アミノペプチダーゼは,タンパク質やペプチドのアミノ(N)末端から1~3個のアミノ酸を順次遊離するエキソペプチダーゼである.その酵素活性から,アミノペプチダーゼはタンパク質やペプチドのプロセシングに伴う活性化または不活性化を介して生体機能の調節に関与すると考えられてきた1).
筆者らは,1990年代の初頭に「(哺乳類)アミノペプチダーゼの構造と機能」をテーマとした研究を開始した.その過程で3種の酵素を新規にクローニングし,現在に至るまでその構造と機能に関する検討を継続している2, 3).これら3種の酵素[筆者らの当初の命名では胎盤性ロイシンアミノペプチダーゼ(placental leucine aminopeptidase:P-LAP),脂肪細胞由来ロイシンアミノペプチダーゼ(adipocyte-derived leucine aminopeptidase:A-LAP),白血球由来アルギニンアミノペプチダーゼ(leukocyte-derived arginine aminopeptidase:L-RAP]は,活性発現に亜鉛を必要とする一群の亜鉛結合型アミノペプチダーゼであるM1アミノペプチダーゼファミリー酵素に分類される4).ヒトM1アミノペプチダーゼファミリーは12種類の酵素からなっており,亜鉛結合モチーフであるHEXXH(X)18E配列,および(一部にバリエーションはあるが)ペプチド基質のN末端認識モチーフであるGXMEN配列を共通に持っている4, 5).
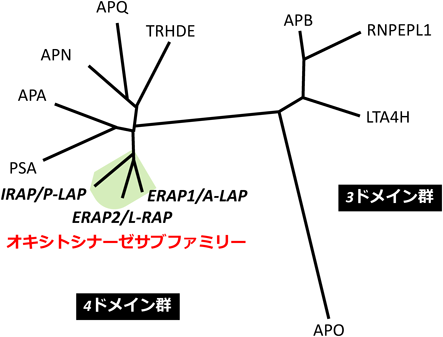
図1はヒトM1アミノペプチダーゼファミリーの系統樹を示している.同ファミリーは四つのドメイン構造を持つ群と三つを持つ群に大別される6).四つのドメイン構造を持つ群の中で,特にP-LAPとA-LAPの一次構造は分子全体にわたってよく保存されており(43%),他のM1酵素との相同性(17~30%)に比べて顕著に高い.またA-LAPとL-RAPの相同性も49%と高い.そこで筆者らはこれら3酵素についてM1ファミリーの中でも特に「オキシトシナーゼサブファミリー酵素」と呼ぶことを提唱し,現在広く使われる用語となっている(オキシトシナーゼサブファミリーの名称は筆者らが最初にクローニングしたP-LAPがオキシトシン分解酵素であることに由来する)2, 7).
現在までにM1アミノペプチダーゼファミリー酵素の実体が遺伝子クローニングを介して明らかになるとともに,これらが非常に多彩な生理的/病理的機能を発揮することが明らかになってきた.特に「オキシトシナーゼサブファミリー酵素」に関してはそのユニークな細胞内局在とともに疾患との関連が注目され,現在までに多くの知見が蓄積している.
本稿では興味深い機能が多く報告されている「オキシトシナーゼサブファミリー酵素」の中でも最も多くの研究成果が報告され,かつ筆者らがその展開に長く関与してきたA-LAPを主に取り上げ,これまでの成果をまとめてみたい.
なお,筆者らは当初脂肪細胞中に見いだされていたEST配列をプローブとしてクローニングしたことから当該酵素をA-LAPと命名した8).その後,筆者らを含めた多くの研究グループにより本酵素が通常細胞内の小胞体内腔に存在することが確認されたことから,現在本酵素は一般的に小胞体アミノペプチダーゼ(endoplasmic reticulum aminopeptidase 1:ERAP1)と呼ばれている9).同様に,主に小胞体内腔に存在するL-RAPは小胞体アミノペプチダーゼ2(ERAP2),さらにGLUT4含有細胞内小胞に存在し,インスリン刺激により細胞膜に輸送されることが示されたP-LAPはインスリン調節性アミノペプチダーゼ(IRAP)と呼ばれることの方が多い9).本稿においても以降の記述においては,より多く用いられる呼称(ERAP1, ERAP2およびIRAP)を主に用いることにする.
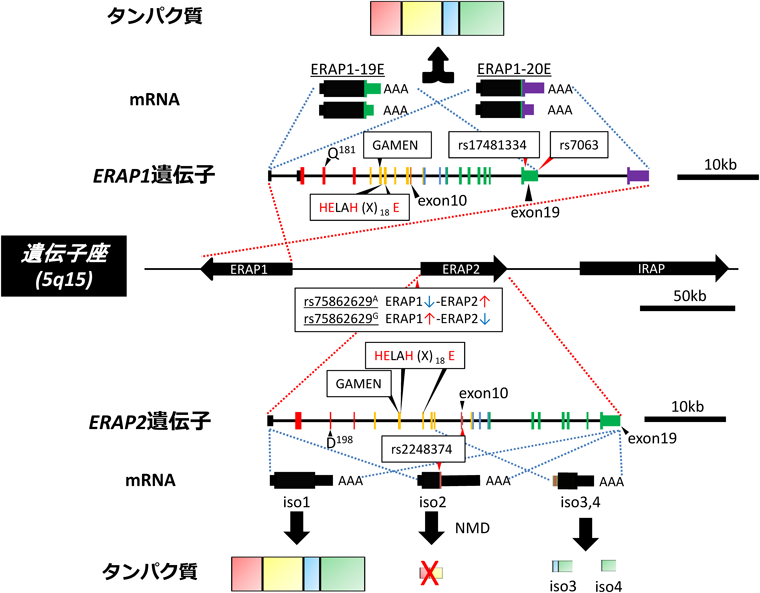
ヒトにおいては5番染色体長腕上の隣接した遺伝子座(5q15)に二つのERAP遺伝子(ERAP1, 2)が存在しており,同じくM1アミノペプチダーゼファミリーに属するIRAPとともに遺伝子クラスターを形成している(図2)10).またそれらのエクソン/イントロンの構成はHEXXH(X)18EおよびGXMEN配列のエクソン上の存在位置を含め,非常によく保存されている[GXMENおよびHEXXH配列はexon6, HEXXH(X)18Eの最後のEはexon7].したがって,三つの酵素は進化の過程で単一の遺伝子から重複して生じたものと考えられる(図2)2, 3).
ERAP1遺伝子は20個のエクソンから構成され,選択的スプライシングによって最終エクソンが異なる二つのアイソフォーム(ERAP1-19EおよびERAP1-20E)が形成される.ERAP1-19EおよびERAP1-20Eのアミノ酸配列上の違いは,C末端のそれぞれ2アミノ酸(RM)と8アミノ酸(DPEADATG)のみにある(図3A)10).またそれぞれのmRNAについて選択的3′末端形成により,長さの異なる二つの3′非翻訳領域(3′UTR)が存在することから,ERAP1には完全長のタンパク質をコードする少なくとも四つの転写物が存在していることになる(図2)8, 10).
最近,筆者らはHeLa細胞を用いた検討で,ERAP1-19Eはインターフェロンγ(IFN-γ)に応答してその発現量が増加するが,ERAP1-20Eはほとんど影響を受けないことを見いだした.この結果はIFN-γによる刺激が転写のみならずERAP1の選択的スプライシングにも影響を及ぼしている可能性を示すものであり,刺激への応答性の違いはそれぞれのアイソフォームの発現量の違いだけでなく,異なる機能への関与を示唆するものと考えられる11).
一方ERAP2遺伝子は19個のエクソンから構成されており,遺伝子構造や塩基配列においてERAP1と高い相同性を示すが,ERAP1においてみられたような選択的スプライシングは確認されていない12, 13).
ERAP1, 2は他のMHCクラスI関連因子と同様にIFN-γ, TNF-α, LPS等に応じて転写誘導されることが示されている14–19).この中でERAP2に関する筆者らのプロモーター解析では,転写因子IRF1やPU.1を介したIFN-γ応答メカニズムが示されている13).この他にERAP1ではNF-κBやp53による発現誘導が知られている17, 20).
ERAP1, 2の遺伝子領域には高度な一塩基多型(SNP)性が認められる(後述).その中で最近ERAP1, 2の機能や発現に影響を及ぼす興味深いSNPが報告された.すなわちERAP2のプロモーター領域上流に位置するrs75862629がAである場合はERAP2の発現が高い傾向になり,Gの場合は発現が低い傾向となる.一方でERAP1に関しては逆にAで発現が低く,Gの場合に発現が高い傾向になることが示されている.この結果は個別の転写誘導だけでなくERAP1とERAP2の発現バランスを調節する統括的制御機構の存在を示唆するものである21).またrs7063は,ERAP1-19E mRNAの3′UTR上に存在するポリ(A)付加配列(もしくはERAP1-20Eの19番目のイントロン)に位置するSNPで,アミノ酸配列上の変化は伴わないが,ヒト末梢血細胞においてrs7063AアリルからはERAP1-19E mRNAが優先的に産生される.その結果,ERAP1-19E/20E mRNAの比率が増加し,かつ全体的なERAP1タンパク質発現量も増加することが示されている22).
ERAP2について特筆すべきは約50%の頻度を示すSNPであるrs2248374の存在である23).rs2248374はERAP2のexon10直下のスプライシング部位に位置し,Aの場合は正常なスプライシングが生じて完全長のERAP2をコードするmRNAが形成されるのに対しGではスプライシングが抑制され,その下流に新たな終止コドンが出現することでC末端を欠失した短縮型ERAP2をコードするmRNAが形成される.このmRNAは,新たに生じた終止コドンが未成熟終止コドン(premature termination codon)として認識されることでmRNAの品質管理機構であるnon-sense mediated decay(NMD)の標的となり速やかに分解される.したがってrs2248374Gに由来するERAP2の発現量は著しく低下し,両アリルにG/Gを持つ個人は機能的ERAP2をほぼ完全に欠損することになる23).興味深いことに,このrs2248374Gアリルからは,インフルエンザウイルスの感染に応答してexon9付近を転写開始点とする二つの新たなERAP2アイソフォームが生成することが見いだされた24).この二つのアイソフォームはともにN末端欠失型で酵素活性部位を含まないが,ERAP1または2とヘテロ二量体を形成し,ドミナントネガティブとして機能する可能性が示唆されている24).このような顕著な発現抑制もしくは機能低下をもたらすSNPがハプロタイプの50%以上の頻度を占めるに至った理由については必ずしも明確ではないが,恒常的かつ過剰なERAP2活性は生体にとってむしろ有害に機能し,平常においては低い状態を維持する必要性があるのかもしれない(後述).
