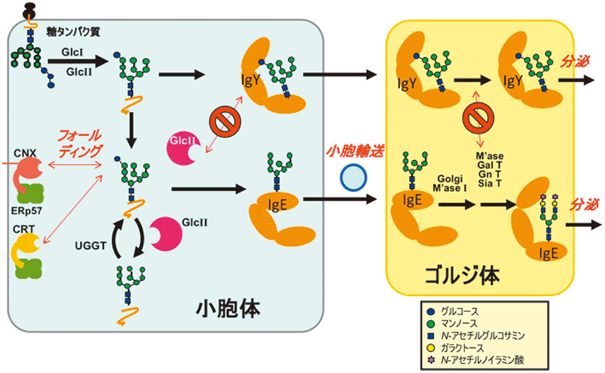
分泌過程における糖タンパク質の糖鎖のトリミングを考えると,IgYに認められる特徴的なモノグルコシル化糖鎖の発現は,IgYのFc領域が四次構造を形成することによりAsn407糖鎖が分子の内部に遮蔽され,小胞体内のグルコシダーゼなどの糖加水分解酵素から隔絶することと関連しているもの予想される(図8).一方で四次構造が類似しているIgEにはモノグルコシル化糖鎖が認められないことから,IgYとIgEではN型糖鎖プロセッシングと四次構造形成のタイミングおよび効率がかなり異なっていることが考えられる.しかもこうした性質は,細胞の状態や種類によって大きく影響を受けることが考えられ,実際にリコンビナントIgYの生産に用いた細胞の違いによってモノグルコシル化糖鎖の発現度合いには有意な差が認められる(図8).
このように,タンパク質の立体構造や四次構造形成のタイミングは,基質タンパク質に対する糖鎖転移酵素および糖加水分解酵素のアクセシビリティーと密接に関わっており,その結果としてタンパク質特異的さらには部位特異的な糖鎖修飾が生じている可能性がある.
本稿で記載したように,基質タンパク質の糖鎖修飾の特異性を規定する要因として,「基質タンパク質中の特定のアミノ酸配列」と「基質タンパク質の3次元構造」があげられる(図9).特定のアミノ酸配列が糖転移酵素との直接的な相互作用を媒介している可能性に加えて,特定配列により細胞内輸送が限定されることで基質タンパク質と糖転移酵素の出会いが制御されるという可能性も想定される.一方,基質タンパク質の3次元構造上の特徴により,糖加水分解酵素や糖転移酵素との立体障害がもたらされ,結果的に特定の基質タンパク質の特定の部位に特異的な糖鎖修飾が生じている可能性も考えられる.タンパク質の立体構造がそのアミノ酸配列によって一義的に決定されていることを考慮すれば,これまでおよそ無秩序に多様と考えられていた糖タンパク質の糖鎖構造も,その基本的な枠組みはゲノムの中にあらかじめコードされている,と言い表すこともできるだろう.
近年のグライコプロテオミクスの進展により,特定の糖鎖の構造を発現しているタンパク質の同定,さらにはそれらの修飾部位に関する情報が網羅的に解析されるようになってきた29).このような解析から,基質タンパク質のアミノ酸配列に組み込まれた糖鎖修飾の制御コードが系統的に見いだされてくるかもしれない.一方で,そうした制御コードを構成するアミノ酸残基は,配列上では離れていても立体構造上では近接しているという可能性もある.こうしたことから,糖鎖修飾コードを見いだすには,糖鎖構造に焦点を当てたグライコプロテオミクスに加えて,基質タンパク質の立体構造の情報も合わせて考慮する必要があるであろう.また,基質タンパク質の立体構造だけでなく,糖鎖プロセッシングと四次構造の形成のタイミングなどによってもタンパク質糖鎖修飾が規定されうることを考慮すると,糖タンパク質の細胞内輸送と高次構造形成の連関に関する知見を蓄積することも重要である.
今後,基質タンパク質のアミノ酸配列に組み込まれたコードによって糖鎖修飾の特異性が決定されている分子メカニズムを体系的に解明することにより,タンパク質の翻訳後修飾機構の基礎的理解が深まるものと期待している.こうした「糖鎖修飾を規定する分子コード」を読み解くことができれば,それを活用してバイオ医薬品をはじめとするタンパク質の糖鎖修飾を人為的に制御することも可能となるかもしれない.
謝辞Acknowledgments
本稿で紹介した研究成果の一部は文部科学省・日本学術振興会科学研究費補助金による支援を得て行われたものです.ここに謝意を表します.また,本稿に記載しさせていただきました筆者の研究は主に,名古屋市大学大学院薬学研究科で行ったものであり,本研究を推進するあたり,有益なご助言と激励をいただきました加藤晃一教授に感謝いたします.また,適切なご助言や有益なデスカッションをしていただきました共同研究者の先生方,ともに研究を行ってくれた学生ならび同僚の皆様にこの場を借りてお礼申し上げます.
