ユビキチン修飾は生体に必須の翻訳後修飾であり,タンパク質の品質管理,シグナル伝達,DNA修復,細胞内輸送,転写とエピゲノム制御など,きわめて広範な細胞機能を制御する.ユビキチン修飾系の機能破綻はがんや神経性変性疾患,炎症疾患などさまざまな疾病に関与することも明らかになっている.ユビキチンは76アミノ酸からなる小型のタンパク質であり,C末端が基質タンパク質のリシン残基に付加されることで,翻訳後修飾として機能する.ユビキチン修飾は可逆的であり,ユビキチン活性化酵素(E1),ユビキチン連結酵素(E2),ユビキチンリガーゼ(E3)のカスケードによって基質に連結され,脱ユビキチン化酵素(deubiquitinase:DUB)によって除去される.主にE3の基質特異性により,基質のユビキチン化は時空間的にダイナミックに調節されている.
ユビキチン修飾の機能的な多様性の分子基盤として,ユビキチン鎖の構造多様性があげられる1).ユビキチン自身が7か所のリシン残基を有しているため,リシン残基あるいはN末端アミノ基を介して連結し,8種類の連結タイプ(K6, K11, K27, K29, K33, K48, K63, M1)のポリユビキチン鎖を形成する.これら構造の異なる8種類のユビキチン鎖およびモノユビキチンは,各々特異的なシグナルとして機能すると考えられている.たとえば,リシン48(K48)を介して連結したK48ユビキチン鎖(K48鎖)はプロテアソーム依存性タンパク質分解の目印として機能し,リシン63(K63)を介して連結したK63ユビキチン鎖(K63鎖)はシグナル伝達やエンドサイトーシスなど,プロテアソーム非依存性の経路を制御する.このようなユビキチン鎖高次構造に埋め込まれた多様な機能情報はユビキチンコードと称されるに至っている1).
ユビキチンコードの制御因子は,コードを形成するWriter(E2, E3),コードを消去するEraser(DUB),コードを解読するデコーダーまたはReader(ユビキチン結合タンパク質)に分類される.まず,WriterであるE2, E3は特定の連結型のユビキチン鎖を形成する.ただし,連結型の特異性を有さないE2, E3も多く存在する.EraserであるDUBも同様に,その一部は特定の連結型のユビキチン鎖を特異的に切断する.またReaderはさまざまな種類のユビキチン結合ドメイン(ubiquitin-binding domain:UBD)を有しており,これらドメインが特定のユビキチン鎖を認識,結合することで,下流にシグナルを伝達する.異なるリシン残基で連結したユビキチン鎖はそれぞれ異なる立体構造をとるため,特異的なUBDによって認識されることが,ユビキチンコードの基本的な原理である2).
近年,ユビキチン鎖が枝分かれして連結する分岐鎖や,異なる連結型のユビキチン鎖が混在する混合鎖が発見された3, 4).加えて,ユビキチン自身がリン酸化やアセチル化といった翻訳後修飾を受けることも明らかになり,ユビキチンコードは従来考えられていた以上に複雑であることがわかってきた5, 6).均質なユビキチン鎖が8種類であるのに対し,分岐型ユビキチン鎖には原理的に28通りの組み合わせがあり,ユビキチンコードの機能的多様性を大幅に拡大する可能性が考えられる.本稿では,我々が最近報告した分岐型ユビキチン鎖およびアセチル化ユビキチンを中心に,ユビキチン鎖高次構造に関する最新の知見を紹介する.
1)分岐鎖の発見
従来,ユビキチンは鎖状に連結して均質な組成のポリユビキチン鎖を形成すると考えられてきた.しかしながら,ユビキチン分子内の複数のリシン残基が連結に用いられることで鎖が枝分かれする可能性については最近まで研究が進んでこなかった.
ユビキチン鎖の分岐についての解析が遅れてきた最大の要因は,検出手法が存在しなかったことにある.ユビキチン鎖のタイプの識別法としては主に,鎖特異的な抗体,あるいは質量分析法が用いられてきた7).しかし,鎖特異的な抗体では異なるユビキチン鎖が共存することは示せるが,分岐鎖なのかあるいは同一の基質が別々のユビキチン鎖で修飾されているのかを区別できない.また,質量分析法ではユビキチン鎖をトリプシンで切断し,ユビキチンC末端のGly75–Gly76が標的リシンに付加したペプチドを検出する.したがって,隣り合ったリシン残基で分岐する場合を除き,ペプチドが切断されてしまうため分岐という情報は失われてしまう.そのため,細胞内に分岐鎖がどの程度存在するのかについては長らく不明だった.Komanderらは自身の総説の中で分岐鎖を“Invisible chain”と表現している6).また,機能的意義の有無についても見解が分かれていたが,2014年以降,現在までに数種類の分岐鎖検出手法が考案され,細胞内に分岐鎖が存在することが明らかにされてきた.
