MCM研究の歴史と最近の発展History and recent progress of MCM research
元茨城大学理学部教授
受付日:2019年10月18日Received: October 18, 2019
発行日:2020年4月25日Published: April 25, 2020
MCM2-7タンパク質はDNA巻き戻し酵素(DNAヘリカーゼ)として真核細胞DNA複製に中心的な役割を果たす.MCM2-7複合体(MCM2, 3, 4, 5, 6, 7タンパク質からなるヘテロ六量体)のDNAヘリカーゼの活性化にはCDC45とGINSなどの数種のタンパク質の働きが必要である.研究の発展の一つは,電子顕微鏡観察による,分子モーターとしてのMCM2-7ヘリカーゼの理解である.他のホモ六量体のDNAヘリカーゼとは異なり,MCM2-7の各タンパク質は独自の役割を果たす.DNA合成との協調などのDNA複製フォークにおけるMCM機能制御には数多くのタンパク質が関わる.DNA複製フォークにおいては,DNAが複製されるだけでなく,ヌクレオソーム構造も再構築される必要がある.MCM2は,古いヒストンH3/H4の複製後DNAへの結合に中心的な役割を果たす.
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
MCM(minichromosome maintenance)2-7タンパク質はDNA巻き戻し酵素として真核細胞DNA複製に中心的な役割を果たす.出芽酵母MCMの遺伝子が明らかになって約30年が経過した現在,MCM研究の中心は,DNAと結合したCMG(ヘテロ六量体であるMCM2-7にCDC45とGINSが結合した複合体)の構造解析,CMGヘリカーゼ活性化機構および複製フォークでの古いヒストンH3/H4の分配機構についてである.構造解析については,ATPの結合と加水分解に伴い,一本鎖DNA上での移動を引き起こすMCM2-7の構造変化の理解が焦点になる.これまでに解析されたホモ六量体ヘリカーゼにはない複雑さが予想されるが,ヘリカーゼ機能におけるMCM2-7各タンパク質の役割を理解する必要がある.鋳型上の古いヒストンH3/H4の複製後DNAへの分配機構の理解は,細胞複製の理解に欠かせないばかりでなく,細胞分化(非対称分裂)の理解につながる発展性を持つ.本稿ではMCMについて,MCM遺伝子とヘリカーゼ活性の発見の経緯,CMG複合体の構造解析,ヒトMCM4の機能,DNA複製フォークでのヒストン集合,細胞老化との関わりの順に述べる.
Tyeの研究室において,高温で小染色体(minichromosome)の保持に欠陥のある出芽酵母変異体16相補群が得られた1).1990年にその中の一つの責任遺伝子であるMcm3遺伝子が同定された2).Botsteinの研究室では1991年に,細胞周期のS期進行に欠陥を持つCDC変異体の中で,CDC45変異体と遺伝学的に相互作用するCDC46, CDC47とCDC54変異体が同定された3).1992年に,Kearseyのグループは,NasmythとNurseが分離した分裂酵母変異体4)の中のCdc21変異体の表現型を解析した5).Yanagidaの研究室は,分裂酵母の細胞周期進行に支障を来す変異体を分離し,1993~1994年にNda4とMis5遺伝子を同定した6, 7).1991年にTyeの研究室は,Mcm3とMcm2の遺伝子産物には,配列に共通な部分があり,さらに,それらと出芽酵母CDC46と分裂酵母Nda4にも共通部分があることを示した.そのことから,酵母MCM遺伝子にはファミリーが存在することが明らかになった8).
1993年に,Knippersの研究室のHuらはMCMに共通して存在する部分を認識する抗体を使って,ヒト細胞にMCMタンパク質が存在することを示した9).Kimuraらは,DNA合成酵素αと相互作用するマウスP1タンパク質(MCM3)の細胞周期での局在変化を調べた10).1996年に,TodorovらはヒトMCM2タンパク質が細胞のDNA複製に重要な役割を示すことを報告した11).この1996年には,MCM2-7の各メンバーが酵母,マウスとヒト細胞で同定されている(表1).一方で,1995年に,Takisawa12),Blow13)およびLaskey14)の各研究室から,アフリカツメガエル卵抽出液を使ったDNA複製系で提唱された概念であるDNA複製許可(細胞周期で一度のみのDNA複製を許可する)について,それをつかさどる因子の中にMcm3などが含まれることが示された.このことによりDNA複製許可と酵母で遺伝学的に同定されたMcmとが結びついた.
| MCM2 | MCM3 | MCM4 | MCM5 | MCM6 | MCM7 | |
|---|---|---|---|---|---|---|
| S. cerevisiae | Mcm2 Sinha et al. (1984) | Mcm3 Gibson et al. (1990)2) | CDC54 Hennessy et al. (1991)3) | CDC46 Hennessy et al. (1991)3) | Mcm6 | CDC47 Hennessy et al. (1991)3) |
| S. pombe | Nda1 Miyake et al. (1993)6) | Mcm3 | Cdc21 Coxon et al. (1992)5) | Nda4 Miyake et al. (1993)6) | Mis5 Takahashi et al. (1994)7) | Mcm7 |
| ヒト/マウス | BM28 Todorov et al. (1994)11) | P1 Kimura et al. (1994)10) | CDC21 Kimura et al. (1995) | CDC46 Kimura et al. (1995) | ラットタンパク質 Sykes & Weiser (1995) | p85MCM Schulte et al. (1996) |
| 出芽酵母,分裂酵母,およびマウス/ヒトでのMCM2-7遺伝子について,初期の遺伝子名と発表論文を示す. | ||||||
1993年に,KooninはMCMタンパク質にはATP結合モチーフがあることを指摘し,DNAヘリカーゼとして複製に機能することを示唆した(図1)15).1997年に,Yanagida研のAdachiらは分裂酵母のMCM2-7ヘテロ六量体を調製し,ヘリカーゼ活性を検出できないことを報告した16).1996年に私のグループは,ヒトHeLa細胞抽出液を0.35 M NaCl存在下でヒストンカラムにかけたところ,MCM3, 5タンパク質は結合しないが,MCM2, 4, 6, 7タンパク質は強く結合し,単一にまで精製されることを見つけた17).そこには,MCM2/4/6/7の四量体とMCM(4/6/7)2の六量体の2成分が含まれていた.翌年,MCM(4/6/7)2が,一本鎖DNA依存的ATP加水分解活性と3′→5′の方向性を示すDNAヘリカーゼ活性を発揮することを報告した18).MCM(4/6/7)2にMCM2あるいはMCM3/5の二量体を加えると,ヘリカーゼ活性は阻害されたことから,MCM4, 6, 7がヘリカーゼ機能に触媒作用を,MCM2, 3, 5が制御作用を持つことが考えられた19, 20).2001年にSchwachaとBellは出芽酵母MCM2-7ヘテロ六量体がATP加水分解活性を発揮すること,さらに,ATP結合部位に変異を導入したMCM2-7の分解活性を調べ,その活性発揮においてMCM4, 6, 7が触媒作用を,MCM2, 3, 5が制御作用を担うことを報告した21).彼らの研究から,特にMCM4とMCM7がATP加水分解活性に決定的な役割を果たすことが示された22).