引用文献References
1) Yamada, E., Tsukamoto, Y., Sasaki, R., Yagyu, K., & Takahashi, N. (1997) Structural changes of immunoglobulin G oligosaccharides with age in healthy human serum. Glycoconj. J., 14, 401–405.
2) Kanagawa, M., Kobayashi, K., Tajiri, M., Manya, H., Kuga, A., Yamaguchi, Y., Akasaka-Manya, K., Furukawa, J., Mizuno, M., Kawakami, H., et al. (2016) Identification of a post-translational modification with ribitol-phosphate and its defect in muscular Dystrophy. Cell Rep., 14, 2209–2223.
3) Yagi, H., Kuo, C.-W., Obayashi, T., Ninagawa, S., Khoo, K.-H., & Kato, K. (2016) Direct mapping of additional modifications on phosphorylated O-glycans of α-dystroglycan by mass spectrometry analysis in conjunction with knocking out of causative genes for dystroglycanopathy. Mol. Cell. Proteomics, 15, 3424–3434.
4) Manya, H. & Endo, T. (2017) Glycosylation with ribitol-phosphate in mammals: New insights into the O-mannosyl glycan. Biochim. Biophys. Acta, Gen. Subj., 1861, 2462–2472.
5) Yagi, H., Yanagisawa, M., Suzuki, Y., Nakatani, Y., Ariga, T., Kato, K., & Yu, R.K. (2010) HNK-1 epitope-carrying tenascin-C spliced variant regulates the proliferation of mouse embryonic neural stem cells. J. Biol. Chem., 285, 37293–37301.
6) Yagi, H., Yanagisawa, M., Kato, K., & Yu, R.K. (2010) Lysosome-associated membrane protein 1 is a major SSEA-1-carrier protein in mouse neural stem cells. Glycobiology, 20, 976–981.
7) Yagi, H., Saito, T., Yanagisawa, M., Yu, R.K., & Kato, K. (2012) Lewis X-carrying N-glycans regulate the proliferation of mouse embryonic neural stem cells via the notch signaling pathway. J. Biol. Chem., 287, 24356–24364.
8) Ubersax, J.A. & Ferrell, J.E. Jr. (2007) Mechanisms of specificity in protein phosphorylation. Nat. Rev. Mol. Cell Biol., 8, 530–541.
9) Narimatsu, Y., Joshi, H., Schjoldager, K.T., Hintze, J., Halim, A., Steentoft, C., Nasson, R., Mandel, U., Bennett, E.P., Clausen, H., et al. (2019) Exploring regulation of protein O-glycosylation in isogenic human HEK293 cells by differential O-glycoproteomics. Mol. Cell. Proteomics, 18, 1396–1409.
10) Takeuchi, H., Schneider, M., Williamson, D.B., Ito, A., Takeuchi, M., Handford, P.A., & Haltiwanger, R.S. (2018) Two novel protein O-glucosyltransferases that modify sites distinct from POGLUT1 and affect Notch trafficking and signaling. Proc. Natl. Acad. Sci. USA, 115, E8395–E8402.
11) Schneider, M., Kumar, V., Nordstrøm, L.U., Feng, L., Takeuchi, H., Hao, H., Luca, V.C., Garcia, K.C., Stanley, P., Wu, P., et al. (2018) Inhibition of Delta-induced Notch signaling using fucose analogs. Nat. Chem. Biol., 14, 65–71.
12) Colley, K.J., Kitajima, K., & Sato, C. (2014) Polysialic acid: Biosynthesis, novel functions and applications. Crit. Rev. Biochem. Mol. Biol., 49, 498–532.
13) Thompson, M.G., Foley, D.A., Swartzentruber, K.G., & Colley, K.J. (2011) Sequences at the interface of the fifth immunoglobulin domain and first fibronectin type III repeat of the neural cell adhesion molecule are critical for its polysialylation. J. Biol. Chem., 286, 4525–4534.
14) Mendiratta, S.S., Sekulic, N., Lavie, A., & Colley, K.J. (2005) Specific amino acids in the first fibronectin type III repeat of the neural cell adhesion molecule play a role in its recognition and polysialylation by the polysialyltransferase ST8Sia IV/PST. J. Biol. Chem., 280, 32340–32348.
15) Bhide, G.P., Fernandes, N.R.J., & Colley, K.J. (2016) Sequence requirements for neuropilin-2 recognition by ST8SiaIV and polysialylation of its O-glycans. J. Biol. Chem., 291, 9444–9457.
16) Kanagawa, M. & Toda, T. (2017) Muscular dystrophy with ribitol-phosphate deficiency: A novel post-translational mechanism in dystroglycanopathy. J. Neuromuscul. Dis., 4, 1–9.
17) Hara, Y., Kanagawa, M., Kunz, S., Yoshida-moriguchi, T., Satz, J.S., Kobayashi, Y.M., Zhu, Z., Burden, S.J., Oldstone, M.B., & Campbell, K.P. (2011) Like-acetylglucosaminyltransferase (LARGE)-dependent modification of dystroglycan at Thr-317/319 is required for laminin binding and arenavirus infection. Proc. Natl. Acad. Sci. USA, 108, 17426–17431.
