もやもや病遺伝子の代謝制御機能Metabolic function of the moyamoya disease gene mysterin/RNF213
昭和大学医学部生化学講座Showa University School of Medicine ◇ 〒142–8555 東京都品川区旗の台1–5–8 ◇ Hatanodai 1–5–8, Shinagawa, Tokyo 142–8555, Japan
発行日:2019年12月25日Published: December 25, 2019
© 2019 公益社団法人日本生化学会© 2019 The Japanese Biochemical Society
もやもや病は東アジアに多い原因不明の脳血管疾患である1).頭蓋内の限局された領域(内頚動脈終末部周辺)において進行性の動脈狭窄・閉塞を左右両側性に生じ,脳虚血・脳梗塞が引き起こされる.発病は10歳以下の子どもに多く,また女性での発病が男性の約2倍多い.病変部においては血管平滑筋細胞の異常な増殖と血管内膜への浸潤(それによる内膜の肥厚と血管内腔の狭小化)などを認めるが,病因・病態の進行プロセスとも謎のままである.また病変が内頚動脈終末部に限られる理由についても解明されていない.発病後期には煙状の特徴的な側副血行路(もやもや血管)が病変部周囲に形成され脳虚血の解消に働くと考えられるが,成人後にここから出血を生じることがありかえって症状を増悪させる.
もやもや病の有病率は民族間で約10倍異なっており(東アジア人種1/10,000 vs白人1/100,000),また東西ともに家族性発症を認めることから民族性および家族性の遺伝要因の寄与が推定されてきた.東北大学医学部・呉繁夫教授を中心とする研究グループにより東アジア孤発例の,また京都大学医学部・小泉昭夫教授を中心とする研究グループにより東アジア家族例の遺伝解析が行われ,興味深いことに17番染色体長腕末端領域に同一の変異が見いだされた2, 3).本変異は新規遺伝子の読み枠内にあり,同遺伝子産物のアルギニンをリシンに置き換えるミスセンス変異(p.R4810K)であった.小泉教授らとの共同研究として筆者の手により本遺伝子の分子クローニングを行い,ミステリン(mysterin:moyamoya steno-occlusive disease-associated AAA+ and RING finger protein(別名RNF213, ALO17)と名づけ,機能解析を開始した3).
ミステリンの生理・病態機能について議論する前に遺伝学の成果についてもう少しふれておきたい.ミステリン遺伝子はホヤ,脊椎動物を含む脊索動物に保存されており,ヒト,マウスでは約600 kDaの巨大な細胞内タンパク質をコードする.ミステリンタンパク質は,二つの隣接するAAA+型ATPアーゼドメインおよび一つのRING finger型ユビキチンリガーゼドメインを含んでいる4)(図1).これまでのところ他に信頼できるドメイン構造は見いだされていない.ヒト,マウスでは血管平滑筋,血管内皮を含むユビキタスな発現パターンを示すが,特に脾臓,白血球など免疫関連組織に発現が高い.加えてインターフェロンなどによる強い誘導を受けることがわかっており,炎症との関連が示唆されている5, 6).
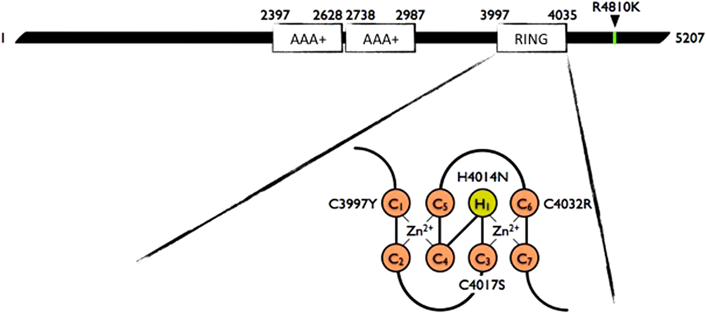
5207アミノ酸長の分子内に二つのAAA+ドメインと一つのRING fingerドメインを持つ.東アジア人患者の多くに認められるR4810K変異はC末端機能未知領域内に位置する.白人患者からRING fingerドメインにおける亜鉛イオンのキレートに必須のシステイン/ヒスチジン変異(C3997Y, C4017S, H4014N, C4032R変異)が同定されている.