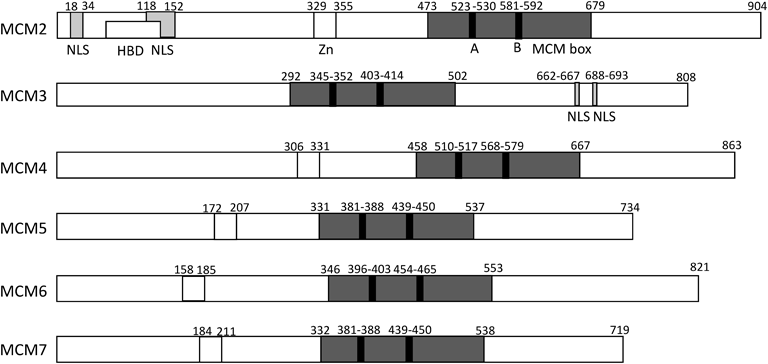
ヒトMCM2-7タンパク質の一次構造の特徴を示す.保存性の高いMCMボックスが灰色で示され,その中のATP結合モチーフであるWalker AとBが黒色で示されている.他に,NLS:核局在化配列,Zn:Znフィンガー,HBD:ヒストン結合ドメインが示されている.数字はアミノ酸番号である.
2000年にBlowの研究室は,アフリカツメガエル卵抽出液からいくつかのMCMサブ複合体を調製し,それらからMCM2-7の六量体形成に至る経路を明らかにするとともに,DNA複製を誘導できるのは,サブ複合体ではなく,ヘテロ六量体であることを示した23).同じ年に,LabibとDiffleyは,出芽酵母DNA複製の開始だけでなく進行にも,すべてのMCM2-7タンパク質が必要であることを示した24).これらの結果から,複製の開始と進行にはMCM2-7の六量体が機能することがわかり,MCM(4/6/7)2複合体が複製の進行に関わる可能性が否定された.つまり,MCM(4/6/7)2複合体はMCM複合体の精製の過程で生成されたと考えられる.MCM2-7の六量体は,リング状の構造を形成するが,それらは,N末端側から見たときに,時計回りにMCM2-6-4-7-3-5の順に配置する25).ただし,MCM2と5の間は安定な結合をとらず,そこでリングが開閉する(図2)22).また,それ以外のMCM間の結合も同等でないことが生化学的な解析からわかっている.つまり,MCM6/4/7がコア複合体を形成し,それにMCM2が結合することでMCM2/6/4/7の四量体が形成される26, 27).MCM3と5は安定な二量体を形成し,それが弱くMCM2/6/4/7複合体に結合することでMCM2-7の六量体が形成される.このことは,細胞質で形成したMCM2/4/6/7四量体はMCM2のもつ核局在化シグナルによって核移行する一方,MCM3/5二量体はMCM3の核局在化シグナルで核移行し,これら二つの複合体が核内で会合するという知見と一致する28, 29).植物の細胞ではMCM5がMCM7より低い塩濃度でクロマチンから遊離する30).このことは,以下に述べるように,CMG複合体の中でMCM4, 6, 7のみがDNAに直接的に結合するというクライオ電子顕微鏡解析の結果と矛盾しない.以上の結果から,MCM4, 6, 7とMCM2, 3, 5は,MCM2-7ヘリカーゼ機能において異なる働きを発揮することが示唆される.最近,我々は,ヒトHeLa細胞でのMCM2, 3あるいは5の過剰発現が,DNA複製異常から生じると考えられる核形態異常をもたらすことを見いだした31).この結果は,MCM2, 3と5はMCM2-7ヘリカーゼ機能に制御的な作用を発揮するという知見と一致する.
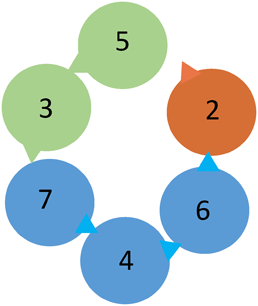
真核細胞のMCM2-7をそれぞれのN末端側から見たときの配置を示す.各サブユニット間の結合には,三角で示す各MCM内のATP結合部位が関わる.ヘリカーゼの触媒サブユニットと考えられるMCM4, 6, 7を青色で,制御サブユニットと考えられるMCM3, 5を緑色で,制御の役割に加えヒストンシャペロンとして機能するMCM2を燈色で示した.
これまでにMCM2-7ヘテロ六量体を含むさまざまなMCM複合体のうち,MCM(4/6/7)2とMCM4/7複合体のみがDNAヘリカーゼ活性を発揮することが示された32).2008年にBochmanとSchwachaは,反応液中にグルタミン酸や酢酸の陰イオンが含まれた特殊条件下で,出芽酵母MCM2-7の六量体がDNAヘリカーゼ活性を発揮することを報告した33).DNA複製の開始に機能するCDC45はMCM2-7と直接的に結合する.Arakiの研究室において,酵母遺伝学を使って,タンパク質四量体であるGINSが新たに発見された34).GINSもMCM2-7に結合する.Labibの研究室では,DNA複製の際にCDC45とGINSはMCM2-7と一緒に働くことが示唆された35).2006年に,Botchanの研究室では,ショウジョウバエの細胞抽出液から,CDC45-MCM2-7-GINSからなるCMG複合体を精製し,それが通常の反応条件下でDNAヘリカーゼ活性を発揮することが示された36, 37).Hurwitzの研究室では,ヒトCMG複合体が比較的長い二本鎖DNAを巻き戻し,DNA合成酵素εによるDNA合成を促進することが示された38).ショウジョウバエCMG複合体でMCM2-7の各ATP結合部位に変異を導入したものを調製し,そのヘリカーゼ活性を調べる実験から,MCM2とMCM5の結合とMCM5とMCM3の結合に関わるATP結合部位が,活性発揮に必須であることが示された37).この結果は,MCM2-7のDNAヘリカーゼ活性発揮において,MCM7のATP結合部位が必須の役割を果たすことと対照的であり,CMG中のMCM2-7はATP結合性において非対称であるといえる.ショウジョウバエCMG複合体において,CDC45はMCM2とMCM5に結合し,GINSはMCM5とMCM3に結合する39).これらの結果から,CDC45とGINSの結合により,CMG複合体中のMCM2, 3と5の構造変化が起こることでDNAヘリカーゼ活性が発揮されると考えられる.Enemarkの研究室のMeagherらによる報告では,古細菌Sulfolobus solfataricus MCMの構造解析を基に真核細胞MCM2-7の構造を分析し,MCM2, 3と5のそれぞれに存在するDNA結合ループであるH2I(helix-2-insert)ヘアピンがDNA結合を許容しない構造をとり,GINSおよびCDC45の結合により初めて,ATP,さらにはDNAと結合することが提唱されている40).