18) Kanagawa, M., Saito, F., Kunz, S., Yoshida-Moriguchi, T., Barresi, R., Kobayashi, Y.M., Muschler, J., Dumanski, J.P., Michele, D.E., Oldstone, M.B., et al. (2004) Molecular recognition by LARGE is essential for expression of functional dystroglycan. Cell, 117, 953–964.
19) Yagi, H., Nakagawa, N., Saito, T., Kiyonari, H., Abe, T., Toda, T., Wu, S.-W., Khoo, K.-H., Oka, S., & Kato, K. (2013) AGO61-dependent GlcNAc modification primes the formation of functional glycans on α-dystroglycan. Sci. Rep., 3, 1–9.
20) Kizuka, Y., Matsui, T., Takematsu, H., Kozutsumi, Y., Kawasaki, T., & Oka, S. (2006) Physical and functional association of glucuronyltransferases and sulfotransferase involved in HNK-1 biosynthesis. J. Biol. Chem., 281, 13644–13651.
21) Bonnon, C.,Bel, C., Goutebroze, L.,Maigret, B.,Girault, J.-A. & Faivre-Sarrailh, C. (2007) PGY repeats and N-glycans govern the trafficking of paranodin and its selective association with contactin and neurofascin-155. Mol. Biol. Cell, 18, 229–241.
22) Yano, H., Yamamoto-hino, M., Abe, M., Kuwahara, R., Haraguchi, S., Kusaka, I., Awano, W., Kinoshita-toyoda, A., Toyoda, H., & Goto, S. (2005) Distinct functional units of the Golgi complex in Drosophila cells. Proc. Natl. Acad. Sci. USA, 102, 13467–13472.
23) Yagi, H., Takakura, D., Roumenina, L.T., Fridman, W.H., Sautes-Fridman, C., Kawasaki, N., & Kato, K. (2018) Site-specific N-glycosylation analysis of soluble Fcγ receptor IIIb in human serum. Sci. Rep., 8, 2719.
24) Kawasaki, N., Okumoto, T., Yamaguchi, Y., Takahashi, N., Fridman, W.H., Sautès-Fridman, C., Yagi, H., & Kato, K. (2014) Site-specific classification of N-linked oligosaccharides of the extracellular regions of Fcγ receptor IIIb expressed in baby hamster kidney cells. J. Glycomics Lipidomics, 4, 1000116.
25) Nagae, M., Kizuka, Y., Mihara, E., Kitago, Y., Hanashima, S., Ito, Y., Takagi, J., Taniguchi, N., & Yamaguchi, Y. (2018) Structure and mechanism of cancer-associated N-acetylglucosaminyltransferase-V. Nat. Commun., 9, 3380.
26) Suzuki, N. & Lee, Y.C. (2004) Site-specific N-glycosylation of chicken serum IgG. Glycobiology, 14, 275–292.
27) Kondo, S., Yagi, H., Kamiya, Y., Ito, A., Kuhara, M., Kudoh, A., & Kato, N.T. and K. (2012) N-glycosylation profiles of chicken immunoglobulin Y glycoproteins expressed by different production vehicles. J. Glycomics Lipidomics, 5, 1–5.
28) Arnold, J.N., Radcliffe, C.M., Wormald, M.R., Royle, L., Harvey, D.J., Crispin, M., Dwek, R.A., Sim, R.B., & Rudd, P.M. (2004) The glycosylation of human serum IgD and IgE and the accessibility of identified oligomannose structures for interaction with mannan-binding lectin. J. Immunol., 173, 6831–6840.
29) Noro, E., Togayachi, A., Sato, T., Tomioka, A., Fujita, M., Sukegawa, M., Suzuki, N., Kaji, H., & Narimatsu, H. (2015) Large-scale identification of N-glycan glycoproteins carrying lewis x and site-specific N-glycan alterations in fut9 knockout mice. J. Proteome Res., 14, 3823–3834.
30) Yagi, H. & Kato, K. (2017) Functional roles of glycoconjugates in the maintenance of stemness and differentiation process of neural stem cells. Glycoconj. J., 34, 757–763.
著者紹介Author Profile
 矢木 宏和(やぎ ひろかず)
矢木 宏和(やぎ ひろかず)名古屋市立大学大学院薬学研究科講師.博士(薬学).
略歴1979年香川県に生る.2003年名古屋市立大学薬学部卒業.08年同大学院薬学研究科博士後期課程修了.その後,同研究科および自然科学研究機構生理学研究所において日本学術振興会特別研究員.09年より名古屋市立大学大学院薬学研究科助教を経て,13年より現職.
研究テーマと抱負核酸やタンパク質と異なり,分岐性,構造異性,不均一性といった特徴を有する糖鎖がいかにして生命現象を担っているかに興味を持ち研究を進めている.特に,近年はタンパク質特異的な糖鎖の発現機構の解明を目指した研究を行っている.
ウェブサイトhttp://www.phar.nagoya-cu.ac.jp/hp/sbk/
趣味息子と遊ぶこと.