1番目に,改変したユビキチンを細胞に発現させる手法である.Rapeらはユビキチン分子内にtobacco etch virus(TEV)切断配列を挿入し,ユビキチン鎖を精製後にTEVで切断することで分岐を検出する方法を考案した.さらに,この手法によりE3の一種であるanaphase-promoting complex/cyclosome(APC/C)がK11/K48分岐型ユビキチン鎖を形成することを報告した8).またLiらはこの手法を利用して,出芽酵母においてK29/K48分岐鎖が存在することを報告した9).
2番目に,我々は質量分析を用いたK48/K63分岐型ユビキチン鎖の検出・定量方法を考案した(3節参照).定量測定により,細胞内でK63鎖のおよそ20%はK48鎖で分岐しており,細胞内に分岐鎖が豊富に存在することを報告した10).さらに炎症性サイトカインinterleukin-1β(IL-1β)シグナルの下流で形成されるK63鎖がK48/K63分岐鎖を含むことを見いだした.
3番目に,特異的抗体を用いた方法である.DixitとRapeらは,K11鎖特異的抗体とK48鎖特異的抗体とをヘテロ二量体化した人工抗体を作製し,K11/K48混合鎖がある種の不良タンパク質の品質管理に関わっていることを報告した11).K11/K48抗体は分岐鎖のみならず混合鎖(K11鎖とK48鎖が直列に混在)をも認識する点は注意が必要である.また,KopitoらおよびFrydmanらは本手法を用い,K11/K48混合鎖がタンパク質品質管理に関与していることを報告した12, 13).
4番目に,ミドルダウン質量分析の利用である.ユビキチン鎖を非変性条件でトリプシン消化すると,球状で安定な構造をとるユビキチン部分は切断を受けず,連結部分であるArg74・Gly75間で切断される.Strieterらはこの性質を利用して部分切断されたユビキチンのミドルダウン分析により分岐鎖を検出できることを示した14).その後Komanderらは,トリプシンより特異的にArg74·Gly75間を切断するLbpro*を開発し,E3であるParkinが分岐鎖を形成することを示した15).
5番目に,混合鎖の検出方法として,Komanderらが報告したUbiCRest法が利用されている16).CohenらはUbiCRest法により,炎症性サイトカイン下流のシグナル伝達で形成されるユビキチン鎖がK63/M1混合鎖を含むことを報告した17).これらの分岐鎖・混合鎖の検出手法にはそれぞれ長所,短所があるが,さまざまな側面からの解析事例により,分岐鎖が細胞内で機能的であることが明らかとなってきた.
2)分岐鎖の形成機構と機能的意義
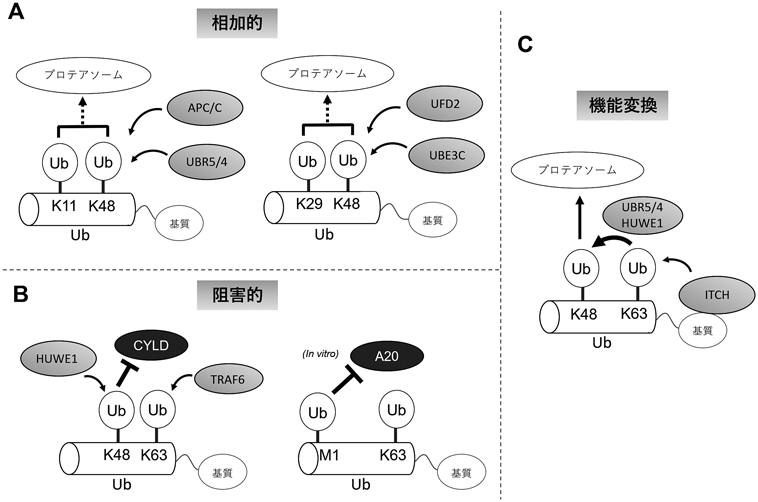
前項に述べた検出方法の開発により,分岐鎖の形成因子と形成機構の一部が明らかになってきた.その機構を大別すると,単一のE3による分岐鎖形成と,複数の鎖特異的なE3が協調的に作用することによる分岐鎖形成とに分けられる(図1).単一のE3としては,K11/K48分岐鎖を形成するAPC/Cが知られている.APC/Cはまず鎖特異性の低いE2であるUBE2Cと結合してK11, K48, K63鎖を形成する.次にK11鎖特異的なE2であるUBE2Sと結合し,先に存在しているユビキチン鎖にK11鎖を伸長する8).また,マイトファジーを制御するE3であるParkinは分岐鎖を形成する15).ParkinはK6, K11, K48, K63鎖を形成することがすでに知られているが18),分岐鎖がどのような連結タイプの組み合わせによって形成されているのかは現在のところ不明である.