出芽酵母のG1期クロマチンから調製したMCM2-7複合体のクライオ電子顕微鏡による構造解析から,古細菌MCMと同様に各MCMに,Ext(external),H2I,そしてPS(presensor)1などのヘアピン構造が存在することがわかった41, 42)(図3).これらのヘアピンは,DNAヘリカーゼ活性が発揮されるときに,一本鎖DNAとの相互作用に関与する.ATP結合,分解と解離に伴い,ヘアピン構造が変化することでDNAとの結合が変わり,MCMのDNA上での移動が起こると考えられる.また,ACL(allosteric communication loop)と呼ばれるループ構造は,隣接するMCMサブユニットとの結合に関わる43).N末端領域に存在するZnフィンガーモチーフは,マウスMCM(4/6/7)2ヘリカーゼ活性発揮における役割は認められなかったが44),古細菌のMCMヘリカーゼ活性には必要であった45).さらに,このモチーフは古細菌Methanobacterium thermoautotrophicum MCMの六量体の二量体化に必要で46),酵母の増殖に必要であることがわかった47).細胞内のDNA複製起点で起こるMCMの二量体化については,それぞれがゲノムの逆方向へ移動することで両方向へのDNA複製が始まると考えられる.低加水分解型ATP類似体存在下でヘリカーゼ基質DNAと結合したショウジョウバエCMGのクライオ電子顕微鏡解析から,MCM4, 6, 7の各サブユニットのPS1ヘアピン内の塩基性アミノ酸とDNAの一本鎖部分が結合することが示された48)(図3).かたや,O’Donnellの研究室は,出芽酵母CMGを用いて同様の解析を行い,MCM2, 3, 5と6のH2IとPS1がDNAと結合することを報告した49).また,出芽酵母DNA複製起点に結合した複製起点認識複合体(origin recognition complex:ORC),Cdt1とMCM2-7複合体の構造解析では,MCM4, 6と7のPS1ヘアピンにDNAが結合することが示された50).さらに,出芽酵母複製起点に結合した二量体化したMCM2-7の六量体の構造解析では,MCM3, 4, 6,と7がDNAに結合すると報告された51).ところで,Froelichらは,N末端領域に存在するMSSB(MCM single-stranded DNA binding motif)と呼ぶ進化的に保存された二つの塩基性アミノ酸が,古細菌MCMの一本鎖DNA結合に必要であることを示した.この二つのアミノ酸は,MCM4, 6, 7のみにそろって存在し,それらは酵母の増殖に必要である52).以上の結果から,MCM2-7複合体の中のMCM4, 6, 7が主にDNAとの結合に関わると考えられる(図4A).一方で,最近,ATP存在下において一本鎖DNA上を移動するショウジョウバエCMGが調製され,その構造解析から,MCM2-7とDNAとの結合において4種の異なる構造が同定された.著者らは,ATP結合によりすべてのサブユニットが順次DNAと相互作用し,MCM2-7リングが回転しながらDNA上を移動するモデルを提唱している(図4B)53).前出のMeagherらの論文でも,同様の回転モデルが提唱されている40).MCM2-7の中でMCM4, 6, 7はDNAと安定に結合できるが,ATP存在下にCMG複合体がヘリカーゼ活性を発揮しているときには,ATPとの結合,加水分解と解離に伴い,すべてのサブユニットが順次DNAと相互作用するのかもしれない.

MCM2-7タンパク質に共通して存在する3種のヘアピン構造のExt, H2IとPS1と,ACLの場所をMCM4内に示す.MCM4のPS1内の600位のリシン,MCM6の486位とMCM7の471位のリシンが一本鎖DNAと結合することが,ヘリカーゼ基質に結合したショウジョウバエCMGのクライオ電子顕微鏡観察により示されている48).MCM4内のMSSBと呼ばれる291位のアルギニンと353位のリシンは,DNAヘリカーゼ機能において一本鎖DNAとの結合に関わると考えられる52).MCM6とMCM7内のMSSBも示す.MCM4の1~120位の領域はCDKなどによりリン酸化を受ける.
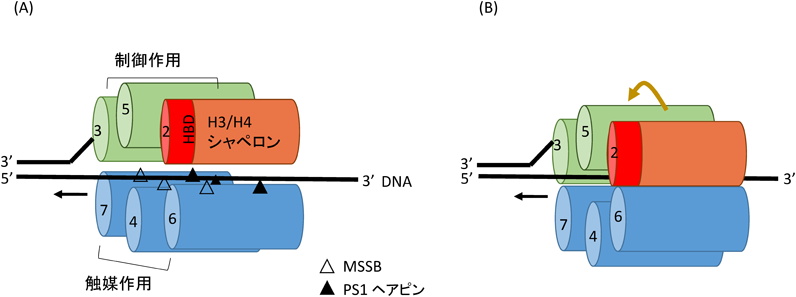
(A)CMG複合体中のMCM2-7タンパク質のDNAとの結合のモデルを示す.MCM4, 6, 7のPS1ヘアピンとMSSBがヘリカーゼ機能の際に一本鎖DNAとの結合に関わる可能性がある48, 52).いくつかの実験結果から,MCM4, 6, 7はDNAヘリカーゼ機能に触媒の役割を,MCM2, 3, 5は制御の役割を果たすことが示唆される.また,MCM2はDNA複製フォークで新旧のヒストンH3/H4の複製後DNAへの結合に関わる.(B)ヘリカーゼ活性を発揮するCMG複合体中のMCM2-7の回転モデル.六量体リング構造をとるMCM2-7タンパク質はATP存在下に回転しながら一本鎖DNA上を移動する.このとき,MCM2-7の各タンパク質は順次DNAと結合する53).
ヘリカーゼ基質に結合した出芽酵母CMGの構造解析から,リングを形成するMCM2-7の六量体中の各MCMタンパク質の配置については,N末端部分がヘリカーゼ移動方向の先頭に,C末部分が後方であることが示された49).しかし以前に,Costaの研究室からは,基質に結合したショウヨウバエCMGの構造解析から,逆のMCM2-7の配置が示されていた48).出芽酵母CMGでのMCM2-7のDNA上の配置は,5節で述べるように,MCM2のN末端部分が,鋳型DNAに結合した古いヒストンH3/H4の,複製したラギング鎖側DNAへの移行に関わるという知見と矛盾しない.前出の,Costaの研究室によるATP存在下において一本鎖DNA上を移動するショウジョウバエCMGの構造解析からは,MCM2-7の各N末端部分が,移動するDNAの先頭に配置される配向が示されている53).DNAの巻き戻し機構については,単純なDNA鎖排除機構(strand exclusion)が支持されている54).その根拠は,ヘリカーゼ基質DNAの二本鎖部分に小タンパク質を化学架橋したものを用いると,CMG複合体が移動するDNA側に小タンパク質が結合しているとヘリカーゼ活性は阻害されるが,逆側のDNAに小タンパク質が結合していても活性は阻害されない,という実験結果である.基質DNAの一本鎖部分と二本鎖部分の境界で,CMGのリング内に二本鎖DNA部分が入る場合には,逆側の鎖に結合した小タンパク質もヘリカーゼ活性を阻害するはずである.ショウジョウバエCMGによるDNA巻き戻しの動力学を,一分子磁性ピンセットを用いて解析すると,CMGは,偏りを持ったランダムな移動性を示し,さらに一方方向のみの移動でないことが示された.著者らは,CMGはATP加水分解に伴い,怠惰なブラウン運動をする鋸歯車(lazy Brownian Ratchet)であると表現する55).