またERAP1, 2はMHCクラスI分子への抗原提示を介して免疫反応の発動に関与するが,一部のがん細胞やウイルス感染細胞でその発現が抑制されることが報告されている.その例として,ヒトサイトメガロウイルス(CMV)には主要なアイソフォームであるERAP1-19Eの発現を特異的に抑制するmiRNAとしてmiR-US4-1, miR-UL112-5pが知られている25, 26).このmiRNAを介した発現抑制の存在は,ウイルスがERAP1タンパク質の生成を抑制することで宿主の免疫監視機構を回避する戦略を用いていることを示唆している.またmiR-UL112-5pについて,その標的部位に位置するrs17481334がAの場合はERAP1の発現が抑制されるが,Gである場合に抑制されないことが示されており26),ERAP遺伝子の多型性が病原体との共進化により獲得された可能性を推測させる.加えて同定されているmiRNAの標的がともにERAP1-19Eであることは,二つのアイソフォームのうちERAP1-19Eがより免疫応答に重要である可能性を示している.この仮説はERAP1-19EがIFN-γに応答するアイソフォームであることを示した筆者らの結果11)とも矛盾しない.
以上のようにERAP1, 2の遺伝子発現は精緻に制御されている.このことはERAP活性の亢進/抑制のバランス調節が生体およびその集団にとって重要な課題であることを示唆している.
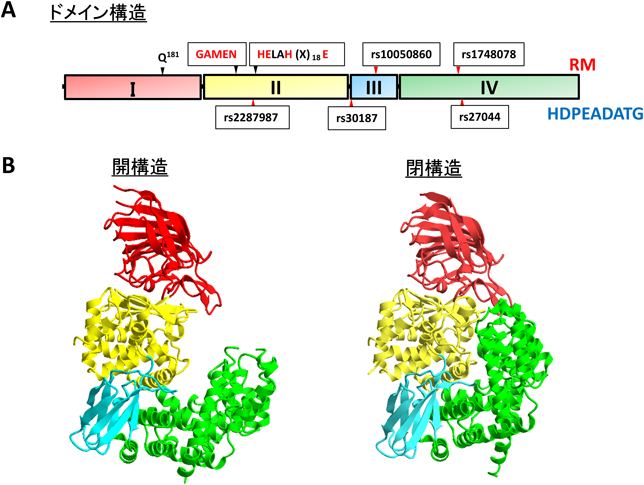
2011年に,アミノペプチダーゼの競合阻害剤であるベスタチンを結合したヒトERAP1の結晶構造が決定された27, 28).その結果,本酵素は活性中心への基質の侵入が妨げられる閉(close)構造と,基質の侵入を可能にする開(open)構造(図3B)の二つの異なる立体構造をとることが示された.同じくオキシトシナーゼサブファミリーに属するERAP2, IRAPの立体構造も決定されている29–33).両酵素とも開構造と閉構造の両方をとりうると考えられているが,現時点で得られているのはERAP2では閉構造のみ,IRAPでは閉構造と開構造の中間状態である半開(semi-open)構造のみである.
ERAP1はドメインI(Met1~Phe254),ドメインII(Glu255~Gln527),ドメインIII(Lys528~Glu613),ドメインIV(Asp614~Met941)と四つのドメイン構造から構成されており(図3A),ERAP2, IRAPも同様に四つのドメインで構成される.ドメインIには基質特異性を決める残基として筆者らが同定したGln181が存在し34),活性部位(S1サイト)を形成することで基質のN末端に結合部位を提供する.ドメインIIは他のM1ファミリー酵素と同様にテルモリシン様のαβ構造を持ち,触媒中心[G317XMEN321,H353EXXH(X)18E376]が存在する.ドメインIIIはβサンドウィッチを形成する二つのβシートからなる小さなドメインであり,ドメインIIとIV間にはさまれることにより,そこに大きなくぼみが形成されている.ドメインIVは酵素活性を調節しており,ドメインIIから離れることにより,活性中心への基質のアクセスを可能にする.ドメインIVは,M1ファミリーの中でも最も多様性に富んだ領域である.ERAP1のドメインIVは16個のさまざまな長さのヘリックスから構成され,活性部位に面するくぼみを形成するように組み立てられている27, 28).
ERAP1の開閉した結晶構造を比較してみると,その主な立体構造の変化はドメインIVの移動に起因する27, 28).ここで重要となるアミノ酸残基はTyr455であり,開構造では活性部位から離れることが見いだされており,Tyr455のPheへの置換は人工基質の加水分解する速度を1/200まで減少させる27).
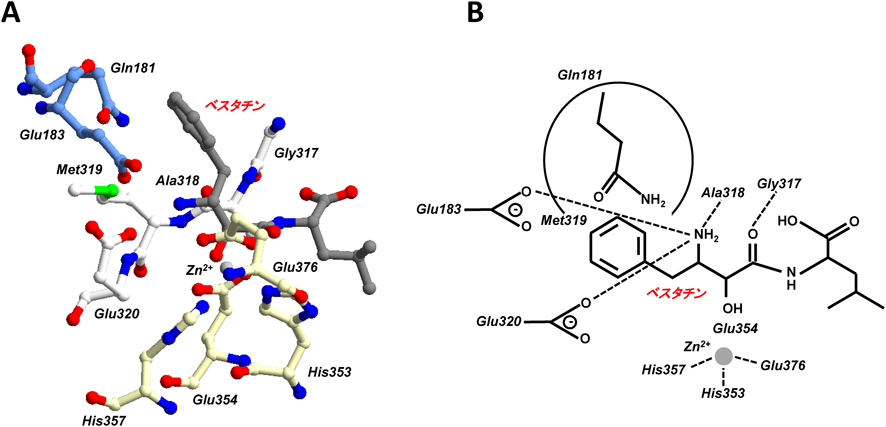
ERAP1の活性部位は,Gln181を含むドメインIのD1ループ,HEXXH(X)18EモチーフのHEXXHとEがそれぞれ含まれるドメインIIのヘリックスH2, H3, H5およびGAMENループの五つの二次構造により構成されている27).さらに,阻害剤であるベスタチンと結合した場合,亜鉛原子はHEXXHヘリックスのHis353とHis357,隣接するヘリックスのGlu376によって配位され,Glu354はベスタチンのα-ヒドロキシ基の隣に配置される.ベスタチンのN末端は,ドメインIループのGlu183およびGAMENモチーフのGlu320と相互作用している.さらに,GAMENモチーフのGly317とAla318は,それぞれベスタチンのカルボキシレート基とアミド基と相互作用する.Met319はS1ポケットの一部を形成し,Glu320のカルボキシ基はベスタチンのN末端と相互作用する27).これらの活性中心の残基と基質との相互作用により酵素活性が発現すると考えられる(図4).
オキシトシナーゼサブファミリー酵素は,MHCクラスIに提示するための抗原ペプチドを生成する役割を持っており,また後述するように,ERAP1は強直性脊椎炎,ベーチェット病,乾癬,ERAP2はクローン病,バードドショット脈絡網膜症などの自己免疫疾患/炎症性疾患のリスク因子になっていることから,治療薬となりうる阻害剤の研究が進んでいる.Węglarz-Tomczakらは,ホスホン酸およびホスフィン酸のスクリーニングから,特にホスフィン酸ジペプチド類似体がERAP1よりもERAP2の強力な阻害剤となることを示した35).Kokkalaらは,phosphinic pseudotripeptidesがERAP1, ERAP2, IRAPの強力な阻害剤となりうることを示し36),P1′およびP2′部位に存在するアミノ酸残基が酵素活性および基質特異性において重要であることを示した.このようなリガンドを用いたスクリーニングは,本酵素群のMHCクラスIによって提示されるペプチドのプロセシング,およびヒト疾患における特定のアミノペプチダーゼの役割を解明するための非常に重要な手段となっている.
アミノペプチダーゼ活性は,アミノ酸に発色団をつけた人工基質により容易に測定できる.人工基質を用いた結果を比較すると,各M1ファミリー酵素がそれぞれに特徴的な基質特異性を有することがみてとれる37, 38).オキシトシナーゼサブファミリー酵素に関しても,82種類のアミノ酸または修飾アミノ酸をつけた人工基質を用いて基質特異性が比較された39).その結果,ERAP1は主に疎水性アミノ酸および芳香族アミノ酸を遊離するのに対し,ERAP2は塩基性アミノ酸を選好することが示されている.さらに興味深いことに,IRAPはオキシトシン分解酵素であることを反映してベンジルシステインに作用するとともに,他の基質に対する反応性はERAP1とERAP2の反応性をあたかも足し合わせたようなものになっている.このことは,ERAP1とERAP2は小胞体内腔に共局在して協調的に機能しているのに対し,IRAPは特定の細胞内小胞に存在して単独で作用していることを反映したものと思われている.系統樹からも,ERAP1とERAP2はIRAPから分岐しており,進化の過程における両者の機能分離が推察される(図1)3).
筆者らは,人工基質への反応性を指標にしてERAP1の基質特異性を決定する残基としてGln181を同定した34).本残基をERAP2の対応する残基であるアスパラギン酸(Asp)に変換すると,人工基質に対する特異性が疎水性アミノ酸からERAP2と同じ塩基性アミノ酸へと劇的に変化した.本残基に相当する位置にあるアミノ酸は,アミノペプチダーゼA(APA)やアミノペプチダーゼB(APB)においてもその基質特異性を決めているとともに,それぞれの酵素のカルシウムイオンや塩化物イオンの要求性にも変化をもたらすことが示された37, 40).したがって,ERAP1のGln181およびM1酵素における対応位置に存在するアミノ酸は,各酵素の基質特異性およびイオン要求性を決める重要な役割を果たしていると考えられる.実際,X線結晶解析の結果はGln181がS1ポケットに存在し,ペプチド基質のP1アミノ酸残基と相互作用していることを示しており,この相互作用の存否が各ペプチドの基質としての反応性を決める要因の一つと考えられる27).
人工基質を用いた検討はアミノペプチダーゼの酵素活性に関し多くの情報をもたらすが,必ずしも実際のペプチド基質への反応性を忠実に反映するものではないことを,筆者らはたびたび経験している.一例として,筆者らはERAP1の天然基質となりうる各種のペプチドホルモンについて検討した.その結果,アンギオテンシン(Ang)II(DRVYIHPF)およびカリジン(KRPPGFSPFR)が良好な基質となることが認められたが,両ペプチドのN末端アミノ酸は荷電を有しており,人工基質の検討からは予想しにくいことであった41).このことは,酵素活性を介したM1各酵素の酵素学的/機能的解析には,実際のペプチド基質を用いた検討が不可欠であることを示している42).