呉,小泉らの遺伝解析により見いだされたp.R4810KはミステリンのC末端機能未知領域内に位置する(図1).日本人全体の1~2%程度がp.R4810K保因者であると見積もられており,したがって遺伝学的には変異(集団内の1%以下が持つ多型)ではなく,まれな一塩基多型と呼ぶ方が正確に近い.興味深いことに孤発例,家族例を問わず,日本人もやもや病患者の多くがp.R4810K保因者によって占められていた(孤発例の約70%および家族例の100%)2, 3).すなわち従来孤発例と考えられてきた患者の多くも,特定の遺伝的バックグラウンド(p.R4810K)を元に発病していたものと考えられる.またp.R4810Kは,孤発例と考えられてきた患者と家族例と考えられてきた患者を区別する遺伝要因ではなかったといえる(家族例を特徴づける因子はいまだ同定されていない).前述のとおり日本人もやもや病有病率は約1/10,000(0.01%)であり,総人口の1~2%を占めるp.R4810K保因者のごく一部しか発病していない.すなわち,p.R4810Kは発病の感受性を著明に高める感受性変異ではあるが,発病に至るには追加の遺伝もしくは環境要因が必要と考えられる(たとえば別の遺伝的変異,炎症などが考えられている).東アジアもやもや病家系において一卵性双生児の片方のみが発病する例があることなどから,もやもや病は単一の遺伝要因では説明できない多因子疾患であると推定されてきた.遺伝解析により判明した東アジアp.R4810Kの不完全な遺伝的効果(浸透率)はこの推定を裏づけている.
2014年の年末から2017年の夏ごろにかけて,欧米の三つのグループから白人もやもや病におけるミステリン遺伝子のシークエンス解析が報告された7–9).ミステリンのRING finger型ユビキチンリガーゼドメインは二つの亜鉛イオンを七つのシステインと一つのヒスチジンでキレートする構造をとると考えられるが,4人の白人もやもや病患者からこれら保存されたシステインもしくはヒスチジンの変異が四つ単離された(図1).うち二つは両親いずれにも変異を認めず,患者にのみ認めるde novo変異(突然変異)であった.これら変異の保因者が4人しか見つかっていないことから統計解析にはなじまないが,4人全員がもやもや病を発症していることから,これらの変異は「感受性変異」ではなく単一で発病を引き起こしうる「原因変異」である可能性が高い.従来ミステリンをもやもや病の感受性遺伝子と呼ぶことが多かったが,原因遺伝子もしくは責任遺伝子と呼んで差支えないようにも思われる.
またここでは詳細にふれないが,p.R4810Kがもやもや病以外に動脈硬化性の脳梗塞とも相関することが報告されている10, 11).もやもや病の病因・病態を理解する上で重要なヒントになるかもしれない.
前述のとおり,ミステリンは二つのAAA+ ATPアーゼドメインと一つのRING finger型ユビキチンリガーゼドメインを持つ(図1).AAA+ ATPアーゼはドーナツ状複合体(多くは六量体)の形成,サブユニット間での協調したATP加水分解に伴うダイナミックな全体構造変化,それによる種々の細胞内物理プロセス(カーゴ輸送,膜動態制御,タンパク質複合体解離など)への寄与などを特徴とするユニークなATPアーゼ群である12).ダイニン,p97/VCP,プロテアソームATPアーゼサブユニットなどがこのクラスのATPアーゼに含まれる.培養細胞より精製した全長ミステリンが電子顕微鏡下で直径20 nm程度の大きなドーナツ状粒子として観察されたことから,ミステリンを正統的なAAA+ ATPアーゼに含めてよいように思われた.一方で,蛍光分子相関法(FCS)によるミステリンの分子サイズ推定や野生型/ATPアーゼ変異型ミステリン分子どうしの結合効率測定などから,細胞内ではミステリンの複合体形成は一過的でありATP結合を契機とした集合とATP加水分解を契機とした解離を繰り返していることが示唆された(図2).大部分のAAA+ ATPアーゼは生理的濃度のATP/マグネシウムイオン存在下で安定な複合体を形成するが,一過的に複合体を形成する例外的なメンバーも存在し,ミステリン以外にカタニン,Vps4などが知られている.後述するとおり,筆者らはミステリンはAAA+ ATPアーゼ活性により脂肪滴表面のプロテオーム再編成に働く可能性を考えているが,その実態・メカニズムについてはさらに詳しい解析を要する.
