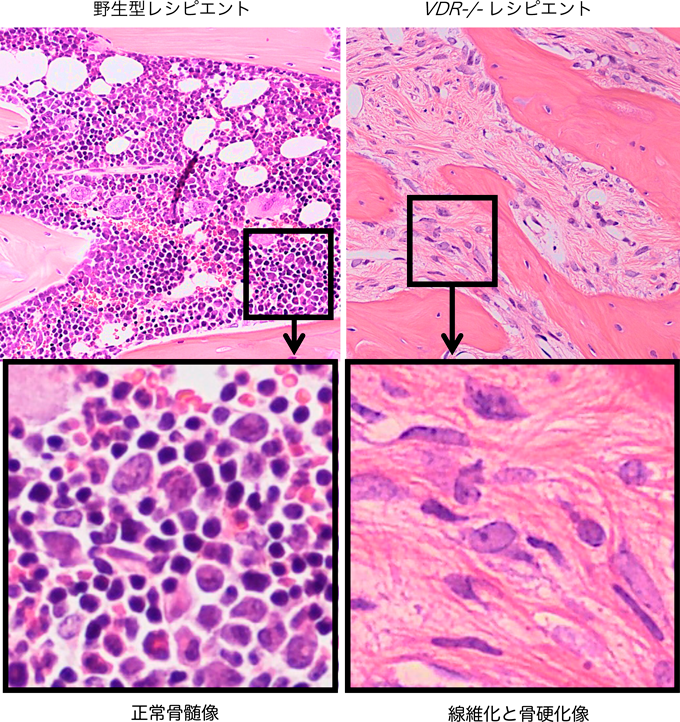
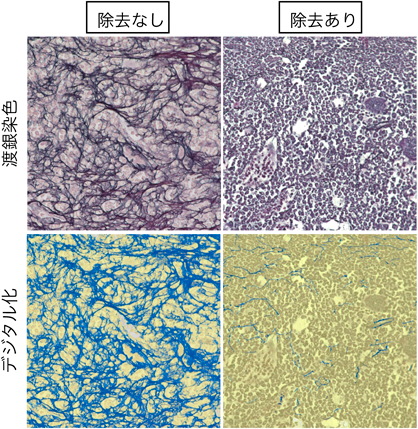
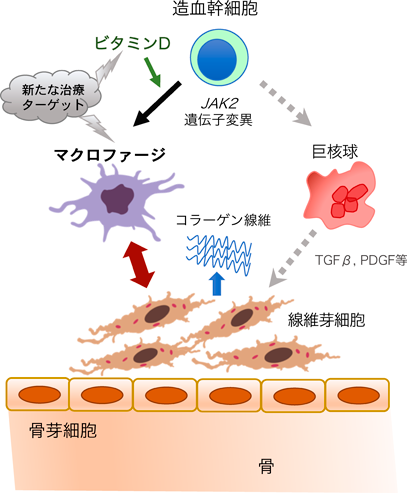
ビタミンDとマクロファージによる骨髄線維症の病態制御Vitamin D and Macrophages regulate Myelofibrosis
神戸大学大学院医学研究科血液内科学Hematology, Kobe University Graduate School of Medicine, Kobe, Japan ◇ 〒650–0017 兵庫県神戸市中央区楠町7–5–1 ◇ 7–5–1 Kusunoki-cho, Chuo-ku, Kobe Hyogo 650–0017 Japan
発行日:2020年2月25日Published: February 25, 2020