ERAP1, 2のペプチド基質に対する酵素活性の解析は,両酵素が小胞体内腔における抗原提示機構において重要な役割を果たすことから,MHCクラスI分子に結合する抗原ペプチドのトリミング(プロセシング)を念頭に多くの検討がなされてきた13, 43).その結果,ERAP1は抗原提示に適した性状を有していることが明らかになっている.すなわち,1)アミノ酸鎖の長さが9~16のペプチドに選好性があり,これより短いペプチドおよび長いペプチドに対する作用は弱いこと27, 44, 45),2)同じN末端側配列を有するペプチド(たとえばRYWANATRSX)で比較すると,C末端のXが疎水性アミノ酸であることがN末端アミノ酸のトリミングに有利であること27, 45),3)ペプチドの中間配列内のアミノ酸(たとえばLYWANATXSG)に対しても特定のアミノ酸残基に対して選好性を示すこと46)などがあげられる.これらの性質は,抗原ペプチドがMHCクラスI分子に結合する際に必要とされる性状(適切な長さ,C末端の疎水性,中間配列中の特定部位でのMHCクラスI分子との相互作用)をトリミングされた抗原ペプチドに付与するために獲得したものと考えられる47).このようなトリミング酵素に好都合な性状はERAP2には認められていない.
ERAP1による抗原ペプチドのN末端アミノ酸のトリミングには,上記のように適当な長さを持ち,疎水性のC末端を持っているペプチドが有利である.これらを踏まえるとERAP1分子上にはN末端を認識する反応部位とC末端を認識するペプチド結合部位の存在が想定され,両部位の距離が酵素によってトリミングされるペプチドの長さを規定するとする「分子ルーラーモデル」が提唱されている27, 45).実際,X線結晶解析の結果,ERAP1には存在し,ERAP2にはないC末端認識部位と想定される構造が同定されている29).これらのことから抗原提示において主要な役割を果たすトリミング酵素は,抗原ペプチドの生成において好都合な性質を有するERAP1であり,ERAP2は補助的な作用にとどまると考えられている(ちなみにERAP2は遺伝子そのものがマウスでは欠損しており,ヒトにおいてもrs2248374の影響で一定の割合の個人で欠損していることからもERAP1の重要性が垣間みえる).
ERAP1, 2の組換え型タンパク質は,ゲルろ過,超遠心分析により単量体であるとされている41, 48).しかし,両酵素が小胞体内腔に共局在している細胞では,一部の単量体酵素どうしがヘテロ二量体を形成し,単独で作用する場合に比べ効率的/協調的に成熟抗原を生成することが報告されている48, 49).両酵素間の親和性が低いとされるため二量体形成が実際に起こっているかいまだ検討の余地があるが,強制的にヘテロ二量体化した酵素タンパク質を用いて,両者による協調的かつ効率的なトリミングが確認されている50, 51).このとき形成されたヘテロ二量体の基質特異性はERAP1のそれに近いことから,ERAP2の役割は結合したERAP1にコンホメーション変化を誘導し,その作用を高めることにあるとされており,主要なトリミング酵素はやはりERAP1であると思われる.
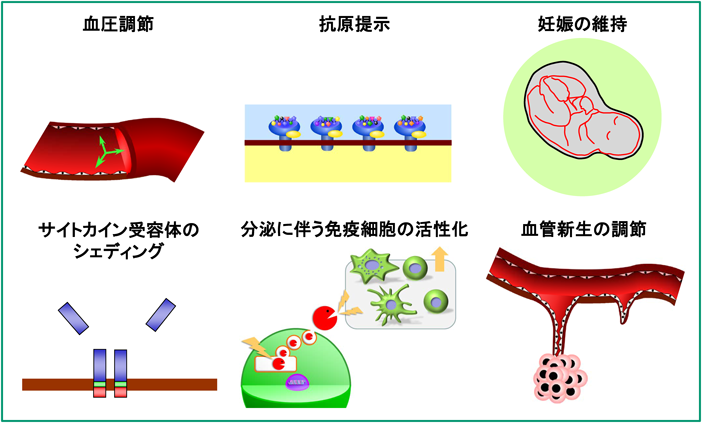
現在,ERAP1は多様な生理的/病理的機能を有する多機能酵素であることが広く認識されている(図5).
本酵素がクローニングされた当初から各研究グループが独自にERAP1の機能研究を行ってきた結果として,本酵素には多くの別名がある.筆者らは,本酵素をEST配列の由来および人工基質に対する反応性からA-LAPと命名し報告した8).このときの検討ではAng IIとカリジンが天然由来ペプチドの中で最もよい基質であったことから,本酵素が血圧調節に関与する可能性が考えられた.その後,本酵素のSNP解析によりrs30187(K528R)が本態性高血圧症の発症に関連していることが報告され,実際,筆者らはR528変異体が野生型(K528)と比べ,Ang II分解活性およびブラジキニン生成活性が低いことを示した.したがって,酵素活性の低いR528変異体を発現している個体は,より高い血圧傾向を示すものと考えられた52, 53).さらに,細胞外に分泌されたERAP1がAng IIの分解を介して血圧の調節に関与していることが最近報告されており54),また広範囲のSNP解析においてもERAP1, 2がともに血圧調節に関係していることを示す報告がある55).
Schomburgらはラットオルソログをクローニングし,PILS-AP(puromycin-insensitive leucyl-specific aminopeptidase)と命名した56).さらにSatoらのグループは,PILS-APが血管内皮細胞の分化に伴って発現し,VEGFによりその発現が亢進されることを報告した.PILS-APの発現を特異的に抑制するとVEGFによる血管新生が阻害されることなどから,PILS-APが血管新生を促進すると考えられている57–60).一方,Mizutaniらのグループは,ERAP1を過剰発現させた子宮内膜がん細胞においてAng IIによるVEGFの発現誘導が低下することを見いだしており,ERAP1が血管新生にはむしろ抑制的に作用していると報告している61).このような血管新生に対する作用の相違は,ERAP1が機能を発揮している“場(細胞内か細胞外か)”が異なることによる反応メカニズムの違いによると思われる.
一方,Cuiらは,ヒト細胞での検討で本酵素がTNFR1, IL-6Rα, IL-1RIIなどの炎症性サイトカイン受容体の切断・遊離(シェディング)に関与することを見いだし,本酵素をARTS-1(aminopeptidase regulator of TNFR1 shedding)と命名した62–65).受容体のシェディングによりサイトカインの情報伝達効率が変化することが考えられることから,ARTS-1は炎症反応および自然免疫系の調節等生体防御反応において重要な役割を果たすことが想定されている.しかし,ERAP1欠損マウスの血中受容体量は正常マウスと有意な差が認められておらず,シェディング機能の意義を疑問視する報告もある66).
さらに,Shastriらのグループはマウスで14),Goldbergらのグループはヒトで15)本酵素が小胞体に局在し,MHCクラスI分子への抗原提示に重要であることを報告した.ちなみにShastriらは,ERAP1のマウスオルソログをERAAP(endoplasmic reticulum aminopeptidase associated with antigen presentation)と命名している.
以上のようにERAP1は多くの興味深い機能を発揮することが報告されているが,これらの機能が発現されるべき“場”がそれぞれ異なっている.したがってERAP1が存在すべき場所が機能ごとに異なることになる(すなわち,A-LAPは細胞外液41),PILS-APは細胞質56, 57),ARTS-1は細胞膜62),ERAP1/ERAAPは小胞体内腔14, 15)).
ERAP1の機能で最も広くかつ深く研究されているものは小胞体内腔における機能である.筆者らはクローニングした当初,ERAP1が膜画分にではなく,可溶性画分に分画されたことから本酵素を細胞質タンパク質と考えた8).しかしその後,ERAP1をCOS-7細胞に過剰発現させると培養液中に分泌されることを観察したため,あらためて本酵素を分泌酵素として報告した41).ところがHeLa細胞は本酵素を発現しているにもかかわらず,培養液中に本酵素が見いだされることはなかった.そこで細胞画分をより細かく検討した結果,本酵素が(小胞体貯留シグナルを持たないにもかかわらず)小胞体内腔にとどまっていることを最終的に確認した15).それまでに,小胞体内腔には抗原提示に関わるアミノペプチダーゼの存在が想定されていたことから,本酵素の抗原ペプチドプロセシング酵素としての検討をGoldbergらのグループとの共同研究として行った.一方,Shastriらのグループは同時期にマウス細胞の小胞体内腔におけるプロセシング酵素としてERAAPを同定していた.
MHCクラスI分子への抗原提示は,ウイルス抗原やがん抗原また自己免疫疾患の原因となる自己抗原など内在性タンパク質抗原の,細胞質におけるプロテアソームによる分解から始まる.分解されたペプチドの一部は小胞体に存在するペプチド輸送体(transporters associated with antigen processing:TAP)により小胞体内腔に輸送され,ERAP1, 2によりプロセシングされたのちMHCクラスI分子に結合して細胞表面に輸送され,細胞傷害性(CD8+)T細胞やNK細胞などの免疫担当細胞による認識の対象となる67).したがって,ERAP1の欠損または機能変化は,ウイルス・がん免疫の発動および自己免疫疾患の発症に影響する.このように,MHCクラスI分子への抗原提示はCD8+T細胞およびNK細胞の活性化に不可欠な過程であることは周知の事実であり,それに結合する抗原ペプチドの最終プロセシング酵素であるERAP1が自然免疫系および獲得免疫系において重要であることは容易に推測される.その結果,ERAP1, 2は免疫研究者の注目されるところとなり,現在までに多くの成果が積み上げられてきた.
ERAP1(またはERAAP)の機能研究では多くの場合,ERAP1発現細胞とノックアウト(またはノックダウン)細胞とが比較されており,1)ノックアウト細胞ではMHCクラスI分子の発現量が全般的に減少していること68),2)ノックアウト細胞ではペプチド抗原と結合したMHCクラスI分子の安定性が全般的に低下していること69),3)ERAP1が欠損することでMHCクラスI分子に提示される抗原ペプチドの組成が変化し,特定のウイルス感染細胞やがん細胞に対するCD8+T細胞の応答性が変化すること47, 69, 70),4)ERAP1の欠損による抗原ペプチドの組成変化で,ノックアウト細胞のMHCクラスI分子に,通常の細胞には生成されないペプチドが提示される結果,ノックアウト細胞はin vivoにおいて高い免疫原性を示すこと71),5)ERAP1をノックアウトしたがん細胞はNK細胞に対する感受性が高いこと72),さらに6)ノックアウトマウスにおいてはNK細胞の成熟・活性化レベルが亢進しており,炎症性サイトカインによるIFN-γの産生も高まっていること73),などが報告されている.これらの報告にある事象はERAP1がMHCクラスI分子に結合する抗原ペプチドの生成・成熟に関与することで起こるものである.ERAP1の欠損(または活性変化)によりMHCクラスI分子に結合する抗原ペプチドの組成が変化することで,認識される抗原ペプチドのヒエラルキーが変化する.その結果,欠損細胞において誘導される免疫応答は(正常)発現細胞におけるそれとは異なることになると考えられる.またERAP2についてもその機能の検討が行われ,1)MHCクラスI分子の発現量24),2)絨毛がんに対するNK細胞の活性化74),3)HIVに対する抵抗性75, 76)への関与など,ERAP1と同様の機能が報告されている.