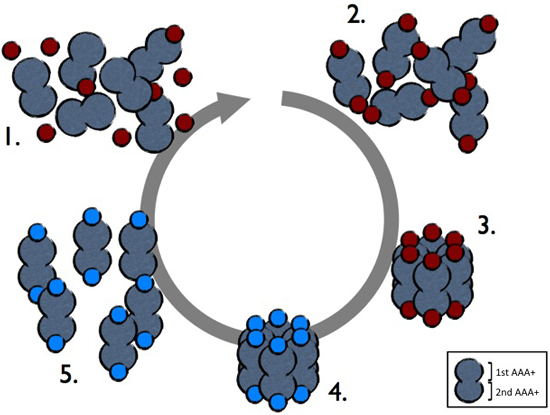
ミステリンは通常単量体として細胞質ゾルに存在し(1),一つ目のAAA+ドメインにATPが結合すると(2),集合して複合体を形成する(3).二つ目のAAA+ドメインで別のATPを加水分解することで(4),解離して単量体にもどる(5).赤丸と青丸がそれぞれATPとADPを表す.
また,ミステリンは知られている中では唯一のATPアーゼ/ユビキチンリガーゼハイブリッド酵素である.ミステリンをHEK293細胞などに過剰発現した場合,自己ユビキチン化活性が検出できる3).このような活性は過剰発現に伴う非生理的なものにすぎないようにも思われたが,次節に述べるとおり,ミステリンの正常機能には非分解性の自己ユビキチン化(による自己制御)が重要である可能性が示唆されている.この他,血管リモデリングに関わるいくつかのWnt経路関連因子などが基質候補として見いだされているが,直接のユビキチン化は確認されておらず検証を要する13).
ミステリンの基質や機能を同定する目的でミステリン発現抑制ゼブラフィッシュの表現型解析や細胞内結合タンパク質探索などを行い,広範な初期発生異常の検出やミステリン安定化因子の捕捉に成功した14, 15).しかし,基質・細胞内機能をつきとめるには至らなかった.ミステリンをHeLa細胞などに過剰発現すると,細胞播種後,培地が貧栄養化する前の比較的早い時間帯に直径1 µm程度の粒子状構造を無数に形成するのが観察される.デコンボリューション処理や高倍率対物レンズを用いた観察により,これらの構造は中空であることがわかった.すなわちミステリンは何らかの小胞/粒子状構造を取り囲むように局在することが示唆された.そこでリソソーム,エンドソーム,ペルオキシソームあるいはストレス顆粒などの小胞/粒子状構造との共局在を検討したところ,思いがけないことにミステリンは細胞内の脂肪貯蔵サイトである脂肪滴に表在していた16).
脂肪滴はほとんどすべての組織・細胞において形成される細胞内構造で,中性脂肪,コレステロールエステル,脂溶性ビタミンなどの貯蔵に働く.小胞体膜上で合成された中性脂肪やコレステロールエステルは高度に水不溶性であり,小胞体脂質二重膜の疎水面に蓄積する.蓄積が少量である間はレンズ状の形態をとり,蓄積が増大するに従って安定な球状構造に移行し,やがて小胞体膜から切り離されて細胞質ゾル中に放出される(図3).したがって脂肪滴の表面は小胞体膜由来の脂質一重膜に覆われているが,その表面に脂肪合成酵素,分解酵素,制御因子などが結合してインスリン,カテコールアミンなどの刺激や細胞の貧/富栄養状態に応じて,活発に脂質の出し入れ(分解と合成)を行う.蓄積された脂肪は分解・酸化されてミトコンドリアでの好気呼吸に用いられる他,リン脂質を含む各種脂質関連分子の合成ソースとしても重要だと考えられている.