数多くのタンパク質がMCM2-7に結合することで,そのヘリカーゼ機能が制御されている.特異的な相互作用が,FACT-MCM4間56),DNA組換えタンパク質TONSL/MMS22L-MCM5間57),アンドロゲン受容体-MCM7間58),DNA組換え修復タンパク質FANCD2-MCM4, 5, 7間59),separase-MCM4, 6, 7間60),MCM10-MCM2, 6間61)で知られている(図5).DNA複製フォーク保護因子であるTIMとTIPINは,MCM3-7に結合するが,特にTIMはMCM4と,TIPINはMCM3, 7との親和性が高い62)(図5).最近のCostaの研究室からの報告では,TIM, TIPINとClaspinからなるフォーク保護複合体は,MCM2-7のN末端側に結合することが示されている53).MCM3/5はSTAT1α63)とRNA合成酵素II64)に結合することから,転写にも関わると考えられる.これらの分子間相互作用の機能的な理解には,MCM2-7ヘリカーゼ機能におけるMCM2-7各タンパク質の独自な働きを理解する必要があると考える.
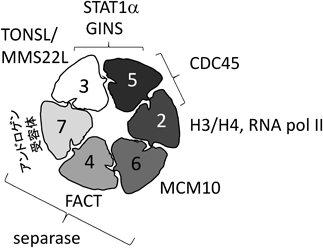
数多くのタンパク質とMCM2-7との結合は,MCM2-7の機能制御に関わると考えられる.MCM2-7との特異的な相互作用が,FACT56),RNA合成酵素II64),TONSL/MMS22L57),MCM1061),アンドロゲン受容体58) とH3/H478, 82) で同定されている.また,いくつかのMCM2-7サブユニットとの結合がわかっているのは,separaseとMCM4, 6, 760),そしてStat2とMCM3, 5の間である.また,FANCD2はMCM4, 5, 7に結合する58).TIMとTIPINは,それぞれ主に,MCM4とMCM3, 7に結合する62).CMG複合体中,CDC45はMCM2とMCM5に,GINSはMCM3とMCM5に結合する39).
マウスMCM(4/6/7)2複合体において,各MCMのATP結合部位変異体の解析から,MCM6は高い親和性でATPと結合することにより,またMCM7はATP加水分解活性により,ヘリカーゼ機能に必須の役割を果たす44).MCM4は一本鎖DNA結合に役割を果たすと考えられる.最近,筆者らは,ヒトMCM4のアミノ末端部位のアミノ酸番号10と11のアルギニンがMCM(4/6/7)2複合体のDNAとの結合性を高めることで,ヘリカーゼ活性に促進的に働くことを報告している65).また,酵母MCM4のアミノ酸番号180位(ヒトMCM4では160位)付近の領域がMCM2-7の二量体化とともにDNAの巻き戻しに必要であることが示された66).
ヒトMCM4は細胞周期に依存してリン酸化される.サイクリン依存性キナーゼ(cyclin-dependent kinase:CDK)によりリン酸化されうる12か所の部位がN末端部位に存在することから,CMGのヘリカーゼ機能はCDKにより制御されると考えられる67)(図3).アフリカツメガエル卵抽出液を使った複製系での初期の研究において,CDKによるMCM4リン酸化がMCMのクロマチン結合を阻害することで,MCMのDNA複製ライセンス機能を阻害することが報告された68, 69).我々は,CDKがヒトMCM4の特異的部位(3, 7, 19, 32, 54と110位のセリンあるいはトレオニン残基)をリン酸化することで,MCM(4/6/7)2複合体のヘリカーゼ活性を阻害することを報告した.このときに,複合体の一本鎖DNAへの結合性が低下することを認めている70, 71).最近,我々は,この6部位のすべてをリン酸化を模擬するグルタミン酸に置換したMCM4の存在により,MCM(4/6/7)2およびMCM2-7複合体形成が阻害されることを見いだした65).このグルタミン酸置換変異MCM4のMCM複合体形成に対する影響は,上記のCDKによるMCM4リン酸化の効果を合理的に説明することができる.よって,細胞周期のG2/M期における,CDKによるMCM4の高度のリン酸化は,CDKによるCDC6, CDT1とORCのリン酸化とともに,DNA複製の再開始の抑制に関わると考えられる72).
DNA複製の開始にはDbf4依存キナーゼ(Dbf4-dependent kinase:DDK)によるMCMのリン酸化が必要である.DDKによるMCM4のリン酸化はCDC45のクロマチン結合を促進する73).また,MCM2とMCM4に対してCDKとDDKによる協調的リン酸化が部位特異的に起こると考えられ74),それらは複製起点でのタンパク質集合に関わる可能性がある67).
DNA複製の進行において,数多くのタンパク質が複製フォークに存在する75).それらの中で,Ctf4/AND1はCMGによるDNA巻き戻しとDNA合成酵素によるDNA合成とを協調させる働きを持つ76, 77)(図6A, B).DNA複製フォークでは,DNA鎖が複製されるだけでなく,ヌクレオソーム構造も作られる必要がある.一つの細胞が同じ機能を持つ二つの細胞に増えるときに,鋳型DNAに結合していた古いヒストンは複製後の2本のDNA鎖に均等に分配される必要がある.つまり,遺伝子発現を制御する化学修飾されたヒストンH3/H4は両DNA鎖に均等に分配されなければならない.そこにさまざまなヒストンシャペロンが機能する(表2).MCM2のN末端部分のヒストン結合ドメイン(HBD)は,ASF1とFACTとともに,古いヒストンH3/H4(二量体)に結合することで,その再結合過程の初期に関わる56, 78–82).よく知られたH3/H4シャペロンであるCAF-1は,古い四量体H3/H4の複製後DNAへの結合の最終段階に機能すると考えられている83).試験管内でのクロマチン構造のDNAヘリカーゼ反応系において,FACTによる活性促進が示されている56).よって,FACTとCMG中のMCM2がまず,DNA複製フォークで鋳型DNA上のヌクレオソーム構造を破壊するのかもしれない.この反応で遊離した古いH3/H4はMCM2のヒストン結合ドメインに保持され,それにASF1が結合し,ASF1-H3/H4-MCM2の複合体が形成される可能性が考えられる84, 85).その後H3/H4は,ASF1とCAF-1との親和性により,CAF-1に移され,CAF-1-(H3/H4)2ができる.(H3/H4)2が複製後DNAに結合することで,ヌクレオソーム構造の核が構築されると考えられる.別の実験から,DNA複製の際に古いH3/H4は新生H3/H4とは混じり合わないことが示されているので86),古いH3/H4の四量体は,一時的に二量体化しても最終的にCAF-1の働きで再び四量体化して,複製後のDNAに結合すると考えられる.一方で,古いH2A/H2BはDNA合成酵素αのヒストン結合活性により複製後のヌクレオソーム構造中に組み込まれるのであろう87).