以上のように,小胞体内腔に局在するERAP1, 2はMHCクラスI分子に提示される内因性抗原ペプチドの最終プロセシング酵素として協調して機能しており,細胞傷害性T細胞およびNK細胞の機能調節を介した生体防御反応において重要な役割を果たしている.抗原ペプチドのプロセシング過程については,抗原ペプチド前駆体が小胞体内腔においてMHCクラスI分子に結合した状態でトリミングされるのか(MHCクラスIテンプレートモデル),遊離した状態でトリミングされるのか(分子ルーラーモデル),その反応様式についても検討されており,現在のところ,抗原ペプチド前駆体のMHCクラスI分子に対する親和性の違いにより両方の様式が使い分けられると考えられている77, 78).
ERAP1, 2はMHCクラスI分子と同様,多型性に富んだタンパク質である.ERAP1のSNPと疾患との関わりで最初に報告された本態性高血圧症では当初rs30187(K528R)のみがリスク因子として同定されたが52),強直性脊椎炎(ankylosing spondylitis:AS)のリスク因子としては5か所のSNPが同定されている[すなわちrs2287987(M349V),rs30187(K528R),rs10050860(D575N),rs1782078(R725Q),rs27044(Q730E)]77).ここでrs30187/rs10050860の組合わせに着目すると,ERAP1の人工基質に対する酵素活性は相加的であり,両変異体の高活性型,低活性型の組合わせによりK528/N575>R528/N575>K528/D575>R528/D575の順になり,酵素活性に大きな差が生じることになる79).また個々の細胞または個人では,ERAP1は異なるアミノ酸がランダムに組み合わされるのではなく,特定のSNPの組合わせを保持した13種類のハプロタイプに分類される.これらは,抗原ペプチドのトリミング活性に基づき1)normal, 2)hypo, 3)hyperの3種のカテゴリーに大別される80, 81).したがって,本酵素の機能を検討する際には,個体レベルでハプロタイプの組合わせが異なることによる表現型の相違に留意することが求められている.
筆者らは,1)ERAP1には小胞体貯留シグナルが存在しない8)こと,および2)COS-7細胞に過剰発現させると細胞外へ分泌される41)ことから,本酵素は本来分泌されるべき酵素ではないかと考え検討した.その結果,1)マクロファージをLPSとIFN-γで処理し活性化すると,ERAP1の分泌が誘導されること,2)培養液にERAP1を添加したマクロファージでは,IgGで標識された被食分子に対する貪食が亢進されることを見いだした19, 82).さらに,活性化されたマクロファージをアミノペプチダーゼ阻害剤で処理すると,食作用活性化が部分的に抑制されたことなどから,細胞外のERAP1は活性化マクロファージによるオプソニン依存性貪食の少なくとも一部を担うことが明らかとなった.
次に筆者らは,分泌ERAP1がマクロファージの一酸化窒素(NO)合成にも寄与しうることを示した83).N末端がアルギニンであるAng III存在下において,アルギニン欠損培地で培養したERAP1遺伝子欠損マクロファージをLPS/IFN-γで活性化すると,野生型マクロファージに比べて低いNO合成活性しか示さなかった.一方,アルギニンを含む培地を利用した同様の解析では,野生型とERAP1遺伝子欠損マウスのNO合成量に違いは認められなかった.したがって,ERAP1はAng IIIの分解を介して(すなわち基質ペプチド依存的に)マクロファージに遊離アルギニンを供給する役割を担っており,本酵素によって産生された遊離アルギニンはマクロファージが産生するNOの原料として利用されうることになる.
さらに筆者らは,in vivoにおいてもERAP1がNO合成に重要であることを確認した84).すなわち,LPSを腹腔内投与したC57BL/6マウスでは急性期に血中におけるERAP1とNOが急増した.一方,同じ処理をしたERAP1遺伝子欠損マウスの血中NO濃度は野生型マウスに比べて半分程度であった.したがって,分泌されたERAP1は血中のNO産生を亢進し,結果として血圧調節に関与することが考えられた.これまでに,「細胞内には一酸化窒素合成酵素(NOS)による消費を考慮に入れても,十分なアルギニンが貯蔵されているにもかかわらず,細胞外のアルギニンが欠乏するとNOは合成されない」ことが知られていた(この現象は“アルギニンパラドックス”と呼ばれている85)).筆者らの結果は,分泌型ERAP1がN末端にアルギニンを有するペプチドへの作用を介して細胞外(血中)にアルギニンを供給しうることを示しており,ERAP1によって“アルギニンパラドックス”が回避される可能性があると考えられる.
最近,筆者らは細胞外のERAP1の一部が,LPSとIFN-γで活性化されたマクロファージから分泌される分泌小胞であるエクソソームと相互作用することを見いだした86).このERAP1結合エクソソーム(AP-exosome)には,いくつかのサイトカイン/ケモカインが含まれており,エクソソーム自体も活性化された状態にあると考えられる.そのうちCCL3とERAP1がエクソソームに高い貪食亢進活性を付与しており,さらに微量のIFN-βが共存する場合,エクソソームのTNF-αとIFN-γがマクロファージの誘導型(i)NOSの発現を強力に促進すると同時に,ERAP1がアルギニンの産生を担うことでNO産生を亢進させることが示された.これらの結果は,AP-exosomeがERAP1や複数のサイトカイン/ケモカインをコンパクトに搭載し,炎症性反応を多面的かつ効率的に制御しうる分子装置であることを示している.
一方,Aldhamenらは分泌ERAP1が自然免疫から獲得免疫にわたる幅広い免疫の賦活化に寄与することを報告した87).すなわち,細胞外ERAP1がヒト末梢血単核細胞の炎症性サイトカイン(TNF-α, IL-1β, IL-6)産生を増加させることや,NK細胞やT細胞,樹状細胞上の活性化マーカー(CD107α, CD69, CD80)量を増加させる.いずれの場合においてもマクロファージの活性化の場合と同様,ERAP1の酵素活性が必要である.彼らの結果を踏まえると,ERAP1はNK細胞の機能を1)小胞体内腔における抗原ペプチドレパートリーの創出,および2)分泌後におけるNK細胞の表面構造との直接的な相互作用,または培養液中に存在するペプチドのプロセシングを介して二重に調節できることになる.このように,ERAP1は存在する“場”の違いによりNK細胞の活性を正にも負にも調節することが可能である.
細胞外ERAP1は免疫反応だけではなく,その他の生命現象にも寄与しうる.組換え型ERAP1を用いた基質ペプチド探索の結果,本酵素は血圧調節性ペプチド(Ang群やカリジンなど)や神経ペプチド(ダイノルフィンやエンケファリン)などさまざまなペプチドに対して分解活性を示すことがわかっている41).(筆者らの見解では)ERAP1は通常小胞体内腔に局在していることから,上記生理活性ペプチドが実際に本酵素の生体内基質となりうるか否かについては長らく不明であった.分泌現象の発見により,本酵素が上記ペプチド群と細胞外で接触する可能性が示されたことになる.最近Hisatsuneらは1)小胞体貯留タンパク質であるERp44を欠損させたマウスでERAP1の分泌が促進されること,および2)ERp44遺伝子の欠損により,血中Ang IIの分解が促進され,収縮期血圧が低下することを報告している54, 88).このように,分泌されたERAP1はAng IIに代表される細胞外の機能性ペプチド/ホルモンの代謝を介して生体の恒常性維持に寄与することができると考えられる.そして,このことが本酵素に多機能性をもたらす一因になっているのかもしれない.
ERAP1の分泌は,LPS[Toll様受容体(TLR)4リガンド]だけでなく,TLR1/2や2/6, 9リガンドなどでも認められる82).ERAP1の分泌を誘導するTLRはいずれもMyD88をアダプターとする受容体であり,MyD88非依存性のTLR3のリガンドではERAP1の分泌はほとんど誘導されなかった.さらに,MyD88遺伝子欠損マウス由来の腹腔内マクロファージをLPSで刺激してもERAP1の分泌は観察されなかった.このERAP1分泌はTNF-αとIFN-βの阻害剤や中和抗体の処理または遺伝子のノックアウトなどにより著しく抑制されたため,受容体の下流に位置するサイトカインのネットワークが分泌に関与していると考えられる.
さらに筆者らは,ERAP1のexon10領域の欠失変異体ERAP1を発現させた細胞では,変異体が小胞体にとどまらず構成的に分泌されることを示した89).この結果は,ERAP1が本領域を介して小胞体内腔に貯留されていることを示唆する.実際,Hisatsuneらは,exon10領域に存在するCys487とERp44のチオレドキシン様ドメインに存在するCys29間におけるジスルフィド結合が,ERAP1の小胞体内腔への局在に重要であることを示し,小胞体内腔の酸化還元状態がERAP1の分泌を制御していると結論している54).ERAP1のexon10に相当するエクソンは,同じく小胞体貯留型のERAP2には存在する(ただし配列の類似性には乏しい)が,膜結合型のIRAPにはない89).また,exon10はエクソソームとの相互作用にも関与することを筆者らは報告している86).exon10を含む領域はX線結晶解析ではランダムな構造をして酵素表面に露出しており,さまざまなモノと相互作用することが可能な領域と考えられる30)(実際,exon10配列は酵素活性の発現には関与しておらず,この配列を欠損させた変異体は野生型に比べ若干高い人工基質加水分解活性を示す89)).
ERAP1の分泌に関し,Hisatsuneらは,敗血症のような全身性炎症状態に際し,ERAP1/ERp44複合体が小胞体内腔で形成され,ERAP1の分泌が抑制されることでAng IIの不活性化が抑制されると報告している.その結果,全身性炎症状態に伴う低血圧状態が改善されるとされている54).一方,筆者らは,炎症惹起物質の処理により分泌が促進されることで炎症性反応が促進されると考えている84).炎症状態が進行している生体内でのERAP1の挙動についてさらなる検討が必要と思われる.