一方,先に形成されたユビキチン鎖を認識して分岐を挿入する酵素も報告されている.我々は,IL-1βシグナルの下流でTRAF6がK63鎖を形成すると,HUWE1がK63鎖をK48で分岐させることを見いだした10).HUWE1はユビキチン結合ubiquitin-associating domain(UBA)およびubiquitin-interacting domain(UIM)を有しており,ユビキチン鎖に結合して分岐鎖を形成する.同様に,アポトーシス制御因子TXNIPのプロテアソーム依存性分解においては,基質を認識するE3であるITCHがK63鎖を形成,UBAドメインを有するE3であるUBR5がK63鎖からK48鎖を伸長することにより,K48/K63分岐鎖が形成される19).また,小胞体ストレス応答においてはKCMF1, UBE3Cが先に形成されているユビキチン鎖を分岐させ,それぞれK11/K48分岐鎖,K29/K48分岐鎖を形成する12).出芽酵母においてもUbr1, San1, Doa10, Hrd1およびUFD2がそれぞれK11/K48分岐鎖,K29/K48分岐鎖形成に関わっていることが報告された9, 13).
では,分岐鎖はどのような機能的意義を有しているのだろうか? 現在までに,いくつかのモデルが提唱されている.第一に,ユビキチンコードの合算である(図1A).K48鎖とK11鎖は両者ともプロテアソーム依存性タンパク質分解を誘導すると考えられており,K11/K48分岐鎖はユビキチンの密度を高めることでより強い分解シグナルとして働く11).またK63/M1混合鎖の場合,K63鎖はTAB2を,M1鎖はNEMOをそれぞれリクルートするため,炎症シグナル伝達を促進すると考えられている17).
第二に,ユビキチンコードの選択的な阻害である(図1B).我々は,K63鎖にK48分岐が形成されると,下流にシグナルを伝達するデコーダーであるTAB2のK63鎖への結合には影響を及ぼさないが,K63鎖に対するDUBであるCYLDの認識を阻害することでK63鎖切断を抑制し,K63鎖シグナルを安定化することでnuclear factor κB(NF-κB)シグナルを促進することを見いだした10).同様に,試験管内反応においてM1分岐はA20によるK63鎖切断を阻害する20).
第三に,ユビキチンコードの変換である(図1C).ユビキチンコードがシグナル伝達からプロテアソーム依存性分解へと変換される可能性は早期に指摘されていた21).我々は,K63鎖がK48分岐形成の足場になることで結果的にプロテアソーム依存性分解を促進する役割を有し,プロテアソーム非依存性のユビキチンコードであるK63鎖が分岐の文脈では機能を変換されることを見いだした19).