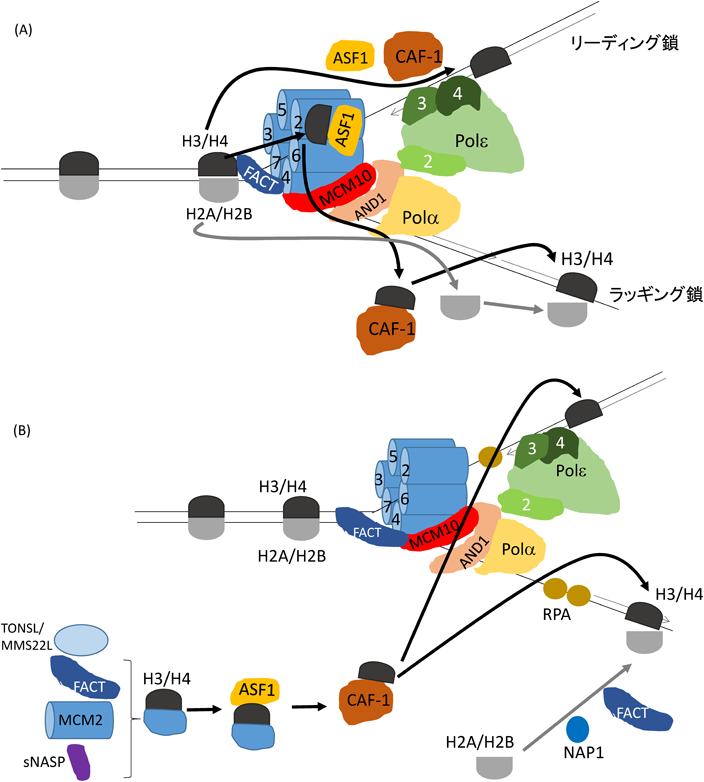
(A)古いヒストンの再結合モデルを示す.複製フォークにおいて,FACT, MCM10とCtf4/AND1はMCM2-7ヘリカーゼに直接的に結合すると思われる.鋳型DNAに結合した古いヒストンH3/H4はFACTとMCM2の働きにより遊離し,MCM2-H3/H4-ASF1なる複合体が形成される.遊離したH3/H4はASF1の働きによりCAF-1に移された後,複製後のラギング鎖DNAに(H3/H4)2として結合する.一方で,DNA合成酵素εのサブユニット3と4はH3/H4の複製後のリーディング鎖への移行に関わる.また,古いH2A/H2BはDNA合成酵素αの働きにより複製後DNAに結合し,ヌクレオソーム構造が再構築される.(B)新生ヒストンの集合モデルを示す.ASF1とCAF-1は,新しいH3/H4の複製後のラギングとリーディング合成鎖への結合に中心的な役割を担う.この反応には,巻き戻されてできた一本鎖DNAに結合するRPAも働く.さらに,MCM2, FACT, TONSL/MMS22LとsNASPは新生H3/H4の集合の初期段階に関わると考えられる.NAP1とFACTは新生H2A/H2Bの複製後DNAへの結合に関わると思われる.
| 古いH2A/H2B | 古いH3/H4 | 新生H2A/H2B | 新生H3/H4 | 古いH3/H4の移行 | 相互作用 | ||
|---|---|---|---|---|---|---|---|
| リーディング鎖 | ラッギング鎖 | ||||||
| MCM2(70-134位) | + | + | ○ | ||||
| FACT | + | + | + | + | MCM4(120-250位) Rtt101(Spt16-Ub) CAF-1, Pol α, RPA | ||
| ASF1 | + | + | RFC | ||||
| CAF-1 | + | + | PCNA | ||||
| TONSL/ MMS22L | + | MCM5, H4K20me0 | |||||
| sNASP | + | ||||||
| Rtt106(酵母) | + | FACT | |||||
| Pol α | + | ○ | |||||
| RPA | + | ||||||
| Ctf4 (AND1) | ○ | Pol α | |||||
| POLE3/POLE4 | + | + | ○ | Pol ε | |||
| NAP1 | + | + | |||||
| DNA複製に関係したヒストンシャペロンに関して,鋳型上の古いヒストンと新生ヒストンのH3/H4とH2A/H2Bとの結合性を示す.+は結合することが示されているものを表す.また,古いH3/H4の複製後のリーディングあるいはラギング合成鎖への移行における各シャペロンの役割を示す.○は移行における役割が明らかにされているものを表す.加えて,各シャペロンと相互作用するタンパク質について示す. | |||||||
最近の研究から,古いヒストンH3/H4は,その約半分がMCM2-Ctf4/AND1-DNA合成酵素αの働きによりラギング鎖側に移行し88, 89),残りの半分がDNA合成酵素εのサブユニット3と4の働きでリーディング鎖側に移行することで90, 91),両鎖に均等に分配されることが示唆されている.MCM2-7ヘリカーゼの活性化に関わるMCM10は,MCM2-7の六量体中で隣り合うサブユニットであるMCM2とMCM6に結合する61).よって,MCM2-MCM6-MCM10-AND1-DNA合成酵素αという軸が,古いヒストンのラギング鎖側への移行に関わると考えられる.DNA合成酵素εサブユニットを含むこれらのタンパク質ネットワークにより,同じ遺伝子発現パターンを持つ細胞の複製が達成されるのであろう.また,複製後DNA上のヒストンは,複製前と同じ位置に結合してヌクレオソーム構造を形成し,その位置の記憶にはMCM2のHBDとDNA合成酵素εのサブユニット3が関与すると報告された92).一方で,細胞が異なる機能を持つ細胞へ分化するとき(非対称分裂)には,特定の遺伝子領域で古いH3/H4はリーディング鎖側とラギング鎖側間に不均一に分配されると考えられる.それに関連して,ショウジョウバエMCM10とDNA合成酵素αのサブユニットが翅の形成に関わることが報告されている93).
新生ヒストンH3/H4の複製したDNAへの結合には,MCM279),ASF194),CAF-1, FACT95),sNASP96),RPA97)そしてTONSL/MMS22L98)などのシャペロンが関わる.それらがどのような個別の役割を担っているのかは今後の課題であるが,全体の経路の後半では,ASF1とCAF-1が働くと考えられる(図6).また,DNAの組換え修復タンパク質であるTONSL/MMS22Lは新生ヒストンH4の20位のリシンの非メチル化体に結合することで,新生DNA鎖を認識して組換え機能を発揮すると考えられ,RPAは複製フォークで巻き戻された一本鎖DNAに結合することで,新生H3/H4のDNAへの結合を促進するのであろう.新生H2A/H2Bの複製後DNAへの結合にはNAP1とFACTが機能するのかもしれない99).
細胞老化はテロメア長の短縮化を含めたさまざまなクロマチン構造の変化により特徴づけられる.MCM2-7タンパク質の存在量は,細胞老化に至る直前から劇的に低下する100–102).その原因の一部は,p53依存的に起こる,MCM2-7 mRNAを不安定化するmicroRNAの発現である103).他のMCM2-7の変化は,老化細胞におけるMCM2とMCM7の細胞質局在である100, 102).MCM2/4/6/7複合体の核局在化はMCM2のN末端部分に存在する核局在化配列に依存する.老化細胞では,MCM2は断片化され,N末端部分を欠いた小タンパク質として存在するので,MCM2の細胞質局在はそのことで説明できるかもしれない102, 104).MCM3に存在する核局在化配列はMCM3/MCM5の核移行に関わるが,MCM3内には核排出配列も存在する.つまり,MCM複合体は細胞周期の進行過程において核と細胞質間を行き来する可能性が考えられる105).実際に,マウス細胞でのMCM3の過剰発現は,核排出配列依存的に細胞増殖を抑制する.出芽酵母の場合,MCM複合体はDNA複製における役割を終えた後に細胞質へ移行することが知られている106).老化細胞においては,MCMの細胞内局在化制御が破綻しているのかもしれない.MCMが細胞質に存在する老化細胞では,4N以上のDNA含有量を持つ細胞が増える102).このことは,G2期で停止した細胞がM期を経ることなしにG1期に入って細胞老化に至るという知見により説明できる107).