以上のように初期検討の結果も含めると,ERAP1は非常に多様な生理的/病理的機能を有していることがわかる.それらのうち特定の機能発現には特定のタンパク質との相互作用/結合が必要であることが示されている62–64, 90–92).ERAP1はその結合パートナーに依存して存在部位を変化させることで,多様な活性を発揮しているのかもしれない.ERAP1が存在する“場”と多様な機能発現の関連は十分に検討されたとはいえず,ERAP1の存在部位を制御する機構の解明は今後に残された重要課題であろう.
MHCクラスI分子への抗原提示におけるERAP1の重要性から,SNPに基づく活性の変化は抗原ペプチドの質・量に影響することが容易に推測される.その結果,特定のMHCクラスI分子に結合する抗原ペプチドのレパートリーが変化し,それを認識するT細胞の応答が異なるようになり,がん・ウイルスに対する免疫応答が変化することが考えられる93, 94).また近年のgenome wide association studies(GWAS)によりERAP1の特定のSNP変異体をリスク因子とし,MHCクラスI分子とリンクした自己免疫疾患/炎症性疾患が数多く報告されている(表1)77, 95).以下に例をあげる.
表1 小胞体アミノペプチダーゼのSNPが関与する疾患 | 疾患名 |
|---|
| 強直性脊椎炎 | ベーチェット病 | 乾癬 | 溶血性尿毒症症候群 | 1型糖尿病 | C型肝炎 | 多発性硬化症 | 炎症性腸疾患 | 本態性高血圧症 | 妊娠中毒症/妊娠高血圧症 |
|---|
| ERAP1 リスクアリル | rs26653 | rs26653 | rs26653 | | | | | | | |
| (R127P) | (R127P) | (R127P) |
| | | | | rs26618 | | | rs26618 | |
| (I276M) | (I276M) |
| rs2287987 | rs2287987 | | | | | | | | |
| (M349V) | (M349V) |
| rs30187 | rs30187 | rs30187 | rs30187 | rs30187 | | rs30187 | rs30187 | rs30187 | |
| (K528R) | (K528R) | (K528R) | (K528R) | (K528R) | (K528R) | (K528R) | (K528R) |
| rs10050860 | rs10050860 | | | | | | | | |
| (D575N) | (D575N) |
| rs17482078 | rs17482078 | | | | | | | | |
| (R725Q) | (R725Q) |
| rs27044 | rs27044 | rs27044 | | | | | | | |
| (Q730E) | (Q730E) | (Q730E) |
| | rs151823 | | | | | | | |
| (intron variant) |
| | rs27043 | | | | | | | |
| (intron variant) |
| | rs27524 | | | | | | | |
| (intron variant) |
| ERAP2 リスクアリル | | | | | | rs2248374 | | | | rs2549782 |
| (ncRNA variant) | (N392K) |
| リスク因子となるバリアントを下線・太字で表示している. |
- 1) 強直性脊椎炎(AS)とは,背骨および骨盤を主として全身の腱や靱帯に原因不明の炎症が起こり,長い年月の中で「強直」して運動制限が生じる病気である.ASに最も強くリンクする遺伝子としてHLA-B*27が古くから(1973年)同定されている.しかし90%以上の患者がHLA-B*27陽性であるのに対し,HLA-B*27陽性のヒトのうち発症する割合はせいぜい5~10%程度であることから,強直性脊椎炎にリンクする他の遺伝子の存在が想定され,GWASによりERAP1の五つのSNP(既述)が同定された96, 97).抗原提示におけるMHCクラスI分子とERAP1の直線的な関係を考慮するとERAP1の各SNP変異体によりトリミングされた抗原ペプチドのHLA-B*27への結合性の違いが重要な発症要因であると考えられる.実際,本疾患の発症リスクはrs30187の高ペプチドトリミング活性型K528変異体が主要な役割を果たしており,高活性変異体酵素を発現する個体は低活性変異体発現個体に比べ3~4倍に発症リスクを高めている.
- 2) ベーチェット病(BD)は,1937年トルコの医師ベーチェットにより提唱された疾患であり,口腔粘膜の潰瘍,皮膚症状,眼のぶどう膜炎,外陰部潰瘍を主症状とする炎症性疾患である.BDもASと同様,特定のMHCクラスI分子(HLA-B*51)およびERAP1[rs10050860(D575N),rs1748078(R725Q)]SNP変異体が発症に深く関連している98).ASとは異なり,BDでは低ペプチドトリミング活性のSNP変異体がリスク因子になっている.特に特定のSNPの組合わせからなるERAP1*001(P127/V349/R528/N575/Q725/E730)はトリミング活性が低く,HLA-B*51に結合する抗原ペプチドのレパートリーを変化させるとともに,親和性の低いペプチドを生成することでHLA-B*51分子を不安定化し,結果としてBDの発症リスクを高めていると考えられている77, 78)(ちなみにASの発症リスクが高いSNPの組合わせはrs26653が加わってR127/M349/K528/D575/R725/Q730である77)).
- 3) 乾癬は表皮が過剰に増殖することによって引き起こされる慢性炎症性疾患であり,特定のMHCクラスI分子(HLA-C*06)とERAP1[rs30187(K528R),rs27044(Q730E)]が発症に関連している.ASと同様,高活性型変異体(K528/Q730)が主要なリスク因子になっている77, 99).
また,ERAP2に関してはアフリカ系アメリカ人の集団において,rs2549782と妊娠中毒症/妊娠高血圧症(preeclampsia)との関連が示されている100, 101).アフリカ系アメリカ人において,rs2549782はrs2248374(イントロン内SNP)と連鎖不均衡(LD)にあり,そのため高活性型であるN392変異体をコードするmRNAはNMDにより分解され,リスクアリルとされるrs2549782Gから産生される低活性型ERAP2(K392)のみが発現する.一方で上記のようなLD関係が認められないチリ人の集団では,ERAP2活性はアフリカ系アメリカ人の例と比較してよりバラエティに富むことになるはずであるが,rs2549782と妊娠中毒症の間に明確な関連は見いだされていない.したがって,rs2549782によるERAP2の活性の変化が直接的に妊娠中毒症/妊娠高血圧症の発症に関与するかどうかは今のところ明確でなく,他のLD関係にあるSNPsの関与も含めて,今後さらなる解析が求められる.その一方で多彩なERAP2活性を持つと推測されるチリ人の集団においても,rs2549782Tアレルのホモ接合体(ERAP2N392)は見いだされていない.その理由は不明であるが,胎盤におけるN392変異体の高発現はトロポブラストの免疫機能に影響を与えるため許容されないと考えられている74, 102).
さらに,ERAP1, 2の遺伝子型の違いにより酵素活性が大きく変化し,抗原ペプチドの組成が変化する結果,特定のハプロタイプを持つ集団は他に比べ統計的に有意な抗がん性または抗ウイルス性を示すことが報告されている103).ERAP1, 2遺伝子の多型性はさまざまな疾患に対応するために進化の過程で獲得した種の保存戦略であるのかもしれない.
本年(2019年)は筆者らがA-LAP/ERAP1のクローニングを報告してから20年目にあたる4).その間ERAP1, 2は筆者らの当初の予想をはるかに超えて,非常に多様な生理的/病理的機能を発揮するとともに多くの疾患にも関与することが明らかになった.特に自己免疫疾患/炎症性疾患との関連から,ERAP1, 2の酵素活性の調節による抗原ペプチドレパートリーの変化が病態の改善をもたらす可能性が考えられている.また最近では,ERAP1, 2がPD-1/PD-L1を標的とする免疫療法の治療効果に多大な影響を与えるタンパク質の一つであるとの報告もなされている104–106).ERAP1, 2を標的とした自己免疫疾患等に対するクスリの開発が期待される.
一方,これからの「オキシトシナーゼサブファミリー酵素」研究の方向性として,筆者らは脳内におけるそれらの機能に注目している.本稿では紹介できなかったが,IRAPは脳内に発現し学習や記憶の維持に重要であるとされ,そのメカニズムとして1)IRAPがAng IVの受容体として働くのか,あるいは2)IRAPの酵素活性による脳内ホルモンの代謝によるのか,が長く争点になっている107, 108).筆者らも最近1)P-LAP/IRAPが脳内におけるバソプレシン分泌の調節に関与していること,および2)ERAP1ノックアウトマウスの解析から本酵素が脳内セロトニンの生成に抑制的に関与することを見いだした.「オキシトシナーゼサブファミリー酵素」を介した脳機能研究の進展が求められる所以である.
謝辞Acknowledgments
本稿を終えるにあたり,妊婦血清に出現するなど興味深い動態を示すP-LAP/IRAPの存在を教えていただき,筆者(辻本)にアミノペプチダーゼ研究のきっかけを与えてくださった,水谷栄彦名誉教授(名大・医)に感謝します.また筆者らが理化学研究所在職中,本研究テーマを継続して実施することにご尽力くださった服部明(京大・薬),松本健(理研),小川健司(日大・生物資源),松本英子(埼玉医大),増田(青木)信奈子(東京女子医大),谷岡利裕(昭和大・薬),丸山正人(岡山大・薬)の元理化学研究所細胞生化学研究室所属の研究員各位に感謝します.
引用文献References
1) Taylor, A. (1993) Aminopeptidases: structure and function. FASEB J., 7, 290–298.
2) 服部明,辻本雅文(2003)オキシトシナーゼサブファミリー:M1アミノペプチダーゼの新しいサブファミリーの同定とその生理機能.生化学,75, 1225–1230.
3) Tsujimoto, M. & Hattori, A. (2005) The oxytocinase subfamily of M1 aminopeptidaess. Biochim. Biophys. Acta, 1751, 9–18.
4) 服部明,辻本雅文(2005)小胞体アミノペプチダーゼ:その発見とMHCクラスI抗原ペプチドプロセシングにおける役割.化学と生物,43, 739–745.
5) Maruyama, M., Arisaka, N., Goto, Y., Ohsawa, Y., Inoue, H., Fujiwara, H., Hattori, A., & Tsujimoto, M. (2009) Histidine 379 of human laeverin/aminopeptidase Q, a nonconserved residue within the exopeptidase motif, defines its distinctive enzymatic properties. J. Biol. Chem., 284, 34692–34702.
6) Cadel, S., Darmon, C., Pernier, J., Herve, G., & Foulon, T. (2015) The M1 family of vertebrate aminopeptidases: role of evolutionarily conserved tyrosines in the enzymatic mechanism of aminopeptidase B. Biochimie, 109, 67–77.
7) Rogi, T., Tsujimoto, M., Nakazato, H., Mizutani, S., & Tomoda, Y. (1996) Human placental leucine aminopeptidase/oxytocinase. A new member of type II membrane-spanning zinc metallopeptidase family. J. Biol. Chem., 271, 56–61.