3)K48/K63分岐型ユビキチン鎖の分析法
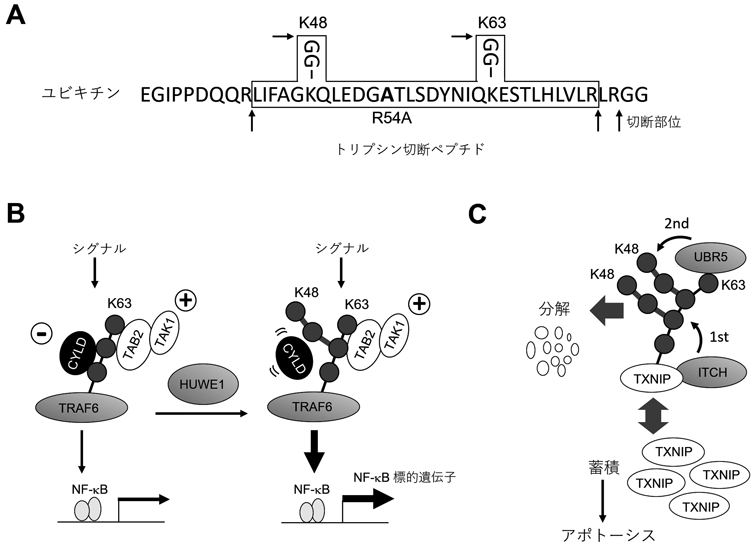
本稿では,定量的解析の進んでいるK48/K63分岐型ユビキチン鎖について紹介したい.我々は,細胞内で量的に最も主要であり,かつ機能的に大きく異なっているK48鎖とK63鎖に着目した.プロテアソーム依存性分解を制御するK48鎖と,プロテアソーム非依存的なシグナル伝達などを制御するK63が分岐によって連結したら,いかなる機能を有するだろうか? そこで,K48/K63分岐鎖を検出する手法を考案した.前述のように,質量分析法を用いたユビキチン鎖の定量では,トリプシン消化により修飾されていないリシン・アルギニン残基が切断され,ユビキチン鎖の高次構造に関する情報が失われてしまう.K48鎖とK63鎖が仮に分岐していた場合,この分岐鎖をトリプシン消化するとArg54で切断されるため,K48鎖とK63鎖が個別に検出される.したがって,分岐鎖なのか,個別の鎖が混在していただけなのかを判断することができない.そこで,ユビキチンのArg54をAlaに置換することによりトリプシン切断を受けないように改変すると,K48とK63の分岐鎖は単一のペプチドとして検出可能となる(図2A).また,この変異体を用いると「K48で分岐していないK63鎖(非分岐K63鎖)」「K63で分岐していないK48鎖(非分岐K48鎖)」を別々に検出できる.ここで,ユビキチンはほぼすべてのアミノ酸配列が進化的に保存されており,置換が機能的に許容されない可能性がある.しかし酵母においてユビキチンのすべてのアミノ酸残基を他のアミノ酸に置換した変異体の網羅的解析が報告されており,Arg54は置換しても生育に影響しない数少ないアミノ酸残基の一つであった22).またR54A置換が酵母の増殖,温度感受性とエンドサイトーシスに影響しないことも報告されている23).加えて,哺乳類培養細胞系およびin vitro系でのユビキチン鎖形成に影響しないこと,この研究で題材にした酵素群との反応性に影響しないことを確認した.
そこでR54A変異型ユビキチンを培養細胞に発現させると,K48/K63分岐鎖が容易に同定された.なお,切断に用いるトリプシン量を1/100まで減少させて部分消化を行うと,内在性のK48/K63分岐鎖(切断を免れたArg54を含む)も検出することができた.
次に,同位体標識した標準ペプチド(AQUAペプチド)を用いて分岐鎖の絶対定量を試みた.そのためには哺乳類培養細胞で野生型のユビキチンを変異体に置換する必要がある.Chenらの構築したユビキチン置換細胞は,内在性の四つのユビキチン遺伝子をテトラサイクリン誘導性にノックダウンし,同時に外来性ユビキチン遺伝子を発現させることができるため,ユビキチン鎖の機能解析に汎用されている24).この方法を用いて細胞内ユビキチンをR54A変異型に置換した.また非分岐K63鎖に関しては,ペプチドのN末端側がGln49であるため,トリプシン消化後に一部が自然に環状化してピログルタミン酸に変換され,精確な定量ができない.そこで,トリプシン消化後のペプチドをGln環状化酵素で反応させることですべてのN末端Glnをピログルタミン酸に変換した.またAQUAペプチドもN末端Gln49をピログルタミン酸に置換した配列を合成することで絶対定量に供することができた.
この手法により細胞全抽出液から内在性の分岐鎖を定量した結果,K48/K63分岐はすべてのK63鎖のおよそ20%程度存在するという結果を得た.さらにK48/K63分岐はプロテアソーム阻害剤の添加により全K63鎖の50%近くにまで増加した.ユビキチン鎖の細胞内での鎖長に関してはいまだ不明な点が多いが,定量結果を踏まえると,数個以上の長さのK63ユビキチン鎖であればK48/K63分岐を含んでいる可能性が高いことが予想され,分岐鎖はこれまで想定されていたよりも豊富に存在することが示唆された.
