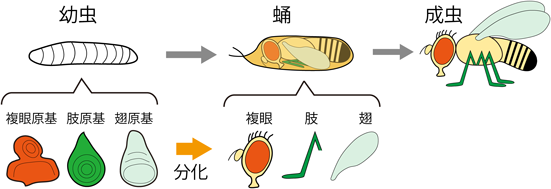
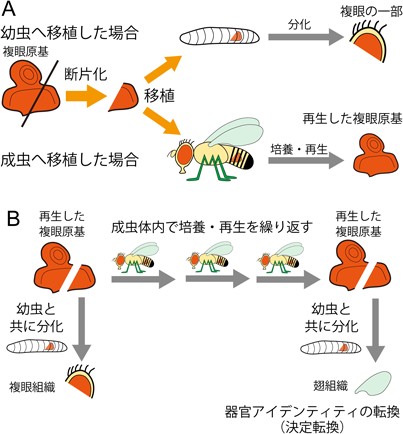
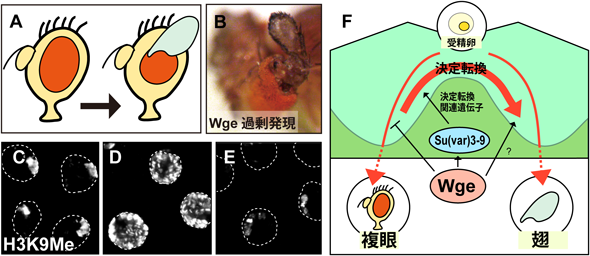
ショウジョウバエ成虫原基の器官運命決定転換におけるクロマチン制御機構Cell fate plasticity and chromatin regulations;a view from Drosophila imaginal disc transdetermination
東北大学大学院薬学研究科生命機能解析学分野Molecular Genetics, Graduate School of Pharmaceutical Sciences, Tohoku University ◇ 〒980–8578 宮城県仙台市青葉区荒巻字青葉6–3 ◇ Aoba 6–3, Aramaki, Aoba-ku, Sendai, Miyagi 980–8578, Japan
発行日:2020年2月25日Published: February 25, 2020