8) Hattori, A., Matsumoto, H., Mizutani, S., & Tsujimoto, M. (1999) Molecular cloning of adipocyte-derived leucine aminopeptidase highly related to placental leucine aminopeptidase/oxytocinase. J. Biochem., 125, 931–938.
9) Hattori, A. & Tsujimoto, M. (2013) Endoplasmic reticulum aminopeptidases: biochemistry, physiology and pathology. J. Biochem., 154, 219–228.
10) Hattori, A., Matsumoto, K., Mizutani, S., & Tsujimoto, M. (2001) Genomic organization of the human adipocyte-derived leucine aminopeptidase gene and its relationship to the placental leucine aminopeptidase/oxytocinase gene. J. Biochem., 130, 235–241.
11) Aoki, K., Furuya, A., Matsumoto, K., & Tsujimoto, M. (2018) The gene expression of two endoplasmic reticulum aminopeptidase 1 isoforms is regulated by distinct posttranscriptional mechanisms. Biochem. Biophys. Res. Commun., 503, 3180–3185.
12) Tanioka, T., Hattori, A., Masuda, S., Nomura, Y., Nakayama, H., Mizutani, S., & Tsujimoto, M. (2003) Human leukocyte-derived arginine aminopeptidase. The third member of the oxytocinase subfamily of aminopeptidases. J. Biol. Chem., 278, 32275–32283.
13) Tanioka, T., Hattori, A., Mizutani, S., & Tsujimoto, M. (2005) Regulation of the human leukocyte-derived arginine aminopeptidase/endoplasmic reticulum-aminopeptidase 2 gene by interferon-γ. FEBS J., 272, 916–928.
14) Serwold, T., Gonzalez, F., Kim, J., Jacob, R., & Shastri, N. (2002) ERAAP customizes peptides for MHC class I molecules in the endoplasmic reticulum. Nature, 419, 480–483.
15) Saric, T., Chang, S.C., Hattori, A., York, I.A., Markant, S., Rock, K.L., Tsujimoto, M., & Goldberg, A.L. (2002) An IFN-γ-induced aminopeptidase in the ER, ERAP1, trims precursors to MHC class I-presented peptides. Nat. Immunol., 3, 1169–1176.
16) Beninga, J., Rock, K.L., & Goldberg, A.L. (1998) Interferon-γ can stimulate post-proteasomal trimming of the N terminus of an antigenic peptide by inducing leucine aminopeptidase. J. Biol. Chem., 273, 18734–18742.
17) Lorenzi, S., Forloni, M., Cifaldi, L., Antonucci, C., Citti, A., Boldrini, R., Pezzullo, M., Castellano, A., Russo, V., van der Bruggen, P., et al. (2012) IRF1 and NF-κB restore MHC class I-restricted tumor antigen processing and presentation to cytotoxic T cells in aggressive neuroblastoma. PLoS One, 7, e46928.
18) Forloni, M., Albini, S., Limongi, M.Z., Cifaldi, L., Boldrini, R., Nicotra, M.R., Giannini, G., Natali, P.G., Giacomini, P., & Fruci, D. (2010) NF-κB, and not MYCN, regulates MHC class I and endoplasmic reticulum aminopeptidases in human neuroblastoma cells. Cancer Res., 70, 916–924.
19) Goto, Y., Ogawa, K., Hattori, A., & Tsujimoto, M. (2011) Secretion of endoplasmic reticulum aminopeptidase 1 is involved in the activation of macrophages induced by lipopolysaccharide and interferon-γ. J. Biol. Chem., 286, 21906–21914.
20) Doody, G.M., Stephenson, S., McManamy, C., & Tooze, R.M. (2007) PRDM1/BLIMP-1 modulates IFN-γ-dependent control of the MHC class I antigen-processing and peptide-loading pathway. J. Immunol., 179, 7614–7623.
21) Paladini, F., Fiorillo, M.T., Vitulano, C., Tedeschi, V., Piga, M., Cauli, A., Mathieu, A., & Sorrentino, R. (2018) An allelic variant in the intergenic region between ERAP1 and ERAP2 correlates with an inverse expression of the two genes. Sci. Rep., 8, 10398.
22) Hanson, A.L., Cuddihy, T., Haynes, K., Loo, D., Morton, C.J., Oppermann, U., Leo, P., Thomas, G.P., Lê Cao, K.A., Kenna, T.J., et al. (2018) Genetic variants in ERAP1 and ERAP2 associated with immune-mediated diseases iInfluence protein expression and the isoform profile. Arthritis Rheumatol., 70, 255–265.
23) Andrés, A.M., Dennis, M.Y., Kretzschmar, W.W., Cannons, J.L., Lee-Lin, S.Q., Hurle, B., Schwartzberg, P.L., Williamson, S.H., Bustamante, C.D., Nielsen, R., et al.; NISC Comparative Sequencing Program. (2010) Balancing selection maintains a form of ERAP2 that undergoes nonsense-mediated decay and affects antigen presentation. PLoS Genet., 14, e1001157.
24) Ye, C.J., Chen, J., Villani, A.C., Gate, R.E., Subramaniam, M., Bhangale, T., Lee, M.N., Raj, T., Raychowdhury, R., Li, W., et al. (2018) Genetic analysis of isoform usage in the human anti-viral response reveals influenza-specific regulation of ERAP2 transcripts under balancing selection. Genome Res., 28, 1812–1825.
25) Kim, S., Lee, S., Shin, J., Kim, Y., Evnouchidou, I., Kim, D., Kim, Y.K., Kim, Y.E., Ahn, J.H., Riddell, S.R., et al. (2011) Human cytomegalovirus microRNA miR-US4-1 inhibits CD8+ T cell responses by targeting the aminopeptidase ERAP1. Nat. Immunol., 12, 984–991.
26) Romania, P., Cifaldi, L., Pignoloni, B., Starc, N., D’Alicandro, V., Melaiu, O., Li Pira, G., Giorda, E., Carrozzo, R., Bergvall, M., et al. (2017) Identification of a Genetic Variation in ERAP1 aminopeptidase that prevents human cytomegalovirus miR-UL112-5p-mediated immunoevasion. Cell Rep., 20, 846–853.
27) Nguyen, T.T., Chang, S.C., Evnouchidou, I., York, I.A., Zikos, C., Rock, K.L., Goldberg, A.L., Stratikos, E., & Stern, L. (2011) Structural basis for antigenic peptide precursor processing by the endoplasmic reticulum aminopeptidase ERAP1. Nat. Struct. Mol. Biol., 18, 604–613.
28) Kochan, G., Krojer, T., Harvey, D., Fischer, R., Chen, L., Vollmar, M., von Delft, F., Kavanagh, K.L., Brown, M.A., Bowness, P., et al. (2011) Crystal structures of the endoplasmic reticulum aminopeptidase-1 (ERAP1) reveal the molecular basis for N-terminal peptide trimming. Proc. Natl. Acad. Sci. USA, 108, 7745–7750.
29) Birtley, J.R., Saridakis, E., Stratikos, E., & Mavridis, I.M. (2012) The crystal structure of human endoplasmic reticulum aminopeptidase 2 reveals the atomic basis for distinct roles in antigen processing. Biochemistry, 51, 286–295.
30) Mpakali, A., Giastas, P., Mathioudakis, N., Mavridis, I.M., Saridakis, E., & Stratikos, E. (2015) Structural basis for antigenic peptide recognition and processing by Endoplasmic Reticulum (ER) Aminopeptidase 2 (2015). J. Biol. Chem., 290, 26021–26032.
31) Hermans, S.J., Ascher, D.B., Hancock, N.C., Holien, J.K., Michell, B.J., Chai, S.Y., Morton, C.J., & Parker, N.W. (2015) Crystal structure of human insulin-regulated aminopeptidase with specificity for cyclic peptides. Protein Sci., 24, 190–199.
32) Mpakali, A., Saridakis, E., Harlos, K., Zhao, Y., Papakyriakou, A., Kokkala, P., Georgiadis, D., & Stratikos, E. (2015) Crystal structure of insulin-regulated aminopeptidase with bound substrate analogue provides insight on antigenic epitope precursor recognition and processing. J. Immunol., 195, 2842–2851.
33) Mpakali, A., Saridakis, E., Harlos, K., Zhao, Y., Kokkala, P., Georgiadis, D., Giastas, P., Papakyriakou, A., & Stratikos, E. (2017) Ligand-induced conformational change of insulin-regulated aminopeptidase:insights on catalytic mechanism and active site plasticity. J. Med. Chem., 60, 2963–2972.
34) Goto, Y., Tanji, H., Hattori, A., & Tsujimoto, M. (2008) Glutamine-181 is crucial in the enzymatic activity and substrate specificity of human endoplasmic-reticulum aminopeptidase-1. Biochem. J., 416, 109–116.
35) Węglarz-Tomczak, E., Vassiliou, S., & Mucha, A. (2016) Identification of methionine aminopeptidase 2 as a molecular target of the organoselenium drug ebselen and its derivatives/analogues: Synthesis, inhibitory activity and molecular modeling study. Bioorg. Med. Chem. Lett., 26, 4122–4126.
36) Kokkala, P., Mpakali, A., Mauvais, F.X., Papakyriakou, A., Daskalaki, I., Petropoulou, I., Kavvalou, S., Papathanasopoulou, M., Agrotis, S., Fonsou, T.M., et al. (2016) Optimization and structure–activity relationships of phosphinic pseudotripeptide inhibitors of aminopeptidases that generate antigenic peptides. J. Med. Chem., 59, 9107–9123.
37) Goto, Y., Hattori, A., Ishii, Y., Mizutani, S., & Tsujimoto, M. (2006) Enzymatic properties of human aminopeptidase A. Regulation of its enzymatic activity by calcium and angiotensin IV. J. Biol. Chem., 281, 23503–23513.
38) Ogawa, Y., Ohnishi, A., Goto, Y., Sakuma, Y., Watanabe, J., Hattori, A., & Tsujimoto, M. (2014) Role of glutamine-169 in the substrate recognition of human aminopeptidase B. Biochim. Biophys. Acta, 1840, 1872–1881.
39) Zervoudi, E., Papakyriakou, A., Georgiadou, D., Evnouchidou, I., Gajda, A., Poreba, M., Salvesen, G.S., Drag, M., Hattori, A., Swevers, L., et al. (2011) Probing the S1 specificity pocket of the aminopeptidases that generate antigenic peptides. Biochem. J., 435, 411–420.
40) Goto, Y., Hattori, A., Mizutani, S., & Tsujimoto, M. (2007) Asparatic acid 221 is critical in the calcium-induced modulation of the enzymatic activity of human aminopeptidase A. J. Biol. Chem., 282, 37074–37081.