4)K48/K63分岐型ユビキチン鎖の細胞内機能
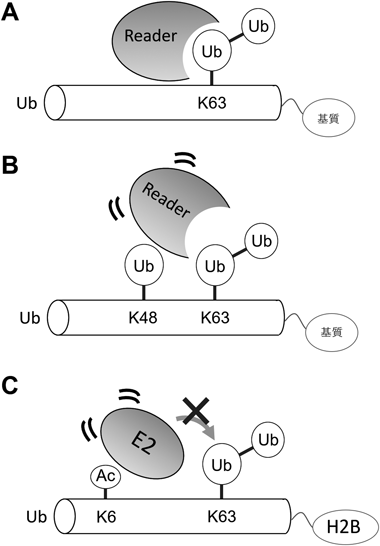
次に我々は,K48/K63分岐型ユビキチン鎖の細胞内機能を探索した.UBDによるユビキチンの認識には,主にIle44を中心とする疎水性パッチが相互作用表面として用いられる.ユビキチン鎖の連結タイプによって,疎水性パッチが並ぶ配向は異なってくるため,連結タイプに固有の相互作用表面が生成される.ここで,K48残基はIle44疎水性パッチの近傍に位置する.したがって,K48で分岐することが,K63鎖とUBDとの相互作用に影響を及ぼす可能性が考えられる.
そこで,K63鎖が制御する代表的な経路であるNF-κBと炎症シグナルに着目して以降の解析を行った(図2B).NF-κBは炎症応答をはじめさまざまなシグナル伝達を制御する転写因子である.炎症性サイトカインIL-1β応答の場合,ユビキチンリガーゼTRAF6が活性化され,K63鎖を形成する.ΤΑΒ2がK63鎖を認識して集積することでTAK1キナーゼが活性化され,linear ubiquitin chain assembly complex(LUBAC)依存的に形成されるM1鎖と協調的にIκBキナーゼ(IKK)複合体を活性化し,IκBの分解誘導によりNF-κBを活性化する25, 26).そこで細胞内でIL-1β依存的に形成されるTRAF6の自己ユビキチン化を定量したところ,K48/K63分岐鎖を含むことが明らかとなった.TRAF6自身はK63鎖を形成することが知られているため,K48分岐を形成する酵素を探索したところ,HECT型のE3であるHUWE1を同定した.HUWE1はin vitroにおいてTRAF6依存的に形成されたK63鎖にK48分岐を付加することでK48/K63分岐鎖を形成した.
次にHUWE1の細胞内機能を検証するため,HUWE1をノックダウンしたところ,内在性のK48/K63分岐鎖の減少に加え,IL-1β·TRAF6依存的なNF-κB活性が抑制された.この結果から,K48/K63分岐鎖はNF-κBシグナル伝達を促進していることが示唆された.そこでこの分子機構を探索するために,K63鎖のデコーダーであるTAB2およびK63鎖のDUBであるCYLDとK48/K63分岐鎖との相互作用を検討した.ユビキチンK48残基はCYLDがK63鎖を認識する際の相互作用表面近傍に位置し,遠位側ユビキチンのK48残基はCYLDの表面に埋まっている27).検討の結果,TAB2はK63鎖およびK48/K63分岐鎖に対して同等に相互作用したのに対し,CYLDによるK63鎖の切断はK48/K63分岐鎖において著しく抑制されることが判明した.すなわち,ユビキチン鎖の分岐は結合因子を特異的に排除あるいは許容することで,均質なユビキチン鎖とは異なる新たなシグナルとして機能することが示唆された(図2B).
では,K48/K63分岐鎖はプロテアソーム依存性分解に関与するのだろうか? TRAF6は活性化の後,CYLDやA20によって迅速に脱ユビキチン化され,炎症シグナルを一過性にとどめるよう制御を受けている.そのため,TRAF6のIL-1β刺激後の分解は観察されない.より一般的な分解基質であればK48/K63分岐鎖がプロテアソーム依存性分解を制御する可能性が考えられる.そこで,細胞内でプロテアソームに結合しているユビキチン鎖を精製し,定量分析に供したところ,K48/K63分岐鎖はK48鎖同様にプロテアソームに濃縮していることが明らかとなった19).一方,非分岐のK63鎖はこれまでの報告どおりプロテアソームには濃縮されなかった.そこで,K48/K63分岐鎖によって分解を制御される基質を探索した結果,アポトーシス制御因子であるTXNIPを見いだした(図2C).解析の結果,TXNIPのプロテアソーム依存性分解にはK63鎖特異的なE3であるITCHとK48鎖を形成するE3であるUBR5/UBR4/HUWE1の両方が必要であることがわかった.すなわち,K63鎖とK48鎖は各々が固有の役割を担っており,単独では分解を促進できない.試験管内反応で詳細に解析したところ,ITCHは基質認識を担っており,TXNIPに直接相互作用してK63鎖を形成すること,UBR5は単独ではTXNIPをユビキチン化できないが,形成されたK63鎖に対してK48分岐を形成し,さらにK48鎖を伸長することが明らかとなった.すなわち,K63鎖はそれ自体がプロテアソーム経路への運搬シグナルとしては機能しないが,特異的な基質にK48鎖を誘導するための目印として機能している.