クライオ電子顕微鏡による構造解析から,CMG複合体中のMCM2-7とDNAとの相互作用の理解が進みつつある.近い将来,CMGによるDNA巻き戻し機構が明らかになると思われる.古いヒストンH3/H4の複製後DNAへの分配については分子レベルでの解析が進んだ.同定されたヒストンシャペロンが,実際にどのような機構でH3/H4の引き渡しを行い,元と同じ位置でDNA結合をもたらすのかを明らかにする必要がある.新しいヒストンH3/H4の集合についてはさまざまなシャペロンの役割分担を明らかにすることが課題である.MCM2-7は細胞がん化と老化において重要な役割を担う.それぞれの過程で起こる反応の分子レベルの理解もこれからの課題である.
1) Maine, G.T., Subgan, O., & Tye, B.-K. (1984) Mutants of S. cerevisiae defective in the maintenance of minichromosomes. Genetics, 106, 365–385.
2) Gibson, S.I., Surosky, R.T., & Tye, B.K. (1990) The phenotype of the minichromosome maintenance mutant mcm3 is characteristic of mutants defective in DNA replication. Mol. Cell. Biol., 10, 5707–5720.
3) Hennessy, K.M., Lee, A., Chen, E., & Botstein, D. (1991) A group of interacting yeast DNA replication genes. Genes Dev., 5, 958–969.
4) Nasmyth, K. & Nurse, P. (1981) Cell division cycle mutants altered in DNA replication and mitosis in the fission yeast Schizosaccharomyces pombe. Mol. Gen. Genet., 182, 119–124.
5) Coxon, A., Maundrell, K., & Kearsey, S.E. (1992) Fission yeast cdc21+ belongs to a family of proteins involved in an early step of chromosome replication. Nucleic Acids Res., 20, 5571–5577.
6) Miyake, S., Okishio, N., Samejima, I., Hiraoka, Y., Toda, T., Saitoh, I., & Yanagida, M. (1993) Fission yeast genes nda1+ and nda4+, mutations of which lead to S-phase block, chromatin alteration and Ca2+ suppression, are members of the CDC46/MCM2 family. Mol. Biol. Cell, 4, 1003–1015.
7) Takahashi, K., Yamada, H., & Yanagida, M. (1994) Fission yeast minichromosome loss mutants mis cause lethal aneuploidy and replication abnormality. Mol. Biol. Cell, 5, 1145–1158.
8) Yan, H., Gibson, S., & Tye, B.K. (1991) Mcm2 and Mcm3, two proteins important for ARS activity, are related in structure and function. Genes Dev., 5, 944–957.
9) Hu, B., Burkhart, R., Schulte, D., Musahl, C., & Knippers, R. (1993) The P1 family: a new class of nuclear mammalian proteins related to yeast Mcm replication proteins. Nucleic Acids Res., 21, 5289–5293.
10) Kimura, H., Nozaki, N., & Sugimoto, K. (1994) DNA polymerase alpha associated protein P1, a murine homolog of yeast MCM3, changes its intranuclear distribution during the DNA synthetic period. EMBO J., 13, 4311–4320.
11) Todorov, I.T., Pepperkok, R., Philipova, R.N., Kearsey, S.E., Ansorge, W., & Werner, D. (1994) A human nuclear protein with sequence homology to a family of early S phase proteins is required for entry into S phase and for cell division. J. Cell Sci., 107, 253–265.
12) Kubota, Y., Mimura, S., Nishimoto, S.-I., Takisawa, H., & Nojima, H. (1995) Identification of the yeast MCM3 related protein as a component of Xenopus DNA replication licensing factor. Cell, 81, 601–609.
13) Chong, J.P.J., Mahbubani, H.M., Khoo, C.-Y., & Blow, J.J. (1995) Purification of an MCM-containing complex as a component of the DNA replication licensing system. Nature, 375, 418–421.
14) Madine, M.A., Khoo, C.-Y., Mills, A.D., & Laskey, R.A. (1995) MCM3 complex required for cell cycle regulation of DNA replication in vertebrate cells. Nature, 375, 421–424.
15) Koonin, E.V. (1993) A common set of conserved motifs in a vast variety of putative nucleic acid-dependent ATPases including MCM proteins involved in the initiation of eukaryotic DNA replication. Nucleic Acids Res., 21, 2541–2547.
16) Adachi, Y., Usukura, J., & Yanagida, M. (1997) A globular complex formation by Nda1 and the other five members of the MCM protein family in fission yeast. Genes Cells, 2, 467–479.
17) Ishimi, Y., Ichinose, S., Omori, A., Sato, K., & Kimura, H. (1996) Binding of human minichromosome maintenance proteins with histone H3. J. Biol. Chem., 271, 24115–24122.
18) Ishimi, Y. (1997) A DNA helicase activity is associated with an MCM4, -6, and -7 protein complex. J. Biol. Chem., 39, 24508–24513.
19) Ishimi, Y., Komamura, Y., You, Z., & Kimura, H. (1998) Biochemical function of mouse minichromosome maintenance 2 protein. J. Biol. Chem., 273, 8369–8375.
20) Sato, M., Gotow, T., You, Z., Komamura-Kohno, Y., Uchiyama, Y., Yabuta, N., Nojima, H., & Ishimi, Y. (2000) Electron microscopic observation and single-stranded DNA binding activity of the Mcm4,6,7 complex. J. Mol. Biol., 300, 421–431.
21) Schwacha, A. & Bell, S.P. (2001) Interactions between two catalytically distinct MCM subgroups are essential for coordinated ATP hydrolysis and DNA replication. Mol. Cell, 8, 1093–1104.
22) Bochman, M.L., Bell, S.P., & Schwacha, A. (2008) Subunit organization of Mcm2-7 and the unequal role of active sites in ATP hydrolysis and viability. Mol. Cell. Biol., 28, 5865–5873.
23) Prokhorova, T.A. & Blow, J.J. (2000) Sequential MCM/P1 subcomplex assembly is required to form a heterohexamer with replication lisencing activity. J. Biol. Chem., 275, 2491–2498.
24) Labib, K., Tercero, J.A., & Diffley, J.F.X. (2000) Uninterrupted MCM2-7 function required for DNA replication fork progression. Science, 288, 1643–1646.
25) Yu, Z., Fend, D., & Liang, C. (2004) Pairwise interactions of the six human MCM protein subunits. J. Mol. Biol., 340, 1197–1206.
26) Coue, M., Amariglio, F., Maiorano, D., Bocquet, S., & Mechali, M. (1998) Evidence for different MCM subcomplexes with different binding to chromatin in Xenopus. Exp. Cell Res., 245, 282–289.
27) Sherman, D.A., Pasion, S.G., & Forsburg, S.L. (1998) Multiple domains of fission yeast Cdc19p (MCM2) are required for its association with the core MCM complex. Mol. Biol. Cell, 9, 1833–1845.
28) Pasion, S.G. & Forsburg, S.L. (1999) Nuclear localization of Schizosaccharomyces pombe Mcm2/Cdc19p requires MCM complex assembly. Mol. Biol. Cell, 10, 4043–4057.
29) Forsburg, S.L. (2004) Eukaryotic MCM proteins: beyond replication initiation. Microbiol. Mol. Biol. Rev., 68, 109–131.
30) Schultz, R.W., Lee, T.-J., Allen, G.C., Thompson, W.F., & Hanley-Bowdoin, L. (2009) Dynamic localization of the DNA replication proteins MCM5 and MCM7 in plants. Plant Physiol., 150, 658–669.