41) Hattori, A., Kitatani, K., Matsumoto, H., Miyazawa, S., Rogi, T., Tsuruoka, N., Mizutani, S., Natori, Y., & Tsujimoto, M. (2000) Characterization of recombinant human adipocyte-derived leucine aminopeptidase expressed in Chinese hamster ovary cells. J. Biochem., 128, 755–762.
42) Hattori, A. & Tsujimoto, M. (2004) Processing of antigenic peptides by aminopeptidases. Biol. Pharm. Bull., 27, 777–780.
43) Hearn, A., York, I.A., & Rock, K.L. (2009) The specificity of trimming of MHC class I-presented peptides in the endoplasmic reticulum. J. Immunol., 183, 5526–5536.
44) York, I.A., Chang, S.C., Saric, T., Keys, J.A., Favreau, J.M., Goldberg, A.L., & Rock, K.L. (2002) The ER aminopeptidase ERAP1 enhances or limits antigen presentation by trimming epitopes to 8–9 residues. Nat. Immunol., 3, 1177–1184.
45) Chang, S.C., Momburg, F., Bhutani, N., & Goldberg, A.L. (2005) The ER aminopeptidase, ERAP1, trims precursors to lengths of MHC class I peptides by a “molecular ruler” mechanism. Proc. Natl. Acad. Sci. USA, 102, 17107–17112.
46) Evnouchidou, I., Momburg, F., Papakyriakou, A., Chroni, A., Leondiadis, L., Chang, S.C., Goldberg, A.L., & Stratikos, E. (2008) The internal sequence of the peptide-substrate determines its N-terminus trimming by ERAP1. PLoS One, 3, e3658.
47) Blanchard, N., Kanaseki, T., Escobar, H., Delebecque, F., Nagarajan, N.A., Reyes-Vargas, E., Crockett, D.K., Raulet, D.H., Delgado, J.C., & Shastri, N. (2010) Endoplasmic reticulum aminopeptidase associated with antigen processing defines the composition and structure of MHC class I peptide repertoire in normal and virus-infected cells. J. Immunol., 184, 3033–3042.
48) Saveanu, L., Carroll, O., Lindo, V., Del Val, M., Lopez, D., Lepelletier, Y., Greer, F., Schomburg, L., Fruci, D., Niedermann, G., et al. (2005) Concerted peptide trimming by human ERAP1 and ERAP2 aminopeptidase complexes in the endoplasmic reticulum. Nat. Immunol., 6, 689–697.
49) Lorente, E., Barriga, A., Johnstone, C., Mir, C., Jiménez, M., & López, D. (2013) Concerted in vitro trimming of viral HLA-B27-restricted ligands by human ERAP1 and ERAP2 aminopeptidases. PLoS One, 8, e79596.
50) Evnouchidou, I., Weimershaus, M., Saveanu, L., & van Endert, P. (2014) ERAP1–ERAP2 dimerization increases peptide-trimming efficiency. J. Immunol., 193, 901–908.
51) Chen, H., Li, L., Weimershaus, M., Evnouchidou, I., van Endert, P., & Bouvier, M. (2016) ERAP1–ERAP2 dimers trim MHC I-bound precursor peptides; implications for understanding peptide editing. Sci. Rep., 6, 28902.
52) Yamamoto, N., Nakayama, J., Yamakawa-Kobayashi, K., Hamaguchi, H., Miyazaki, R., & Arinami, T. (2002) Identification of 33 polymorphisms in the adipocyte-derived leucine aminopeptidase (ALAP) gene and possible association with hypertension. Hum. Mutat., 19, 251–257.
53) Goto, Y., Hattori, A., Ishii, Y., & Tsujimoto, M. (2006) Reduced activity of the hypertension-associated Lys528Arg mutant of human adipocyte-derived leucine aminopeptidase (A-LAP)/ER-aminopeptidase-1. FEBS Lett., 580, 1833–1838.
54) Hisatsune, C., Ebisui, E., Usui, M., Ogawa, N., Suzuki, A., Mataga, N., Takahashi-Iwanaga, H., & Mikoshiba, K. (2015) ERp44 exerts redox-dependent control of blood pressure at the ER. Mol. Cell, 58, 1015–1027.
55) Zee, R.Y.L., Rivera, A., Inostroza, Y., Ridker, P.M., Chasman, D.I., & Romero, J.R. (2018) Gene variation of endoplasmic reticulum aminopeptidases 1 and 2, and risk of blood pressure progression and incident hypertension among 17,255 initially healthy women. Int. J. Genomics, 2018, 2308585.
56) Schomburg, L., Kollmus, H., Friedrichsen, S., & Bauer, K. (2000) Molecular characterization of a puromycin-insensitive leucyl-specific aminopeptidase, PILS-AP. Eur. J. Biochem., 267, 3198–3207.
57) Miyashita, H., Yamazaki, T., Akada, T., Niizeki, O., Ogawa, M., Nishikawa, S., & Sato, Y. (2002) A mouse orthologue of puromycin-insensitive leucyl-specific aminopeptidase is expressed in endothelial cells and plays an important role in angiogenesis. Blood, 99, 3241–3249.
58) Akada, T., Yamazaki, T., Miyashita, H., Niizeki, O., Abe, M., Sato, A., Satomi, S., & Sato, Y. (2002) Puromycin insensitive leucyl-specific aminopeptidase (PILSAP) is involved in the activation of endothelial integrins. J. Cell. Physiol., 193, 253–262.
59) Yamazaki, T., Akada, T., Niizeki, O., Suzuki, T., Miyashita, H., & Sato, Y. (2004) Puromycin-insensitive leucyl-specific aminopeptidase (PILSAP) binds and catalyzes PDK1, allowing VEGF-stimulated activation of S6K for endothelial cell proliferation and angiogenesis. Blood, 104, 2345–2352.
60) Sato, Y. (2004) Role of aminopeptidase in angiogenesis. Biol. Pharm. Bull., 27, 772–776.
61) Watanabe, Y., Shibata, K., Kikkawa, F., Kajiyama, H., Ino, K., Hattori, A., Tsujimoto, M., & Mizutani, S. (2003) Adipocyte-derived leucine aminopeptidase suppresses angiogenesis in human endometrial carcinoma via renin-angiotensin system. Clin. Cancer Res., 9, 6497–6503.
62) Cui, X., Hawari, F., Alsaaty, S., Lawrence, M., Combs, C.A., Geng, W., Rouhani, F.N., Miskinis, D., & Levine, S.J. (2002) Identification of ARTS-1 as a novel TNFR1-binding protein that promotes TNFR1 ectodomain shedding. J. Clin. Invest., 110, 515–526.
63) Cui, X., Rouhani, F.N., Hawari, F., & Levine, S.J. (2003) An aminopeptidase, ARTS-1, is required for interleukin-6 receptor shedding. J. Biol. Chem., 278, 28677–28685.
64) Cui, X., Rouhani, F.N., Hawari, F., & Levine, S.J. (2003) Shedding of the type II IL-1 decoy receptor requires a multifunctional aminopeptidase, aminopeptidase regulator of TNF receptor type 1 shedding. J. Immunol., 171, 6814–6819.
65) Islam, A., Adamik, B., Hawari, F.I., Ma, G., Rouhani, F.N., Zhang, J., & Levine, S.J. (2006) Extracellular TNFR1 release requires the calcium-dependent formation of a nucleobindin 2-ARTS-1 complex. J. Biol. Chem., 281, 6860–6873.
66) Haroon, N., Tsui, F.W., Chiu, B., Tsui, H.W., & Inman, R.D. (2010) Serum cytokine receptors in ankylosing spondylitis: relationship to inflammatory markers and endoplasmic reticulum aminopeptidase polymorphisms. J. Rheumatol., 37, 1907–1910.
67) Blum, J.S., Wearsch, P.A., & Cresswell, P. (2013) Pathways of antigen processing. Annu. Rev. Immunol., 31, 443–473.
68) Hammer, G.E., Gonzalez, F., Champsaur, M., Cado, D., & Shastri, N. (2006) The aminopeptidase ERAAP shapes the peptide repertoire displayed by major histocompatibility complex class I molecules. Nat. Immunol., 7, 103–112.
69) York, I.A., Brehm, M.A., Zendzian, S., Towne, C.F., & Rock, K.L. (2006) Endoplasmic reticulum aminopeptidase 1 (ERAP1) trims MHC class I-presented peptides in vivo and plays an important role in immunodominance. Proc. Natl. Acad. Sci. USA, 103, 9202–9207.
70) James, E., Bailey, I., Sugiyarto, G., & Elliott, T. (2013) Induction of protective antitumor immunity through attenuation of ERAAP function. J. Immunol., 190, 5839–5846.
71) Hammer, G.E., Gonzalez, F., James, E., Nolla, H., & Shastri, N. (2007) In the absence of aminopeptidase ERAAP, MHC class I molecules present many unstable and highly immunogenic peptides. Nat. Immunol., 8, 101–108.
72) Cifaldi, L., Lo Monaco, E., Forloni, M., Giorda, E., Lorenzi, S., Petrini, S., Tremante, E., Pende, D., Locatelli, F., Giacomini, P., et al. (2011) Natural killer cells efficiently reject lymphoma silenced for the endoplasmic reticulum aminopeptidase associated with antigen processing. Cancer Res., 71, 1597–1606.
73) Aldhamen, Y.A., Seregin, S.S., Rastall, D.P., Aylsworth, C.F., Pepelyayeva, Y., Busuito, C.J., Godbehere-Roosa, S., Kim, S., & Amalfitano, A. (2013) Endoplasmic reticulum aminopeptidase-1 functions regulate key aspects of the innate immune response. PLoS One, 8, e69539.
74) Warthan, M.D., Washington, S.L., Franzese, S.E., Ramus, R.M., Kim, K.R., York, T.P., Stratikos, E., Strauss, J.F. 3rd, & Lee, E.D. (2018) The role of endoplasmic reticulum aminopeptidase 2 in modulating immune detection of choriocarcinoma. Biol. Reprod., 98, 309–322.
75) Cagliani, R., Riva, S., Biasin, M., Fumagalli, M., Pozzoli, U., Lo Caputo, S., Mazzotta, F., Piacentini, L., Bresolin, N., Clerici, M., et al. (2010) Genetic diversity at endoplasmic reticulum aminopeptidases is maintained by balancing selection and is associated with natural resistance to HIV-1 infection. Hum. Mol. Genet., 19, 4705–47014.
76) Biasin, M., Sironi, M., Saulle, I., de Luca, M., la Rosa, F., Cagliani, R., Forni, D., Agliardi, C., lo Caputo, S., Mazzotta, F., et al. (2013) Endoplasmic reticulum aminopeptidase 2 haplotypes play a role in modulating susceptibility to HIV infection. AIDS, 27, 1697–1706.