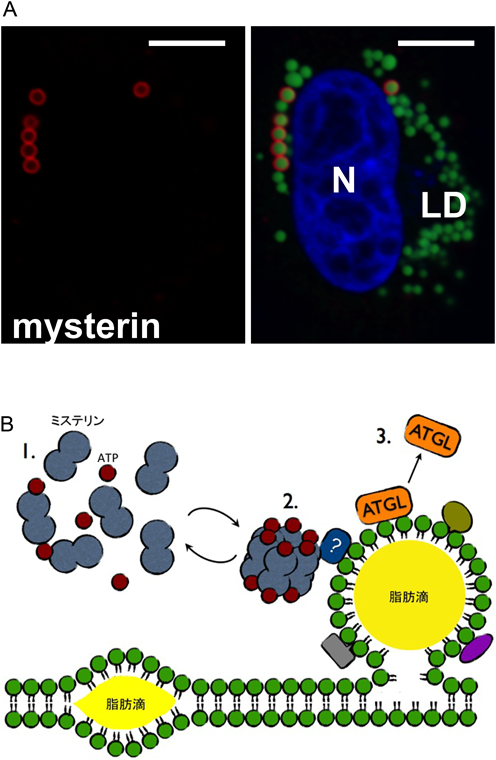
(A)ミステリンは直径1 µm程度のリング状構造をとる(左図.共焦点顕微鏡による焦点面観察であり実際には球状構造と思われる).リングの内部には中性脂肪が充満していたことから,この構造が脂肪滴であることが分かる(右図).N, LDはそれぞれ核,脂肪滴を表す.スケールバーは10 µmを示す.(B)小胞体膜上で合成された中性脂肪やコレステロールエステルは小胞体脂質二重膜の疎水面に蓄積して,初期にはレンズ状の構造,蓄積に従って球状の構造をとるようになる.脂肪滴の表面にはATGLはじめ複数のタンパク質が特異的に結合し,脂肪滴の挙動を制御する.ミステリンは通常単量体として細胞質ゾルに局在し(1),ATP結合に従って脂肪滴に移行,複合体化し(2),脂肪滴からATGLを解離させる(3).ATGL解離のメカニズムは不明であるが,未同定のタンパク質アンカーを介して間接的にATGLに働きかける可能性などが考えられる.
HeLa細胞などにおいてミステリンを過剰発現したとき,脂肪滴の数・量は著明に増大し,逆にミステリンの発現抑制/ノックアウトによって減少する.このことからミステリンは脂肪合成を促進するか,あるいは脂肪分解を抑制することが考えられた.脂肪合成の阻害剤(トリアクシンC)および脂肪分解の阻害剤(オーリスタット)を用いた実験から,ミステリンは脂肪合成経路ではなく脂肪分解経路を調節することで脂肪滴量の増大に働くことが示唆された.脂肪滴の主成分である中性脂肪(トリグリセリド)は3種のリパーゼATGL(adipose triglyceride lipase),HSL(hormone-sensitive lipase),MGL(monoglyceride lipase)により段階的に分解されるが,このうちトリグリセリドから最初の脂肪酸鎖を切り離すATGLが律速酵素であり,最も影響が大きい.興味深いことに,ミステリンの過剰発現により脂肪滴上からATGLが消失し,発現抑制/ノックアウトによりATGLの脂肪滴局在が著明に増大した.すなわち,ミステリンはATGLを脂肪滴から排除することで脂肪分解を止め,脂肪滴安定化に働く因子であった16).
ミステリンの脂肪滴局在にはATPアーゼサイクルが重要な役割を果たしていた.ミステリンATPアーゼドメイン内のATP結合モチーフおよびATP加水分解に必須のMg2+結合モチーフに変異を導入することで,それぞれミステリンを単量体優位および複合体優位の状態に固定することができる.いずれの変異導入によってもミステリンは脂肪滴に局在しなくなり,細胞質ゾルにおいて拡散性の局在を示すようになった.すなわちミステリンの局在化にはATPアーゼサイクル(と同調した複合体サイクル)の進行が重要であると考えられた(図3).ミステリンと類似の複合体形成サイクルを示すカタニン,Vps4は,それぞれ基質である微小管,ESCRT-III複合体上に集合してそれら高次構造の解離に働く[セグリガーゼ(segregase)活性].ミステリンの直接の標的は同定されていないが(ATGLとの結合は検出されていない),脂肪滴タンパク質複合体の解離に働くことで間接的にATGLを排除する可能性が考えられた16).