31) Ishimi, Y., Sagara, C., & Koseki, Y. (2019) Effects of forcedly expressed MCM2-7 proteins on nuclear structure in HeLa cells. Curr. Top. Biochem. Res., 20, 17–31.
32) Kanter, D.M., Bruck, I., & Kaplan, D.L. (2008) Mcm subunits can assemble into two different active unwinding complexes. J. Biol. Chem., 283, 31172–31182.
33) Bochman, M.L. & Schwacha, A. (2008) The Mcm2-7 complex has in vitro helicase activity. Mol. Cell, 31, 287–293.
34) Takayama, Y., Kamimura, Y., Okawa, M., Muramatsu, M., Sugino, A., & Araki, H. (2003) GINS, a novel multiprotein complex required for chromosomal DNA replication in budding yeast. Genes Dev., 17, 1153–1165.
35) Gambus, A., Jones, R.C., Sanchez-Diaz, A., Kanemaki, M., van Deursen, F., Edmondson, R.D., & Labib, K. (2006) GINS maintains association of Cdc45 with MCM in replisome progression complexes at eukaryotic DNA replication forks. Nat. Cell Biol., 8, 358–366.
36) Moyer, S.E., Lewis, P.W., & Botchan, M.R. (2006) Isolation of the Cdc45/Mcm2-7/GINS (CMG) complex, a candidate for the eukaryotic DNA replication fork helicase. Proc. Natl. Acad. Sci. USA, 103, 10236–10241.
37) Ilves, I., Petojevic, T., Pesavento, J.J., & Botchan, M.R. (2010) Activation of the MCM2-7 helicase by association with Cdc45 and GINS proteins. Mol. Cell, 37, 247–258.
38) Kang, Y.-H., Galal, W.C., Farina, A., Tappin, I., & Hurwitz, J. (2012) Properties of the human Cdc45/Mcm2-7/GINS helicase complex and its action with DNA polymerase ε in rolling circle DNA synthesis. Proc. Natl. Acad. Sci. USA, 109, 6042–6047.
39) Costa, A., Renault, L., Swuec, P., Petojevic, T., Pesavento, J.J., Ilves, I., MacLellan-Gibson, K., Fleck, R.A., Botchan, M.R., & Berger, J.M. (2014) DNA binding polarity, dimerization, and ATPase ring remodeling in the CMG helicase of the eukaryotic replisome. eLife, 3, e03273.
40) Meagher, M., Epling, L.B., & Enemark, E.J. (2019) DNA translocation mechanism of the MCM complex and implications for replication initiation. Nat. Commun., 10, 3117.
41) Li, N., Zhai, Y., Zhang, Y., Li, W., Yang, M., Lei, J., Tye, B.-K., & Gao, N. (2015) Structure of the eukaryotic MCM complex at 3.8 Å. Nature, 524, 186–191.
42) Brewster, A.S., Wang, G., Yu, X., Greenleaf, W.B., Carazo, J.M., Tjajadi, M., Klein, M.G., & Chen, X.S. (2008) Crystal structure of a near-full-length archaeal MCM: functional insights for an AAA+ hexameric helicase. Proc. Natl. Acad. Sci. USA, 105, 20191–20196.
43) Barry, E., Lovett, J.E., Costa, A., Lea, S.M., & Bell, S.D. (2009) Intersubunit allosteric communication mediated by a conserved loop in the MCM helicase. Proc. Natl. Acad. Sci. USA, 106, 1051–1056.
44) You, Z., Ishimi, Y., Masai, H., & Hanaoka, F. (2002) Roles of Mcm7 and Mcm4 subunits in the DNA helicase activity of the mouse Mcm4/6/7 complex. J. Biol. Chem., 277, 42471–42479.
45) Poplawski, A., Grabowski, B., Long, S.E., & Kelman, Z. (2001) Domain of the archaeal minichromosome maintenance protein is required for helicase activity. J. Biol. Chem., 276, 49371–49377.
46) Fletcher, R.J., Shen, J., Gómez-Llorente, Y., Martin, C.S., Carazo, J.M., & Chen, X.S. (2005) Double hexamer disruption and biochemical activities of Methanobacterium thermoautotrophicum MCM. J. Biol. Chem., 280, 42405–42410.
47) Evrin, C., Fernández-Cid, A., Riera, A., Zech, J., Clarke, P., Herrera, M.C., Tognetti, S., Lurz, R., & Speck, C. (2013) The ORC/Cdc6/MCM2-7 complex facilitates MCM2-7 dimerization during prereplicative complex formation. Nucleic Acids Res., 42, 2257–2269.
48) Ali, F.A., Renault, L., Gannon, J., Gahlon, H.L., Kotecha, A., Zhou, J.C., Rueda, D., & Costa, A. (2016) Cryo-EM structures of the eukaryotic replicative helicase bound to a translocation substrate. Nat. Commun., 7, 10708.
49) Georgescu, R., Yuan, Z., Bai, L., de Luna Almeida Santos, R., Sun, J., Zhang, D., Yurieva, O., Li, H., & O’Donnell, M.E. (2017) Structure of eukaryotic CMG helicase at a replication fork and implications to replisome architecture and origin initiation. Proc. Natl. Acad. Sci. USA, 114, E697–E706.
50) Yuan, Z., Riera, A., Bai, L., Sun, J., Nandi, S., Spanos, C., Chen, Z.A., Barbon, M., Rappsilber, J., Stillman, B., et al. (2017) Structural basis of MCM2-7 replicative helicase loading by ORC-Cdc6 and Cdt1. Nat. Struct. Mol. Biol., 24, 316–324.
51) Noguchi, Y., Yuan, Z., Bai, L., Schneider, S., Zhao, G., Stillman, B., Speck, C., & Li, H. (2017) Cryo-EM structure of Mcm2-7 double hexamer on DNA suggests a lagging-strand extrusion model. Proc. Natl. Acad. Sci. USA, 114, E9529–E9538.
52) Froelich, C.A., Kang, S., Epling, L.B., Bell, S.P., & Enemark, E.J. (2014) A conserved MCM single-stranded DNA binding element is essential for replication initiation. eLife, 3, e01993.
53) Eickhoff, P., Kose, H.B., Martino, F., Petojevic, T., Ali, F.A., Locke, J., Tamberg, N., Nans, A., Berger, J.M., Botchan, M.R., et al. (2019) Molecular basis for ATP-hydrolysis-driven DNA translocation by the CMG helicase of the eukaryotic replisome. Cell Rep., 28, 2673–2688.
54) Kose, H.B., Larsen, N.B., Duxin, J.P., & Yardimci, H. (2019) Dynamics of the eukaryotic replicative helicase at lagging-strand protein barriers support the steric exclusion model. Cell Rep., 26, 2113–2125.
55) Burnham, D.R., Kose, H.B., Hoyle, R.B., & Yardimci, H. (2019) The mechanism of DNA unwinding by the eukaryotic replicative helicase. Nat. Commun., 10, 2159.
56) Tan, B.C.-M., Chien, C.-T., Hirose, S., & Lee, S.-C. (2006) Functional cooperation between FACT and MCM helicase facilitates initiation of chromatin DNA replication. EMBO J., 25, 3975–3985.