以上の結果を合わせて,K48/K63分岐鎖は均質なユビキチン鎖の足し算ではなく,固有の機能を有する新たなユビキチンコードであることが明らかとなった.
1)アセチル化ユビキチンの定量解析
近年,ユビキチン自身が翻訳後修飾を受けることで新たなユビキチンコードとして機能する事例が明らかとなってきた.本節では,我々が解析してきたアセチル化ユビキチンの機能について紹介する28).
ユビキチン修飾の特徴的な点として,タンパク質性の翻訳後修飾であることがあげられる.そのため,修飾分子であるユビキチン分子自身が翻訳後修飾を受ける可能性が考えられたが,当時そのような事例はほとんど未知であった.そこで,ユビキチンに対する翻訳後修飾の探索を行った.測定はターゲットプロテオミクスの手法であるparallel reaction monitoring(PRM)を用いた29).ショットガン解析での予備検討からアセチル化とリン酸化の存在が予想されたので,すべての標的残基に対するアセチル化とリン酸化の理論値を設定して測定を行った.哺乳類培養細胞から基質に付加された内在性ユビキチンおよびユビキチン鎖を精製し,測定に供した結果,複数のアセチル化サイト,リン酸化サイトを同定した.AQUAペプチドを用いた定量の結果,リシン6番のアセチル化(K6Ac)はtrichostatin A(TSA)などの脱アセチル化阻害剤に感受性であり,細胞内で酵素的に制御されていることが示唆された.
2)アセチル化ユビキチンの分子作用機構
同定した翻訳後修飾の中で,K6およびK48のアセチル化に着目して以降の解析を行った.両残基は,ユビキチンとデコーダーとの相互作用に汎用されるIle44疎水性パッチの近傍に位置するため,機能的な影響が大きいと予想されたためである.機能解析を進める上で,目的のリシン残基特異的にアセチル化修飾を施したリコンビナントのアセチル化ユビキチンを使用した.検討の結果,アセチル化ユビキチンはin vitroでのE1, E2への連結や,ヒストンのモノユビキチン化反応には影響を及ぼさなかった.しかし,E2酵素によるユビキチン鎖の伸長はアセチル化ユビキチンによって著しく抑制された.定量解析の結果,ほぼすべての連結タイプのポリユビキチン鎖形成がK6アセチル化またはK48アセチル化ユビキチンによって抑制された.
E2酵素はユビキチン鎖伸長の際にユビキチンと非共有結合的に相互作用することが知られている.そこで,鎖形成機構が詳細に解明されているE2酵素であるUbc13-Uev1aを用いてさらに検討を行った.Uev1aはアクセプター(修飾される側)のユビキチンとIle44疎水性パッチで相互作用し,そのK63残基をUbc13の活性中心近傍に位置させることでK63連結を形成する.この相互作用にはユビキチンのK6およびK48残基が関わっており,リシン残基のアセチル化により正電荷が中和され,相互作用が失われることが判明した.
さらに,内在性のアセチルユビキチン化基質をプロテオミクス解析によって探索した結果,ヒストンH2Bを同定した.H2BのK120モノユビキチン化は転写活性化に関わるヒストン修飾であることが知られている.そこでH2BのK120モノユビキチン化の変動を検討したところ,アセチル化をミミックしたユビキチンによってK120モノユビキチン化が安定化されることが判明した.これらの結果から,翻訳後修飾分子であるユビキチン自身がアセチル化修飾によって制御されることが明らかとなった.
本稿で紹介したアセチル化ユビキチン以外にも,リン酸化やADPリボシル化といった低分子化学修飾によるユビキチンの機能制御が報告された5, 30).ユビキチンの翻訳後修飾はごく微量であることがわかっており,特定の基質や局在において濃縮される,あるいは特定のシグナルによって活性化される未知の翻訳後修飾の発見が期待される.