また興味深いことにユビキチンリガーゼ活性も重要であった.ユビキチンリガーゼドメインの欠損および前述の白人患者原因変異(図1)によってミステリンの脂肪滴局在は著明に障害され,凝集様の構造体を形成するようになった.多くの場合,ユビキチンリガーゼは基質タンパク質をユビキチン化し,活性・安定性などの制御に関わるが,ミステリンユビキチンリガーゼドメインの変異によりミステリン自身の挙動が変化したことから,自己ユビキチン化によって自己の挙動を制御している可能性が示唆された16).今後,ミステリン自己ユビキチン化の様式(鎖型,位置,機能など)の検証が必要である.
筆者らの検討した範囲において,野生型ミステリンと東アジアp.R4810K変異体の間に機能的な違いを見いだすことはできていない3, 4, 14–16).遺伝学的にp.R4810Kがもやもや病と相関することは間違いないと考えられるが,その病因効果は弱いもしくは潜在的(他の要因を必要とする)なものにとどまっており,分子レベルの挙動においても同様であるように思える.一方で白人変異体は凝集様の構造を形成し脂肪滴への局在をほとんど失う16).凝集の持つであろう細胞毒性と代謝機能の喪失のどちらかあるいは両方がもやもや病の病因ではないかと筆者らは考えている.また未発表であるが,ミステリンはATGL以外にもいくつかの因子を選択的に脂肪滴から排除する.脂肪滴の挙動は表在性プロテオームの組成・活性により決定されると考えられるが,その編成は組織間で,あるいは同一細胞内であっても各脂肪滴ごとに大きく異なる.脂肪滴表在性プロテオームがどのように特異的に編成されるかは関連領域の中心的な課題の一つであるが,ミステリンが特異的プロテオーム編成システムの一翼を担っていることは十分に考えられる.今後は,変異ミステリンがもやもや病を引き起こす仕組み(病態機能)の解明に加えて,ミステリンの生理的な働き(生理機能)の解明に取り組んでいきたい.
ミステリンの分子クローニング,活性・機能同定の大部分は京都大学・京都産業大学(永田和宏教授)において行われた.多数の共同研究者の貢献に加えて,最近では特に會退詩央莉氏(京都産業大学)の貢献が大きかった.本稿の図はすべて折井みなみ博士(昭和大学)によって作成された.これらの先生方および現所属の宮崎章教授にあらためて謝意を表したい.
1) 冨永悌二,鈴木則宏,宮本亨,小泉昭夫,黒田敏,高橋淳,藤村幹,寶金清博(2018)もやもや病(ウイリス動脈輪閉塞症)診断・治療ガイドライン(改訂版).脳卒中の外科,46, 1–24.
2) Kamada, F., Aoki, Y., Narisawa, A., Abe, Y., Komatsuzaki, S., Kikuchi, A., Kanno, J., Niihori, T., Ono, M., Ishii, N., et al. (2011) A genome-wide association study identifies RNF213 as the first Moyamoya disease gene. J. Hum. Genet., 56, 34–40.
3) Liu, W., Morito, D., Takashima, S., Mineharu, Y., Kobayashi, H., Hitomi, T., Hashikata, H., Matsuura, N., Yamazaki, S., Toyoda, A., et al. (2011) Identification of RNF213 as a susceptibility gene for moyamoya disease and its possible role in vascular development. PLoS One, 6, e22542.
4) Morito, D., Nishikawa, K., Hoseki, J., Kitamura, A., Kotani, Y., Kiso, K., Kinjo, M., Fujiyoshi, Y., & Nagata, K. (2014) Moyamoya disease-associated protein mysterin/RNF213 is a novel AAA+ ATPase, which dynamically changes its oligomeric state. Sci. Rep., 4, 4442.
5) Kobayashi, H., Matsuda, Y., Hitomi, T., Okuda, H., Shioi, H., Matsuda, T., Imai, H., Sone, M., Taura, D., Harada, K.H., et al. (2015) Biochemical and Functional Characterization of RNF213 (Mysterin) R4810K, a Susceptibility Mutation of Moyamoya Disease, in Angiogenesis In Vitro and In Vivo. J. Am. Heart Assoc., 4, e002146.
6) Ohkubo, K., Sakai, Y., Inoue, H., Akamine, S., Ishizaki, Y., Matsushita, Y., Sanefuji, M., Torisu, H., Ihara, K., Sardiello, M., et al. (2015) Moyamoya disease susceptibility gene RNF213 links inflammatory and angiogenic signals in endothelial cells. Sci. Rep., 5, 13191.