77) Reeves, E. & James, E. (2018) The role of polymorphic ERAP1 in autoinflammatory disease. Biosci. Rep., 38, BSR20171503.
78) Guasp, P., Barnea, E., González-Escribano, M.F., Jiménez-Reinoso, A., Regueiro, J.R., Admon, A., & López de Castro, J.A. (2017) The Behçet’s disease-associated variant of the aminopeptidase ERAP1 shapes a low-affinity HLA-B*51 peptidome by differential subpeptidome processing. J. Biol. Chem., 292, 9680–9689.
79) Martín-Esteban, A., Gómez-Molina, P., Sanz-Bravo, A., & López de Castro, J.A. (2014) Combined effects of ankylosing spondylitis-associated ERAP1 polymorphisms outside the catalytic and peptide-binding sites on the processing of natural HLA-B27 ligands. J. Biol. Chem., 289, 3978–3990.
80) Reeves, E., Edwards, C.J., Elliott, T., & James, E. (2013) Naturally occurring ERAP1 haplotypes encode functionally distinct alleles with fine substrate specificity. J. Immunol., 191, 35–43.
81) Reeves, E., Colebatch-Bourn, A., Elliott, T., Edwards, C.J., & James, E. (2014) Functionally distinct ERAP1 allotype combinations distinguish individuals with Ankylosing Spondylitis. Proc. Natl. Acad. Sci. USA, 111, 17594–17599.
82) Goto, Y., Ogawa, K., Nakamura, T.J., Hattori, A., & Tsujimoto, M. (2014) TLR-mediated secretion of endoplasmic reticulum aminopeptidase 1 from macrophages. J. Immunol., 192, 4443–4452.
83) Goto, Y., Ogawa, K., Nakamura, T.J., Hattori, A., & Tsujimoto, M. (2015) Substrate-dependent nitric oxide synthesis by secreted endoplasmic reticulum aminopeptidase 1 in macrophages. J. Biochem., 157, 439–449.
84) Goto, Y., Nakamura, T.J., Ogawa, K., Hattori, A., & Tsujimoto, M. (2019) Acute-phase protein-like properties of endoplasmic reticulum aminopeptidase 1. J. Biochem., 165, 159–165.
85) Bogdan, C. (2015) Nitric oxide synthase in innate and adaptive immunity: an update. Trends Immunol., 36, 161–178.
86) Goto, Y., Ogawa, Y., Tsumoto, H., Miura, Y., Nakamura, T.J., Ogawa, K., Akimoto, Y., Kawakami, H., Endo, T., Yanoshita, R., et al. (2018) Contribution of the exosome-associated form of secreted endoplasmic reticulum aminopeptidase 1 to exosome-mediated macrophage activation. Biochim. Biophys. Acta, 1865, 874–888.
87) Aldhamen, Y.A., Pepelyayeva, Y., Rastall, D.P., Seregin, S.S., Zervoudi, E., Koumantou, D., Aylsworth, C.F., Quiroga, D., Godbehere, S., Georgiadis, D., et al. (2015) Autoimmune disease-associated variants of extracellular endoplasmic reticulum aminopeptidase 1 induce altered innate immune responses by human immune cells. J. Innate Immun., 7, 275–289.
88) Watanabe, S., Amagai, Y., Sannino, S., Tempio, T., Anelli, T., Harayama, M., Masui, S., Sorrentino, I., Yamada, M., Sitia, R., et al. (2019) Zinc regulates ERp44-dependent protein quality control in the early secretory pathway. Nat. Commun., 10, 603.
89) Hattori, A., Goto, Y., & Tsujimoto, M. (2012) Exon 10 coding sequence is important for endoplasmic reticulum retention of endoplasmic reticulum aminopeptidase 1. Biol. Pharm. Bull., 35, 601–605.
90) Yamazaki, T., Akada, T., Niizeki, O., Suzuki, T., Miyashita, H., & Sato, Y. (2004) Puromycin-insensitive leucyl-specific aminopeptidase (PILSAP) binds and catalyzes PDK1, allowing VEGF-stimulated activation of S6K for endothelial cell proliferation and angiogenesis. Blood, 104, 2345–2352.
91) Islam, A., Adamik, B., Hawari, F.I., Ma, G., Rouhani, F.N., Zhang, J., & Levine, S.J. (2006) Extracellular TNFR1 release requires the calcium-dependent formation of a nucleobindin 2-ARTS-1 complex. J. Biol. Chem., 281, 6860–6873.
92) Adamik, B., Islam, A., Rouhani, F.N., Hawari, F.I., Zhang, J., & Levine, S.J. (2008) An association between RBMX, a heterogeneous nuclear ribonucleoprotein, and ARTS-1 regulates extracellular TNFR1 release. Biochem. Biophys. Res. Commun., 371, 505–509.
93) Stratikos, E., Stamogiannos, A., Zervoudi, E., & Fruci, D. (2014) A role for naturally occurring alleles of endoplasmic reticulum aminopeptidases in tumor immunity and cancer pre-disposition. Front. Oncol., 4, 363.
94) Rastall, D.P.W., Alyaquob, F.S., O’Connell, P., Pepelyayeva, Y., Peters, D., Godbehere-Roosa, S., Pereira-Hicks, C., Aldhamen, Y.A., & Amalfitano, A. (2017) Mice expressing human ERAP1 variants associated with ankylosing spondylitis have altered T-cell repertoires and NK cell functions, as well as increased in utero and perinatal mortality. Int. Immunol., 29, 277–289.
95) López de Castro, J.A. (2018) How ERAP1 and ERAP2 shape the peptidomes of disease-associated MHC-I proteins. Front. Immunol., 9, 2463.
96) Burton, P.R., Clayton, D.G., Cardon, L.R., Craddock, N., Deloukas, P., Duncanson, A., Kwiatkowski, D.P., McCarthy, M.I., Ouwehand, W.H., Samani, N.J., et al.Breast Cancer Susceptibility Collaboration (UK). (2007) Association scan of 14,500 nonsynonymous SNPs in four diseases identifies autoimmunity variants. Nat. Genet., 39, 1329–13237.
97) Hanson, A.L., Morton, C.J., Parker, M.W., Bessette, D., & Kenna, T.J. (2019) The genetics, structure and function of the M1 aminopeptidase oxytocinase subfamily and their therapeutic potential in immune-mediated disease. Hum. Immunol., 80, 281–289.
98) Kirino, Y., Bertsias, G., Ishigatsubo, Y., Mizuki, N., Tugal-Tutkun, I., Seyahi, E., Ozyazgan, Y., Sacli, F.S., Erer, B., Inoko, H., et al. (2013) Genome-wide association analysis identifies new susceptibility loci for Behçet’s disease and epistasis between HLA-B*51 and ERAP1. Nat. Genet., 45, 202–207.
99) Genetic Analysis of Psoriasis Consortium & the Wellcome Trust Case Control Consortium 2 (2010) A genome-wide association study identifies new psoriasis susceptibility loci and an interaction between HLA-C and ERAP1. Nat. Genet., 42, 985–990.
100) Hill, L.D., Hilliard, D.D., York, T.P., Srinivas, S., Kusanovic, J.P., Gomez, R., Elovitz, M.A., Romero, R., & Strauss, J.F. 3rd. (2011) Fetal ERAP2 variation is associated with preeclampsia in African Americans in a case-control study. BMC Med. Genet., 12, 64.
101) Vanhille, D.L., Hill, L.D., Hilliard, D.D., Lee, E.D., Teves, M.E., Srinivas, S., Kusanovic, J.P., Gomez, R., Stratikos, E., Elovitz, M.A., et al. (2013) A novel ERAP2 haplotype structure in a chilean population: implications for ERAP2 protein expression and preeclampsia risk. Mol. Genet. Genomic Med., 1, 98–107.
102) Evnouchidou, I., Birtley, J., Seregin, S., Papakyriakou, A., Zervoudi, E., Samiotaki, M., Panayotou, G., Giastas, P., Petrakis, O., Georgiadis, D., et al. (2012) A common single nucleotide polymorphism in endoplasmic reticulum aminopeptidase 2 induces a specificity switch that leads to altered antigen processing. J. Immunol., 189, 2383–2392.
103) Yao, Y., Liu, N., Zhou, Z., & Shi, L. (2018) Influence of ERAP1 and ERAP2 gene polymorphisms on disease susceptibility in different populations. Hum. Immunol., 80, 325–334.
104) Manguso, R.T., Pope, H.W., Zimmer, M.D., Brown, F.D., Yates, K.B., Miller, B.C., Collins, N.B., Bi, K., LaFleur, M.W., Juneja, V.R., et al. (2017) In vivo CRISPR screening identifies Ptpn2 as a cancer immunotherapy target. Nature, 27, 413–418.
105) Lim, Y.W., Chen-Harris, H., Mayba, O., Lianoglou, S., Wuster, A., Bhangale, T., Khan, Z., Mariathasan, S., Daemen, A., Reeder, J., et al. (2018) Germline genetic polymorphisms influence tumor gene expression and immune cell infiltration. Proc. Natl. Acad. Sci. USA, 115, E11701–E11710.
106) Compagnone, M., Cifaldi, L., & Fruci, D. (2019) Regulation of ERAP1 and ERAP2 genes and their disfunction in human cancer. Hum. Immunol., 80, 318–324.
107) Albiston, A.L., McDowall, S.G., Matsacos, D., Sim, P., Clune, E., Mustafa, T., Lee, J., Mendelsohn, F.A., Simpson, R.J., Connolly, L.M., et al. (2001) Evidence that the angiotensin IV (AT(4)) receptor is the enzyme insulin-regulated aminopeptidase. J. Biol. Chem., 276, 48623–48626.
108) Wright, J.W. & Harding, J.W. (2019) Contributions by the brain renin-angiotensin system to memory, cognition, and Alzheimer’s disease. J. Alzheimers Dis., 67, 469–480.
著者紹介Author Profile
 辻本 雅文(つじもと まさふみ)
辻本 雅文(つじもと まさふみ)帝京平成大学薬学部教授.薬学博士.
略歴1952年大阪府に生る.81年東京大学大学院薬学研究科博士課程修了,同年サントリー(株)生物医学研究所研究員,96年理化学研究所主任研究員,2011年より現職.
研究テーマと抱負疾患に関与する可能性があり,興味深い活性を示すタンパク質としてM1アミノペプチダーゼを取り上げている.その構造と機能を解析することで,病態への関与やクスリの開発に結びつくことを期待している.
趣味乗り鉄,読書(時代小説),ジョギング.