7) Cecchi, A.C., Guo, D., Ren, Z., Flynn, K., Santos-Cortez, R.L., Leal, S.M., Wang, G.T., Regalado, E.S., Steinberg, G.K., Shendure, J., et al.; University of Washington Center for Mendelian Genomics. (2014) RNF213 rare variants in an ethnically diverse population with Moyamoya disease. Stroke, 45, 3200–3207.
8) Raso, A., Biassoni, R., Mascelli, S., Nozza, P., Ugolotti, E.D.I., Marco, E.D.E., Marco, P., Merello, E., Cama, A., Pavanello, M., et al. (2016) Moyamoya vasculopathy shows a genetic mutational gradient decreasing from East to West. J. Neurosurg. Sci., 27.
9) Guey, S., Kraemer, M., Hervé, D., Ludwig, T., Kossorotoff, M., Bergametti, F., Schwitalla, J.C., Choi, S., Broseus, L., Callebaut, I., et al.; FREX consortium. (2017) Rare RNF213 variants in the C-terminal region encompassing the RING-finger domain are associated with moyamoya angiopathy in Caucasians. Eur. J. Hum. Genet., 25, 995–1003.
10) Miyawaki, S., Imai, H., Shimizu, M., Yagi, S., Ono, H., Mukasa, A., Nakatomi, H., Shimizu, T., & Saito, N. (2013) Genetic variant RNF213 c.14576G>A in various phenotypes of intracranial major artery stenosis/occlusion. Stroke, 44, 2894–2897.
11) Okazaki, S., Morimoto, T., Kamatani, Y., Kamimura, T., Kobayashi, H., Harada, K., Tomita, T., Higashiyama, A., Takahashi, J.C., Nakagawara, J., et al. (2019) Moyamoya Disease Susceptibility Variant RNF213 p.R4810K Increases the Risk of Ischemic Stroke Attributable to Large-Artery Atherosclerosis. Circulation, 139, 295–298.
12) Ogura, T. & Wilkinson, A.J. (2001) AAA+ superfamily ATPases: common structure–diverse function. Genes Cells, 6, 575–597.
13) Scholz, B., Korn, C., Wojtarowicz, J., Mogler, C., Augustin, I., Boutros, M., Niehrs, C., & Augustin, H.G. (2016) Endothelial RSPO3 Controls Vascular Stability and Pruning through Non-canonical WNT/Ca2+/NFAT Signaling. Dev. Cell, 36, 79–93.
14) Kotani, Y., Morito, D., Yamazaki, S., Ogino, K., Kawakami, K., Takashima, S., Hirata, H., & Nagata, K. (2015) Neuromuscular regulation in zebrafish by a large AAA+ ATPase/ubiquitin ligase, mysterin/RNF213. Sci. Rep., 5, 16161.
15) Kotani, Y., Morito, D., Sakata, K., Ainuki, S., Sugihara, M., Hatta, T., Iemura, S.I., Takashima, S., Natsume, T., & Nagata, K. (2017) Alternative exon skipping biases substrate preference of the deubiquitylase USP15 for mysterin/RNF213, the moyamoya disease susceptibility factor. Sci. Rep., 7, 44293.
16) Sugihara, M., Morito, D., Ainuki, S., Hirano, Y., Ogino, K., Kitamura, A., Hirata, H., & Nagata, K. (2019) The AAA+ ATPase/ubiquitin ligase mysterin stabilizes cytoplasmic lipid droplets. J. Cell Biol., 218, 949–960.

昭和大学医学部講師.博士(理学).
1975年京都市生.99年京都大学理学部卒業.2008年同大学院理学研究科修了.京都大学再生医科学研究所(研究員),京都産業大学総合生命科学部/タンパク質動態研究所(研究員/主任研究員)を経て,18年より現職.
研究テーマと抱負ミステリンの病態機能解明をまず進めてから,ゆっくり生理機能探索を楽みたい.
ウェブサイトhttp:// researchmap.jp/7000011224/
趣味研究(いい意味でも悪い意味でも趣味です).

This page was created on 2019-10-28T14:20:54.778+09:00
This page was last modified on 2019-12-02T16:23:20.000+09:00
このサイトは(株)国際文献社によって運用されています。