57) Campos, E.I., Smits, A.H., Kang, Y.-H., Landry, S., Escobar, T.M., Nayak, S., Ueberheide, B.M., Durocher, D., Vermeulen, M., Hurwitz, J., et al. (2015) Analysis of the histone H3.1 interactome: a suitable chaperone for the right event. Mol. Cell, 60, 697–709.
58) Shi, Y.K., Yu, Y.P., Zhu, Z.H., Han, Y.C., Ren, B., Nelson, J.B., & Luo, J.H. (2008) MCM7 interacts with androgen receptor. Am. J. Pathol., 173, 1758–1767.
59) Lossaint, G., Larroqu, M., Ribeyre, C., Bec, N., Larroque, C., Décaillet, C., Gari, K., & Constantinou, A. (2013) FANCD2 binds MCM proteins and controls replisome function upon activation of S phase checkpoint signaling. Mol. Cell, 51, 678–690.
60) Cucco, F., Palumbo, E., Camerini, S., D’Alessio, B., Quarantotti, V., Casella, M.L., Rizzo, I.M., Cukrov, D., Della, D., Russo, A., et al. (2018) Separase prevents genomic instability by controlling replication fork speed. Nucleic Acids Res., 46, 267–278.
61) Mayle, R., Langston, L., Molloy, K.R., Zhang, D., Chait, B.T., & O’Donnell, M.E. (2019) Mcm10 has potent strand-annealing activity and limits translocase-mediated fork regression. Proc. Natl. Acad. Sci. USA, 116, 798–803.
62) Numata, Y., Ishihara, S., Hasegawa, N., Nozaki, N., & Ishimi, Y. (2010) Interaction of human MCM2-7 proteins with TIM, TIPIN and Rb. J. Biochem., 147, 917–927.
63) DeFonseca, C.J., Shu, F., & Zhang, J.J. (2001) Identification of two residues in MCM5 critical for the assembly of MCM complexes and Stat1-mediated transcription activation in response to IFN-γ. Proc. Natl. Acad. Sci. USA, 98, 3034–3039.
64) Holland, L., Downey, M., Song, X., Gauthier, L., Bell-Rogers, P., & Yankulov, K. (2002) Distinct parts of minichromosome maintenance protein 2 associate with histone H3/H4 and RNA polymerase II holoenzyme. Eur. J. Biochem., 269, 5192–5202.
65) Wang, X. & Ishimi, Y. (2018) Function of the amino-terminal region of human MCM4 in helicase activity. J. Biochem., 164, 449–460.
66) Champasa, K., Blank, C., Friedman, L.J., Gelles, J., & Bell, S.P. (2019) A conserved Mcm4 motif is required for Mcm2-7 double-hexamer formation and origin DNA unwinding. eLife, 8, e45538.
67) Ishimi, Y. (2018) Regulation of MCM2-7 function. Genes Genet. Syst., 93, 125–133.
68) Hendrickson, M., Madine, M., Dalton, S., & Gautier, J. (1996) Phosphorylation of MCM4 by cdc2 protein kinase inhibits the activity of the minichromosome maintenance complex. Proc. Natl. Acad. Sci. USA, 93, 12223–12228.
69) Findeisen, M., El Denary, M., Kapitza, T., Graf, R., & Strausfeld, U. (1999) Cyclin-dependent kinase activity affects chromatin binding of ORC and Cdc6 and MCM in egg extracts of Xenopus laevis. Eur. J. Biochem., 264, 415–426.
70) Ishimi, Y., Komamura-Kohno, Y., You, Z., Omori, A., & Kitagawa, M. (2000) Inhibition of Mcm4,6,7 helicase activity by phosphorylation with cyclinA/Cdk2. J. Biol. Chem., 275, 16235–16241.
71) Ishimi, Y. & Komamura-Kohno, Y. (2001) Phosphorylation of Mcm4 at specific sites by cyclin-dependent kinase leads to loss of Mcm4,6,7 helicase activity. J. Biol. Chem., 276, 34428–34433.
72) Blow, J.J. & Dutta, A. (2005) Preventing re-replication of chromosomal DNA. Nat. Rev. Mol. Cell Biol., 6, 476–486.
73) Masai, H., Taniyama, C., Ogino, K., Matsui, E., Kakusho, N., Matsumoto, S., Kim, J.M., Ishii, A., Tanaka, T., Kobayashi, T., et al. (2006) Phosphorylation of MCM4 by Cdc7 kinase facilitates its interaction with Cdc45 on the chromatin. J. Biol. Chem., 281, 39249–39261.
74) Masai, H., Matsui, E., You, Z., Ishimi, Y., Tamai, K., & Arai, K. (2000) Human Cdc7-related kinase complex. In vitro phosphorylation of MCM by concerted actions of Cdks and Cdc7 and that of a critical threonine residue of Cdc7 by Cdks. J. Biol. Chem., 275, 29042–29052.
75) Bell, S.P. & Labib, K. (2016) Chromosome duplication in Saccharomyces cerevisiae. Genetics, 203, 1027–1067.
76) Tanaka, H., Katou, Y., Yagura, M., Saitoh, K., Itoh, T., Araki, H., Bando, M., & Shirahige, K. (2009) Ctf4 coordinates the progression of helicase and DNA polymerase α. Genes Cells, 14, 807–820.
77) Villa, F., Simon, A.C., Bazan, M.A.O., Kikenny, M.L., Wirthensohn, D., & Wightman, M. (2016) Ctf4 is a hub in the eukaryotic replisome that links multiple CIP-box proteins to the CMG helicase. Mol. Cell, 63, 385–396.
78) Ishimi, Y., Komamura, Y., You, Z., & Kimura, H. (1998) Biochemical function of mouse minichromosome maintenance 2 protein. J. Biol. Chem., 273, 8369–8375.
79) Ishimi, Y., Komamura-Kohno, Y., Arai, K., & Masai, H. (2001) Biochemical activities associated with mouse Mcm2 protein. J. Biol. Chem., 276, 42744–42752.
80) Groth, A., Corpet, A., Cook, A.J.L., Roche, D., Bartek, J., Lukas, J., & Almouzni, G. (2007) Regulation of replication fork progression through histone supply and demand. Science, 318, 1928–1931.
81) Foltman, M., Evrin, C., De Piccoli, G., Jones, R.C., Edmondson, R.D., Katou, Y., Nakato, R., Shirahige, K., & Labib, K. (2013) Eukaryotic replisome components cooperate to process histones during chromosome replication. Cell Rep., 3, 892–904.
82) Huang, H., Strømme, C.B., Saredi, G., Hödl, M., Strandsby, A., González-Aguilera, C., Chen, S., Groth, A., & Patel, D.J. (2015) A unique binding mode enables MCM2 to chaperon histones H3-H4 at replication forks. Nat. Struct. Mol. Biol., 22, 618–626.
83) Liu, W.H., Roemer, S.C., Zhou, Y., Shen, Z.-J., Dennehey, B.K., Balabaugh, J.L., Liddle, J.C., Nemkov, T., Ahn, N.G., Hansen, K.C., et al. (2016) The Cac1 subunit of histone chaperon CAF-1 organizes CAF-1-H3/H4 architecture and tetramerizes histones. eLife, 5, e18023.
84) Wang, H., Wang, M., Yang, N., & Xu, R.-M. (2015) Structure of the quaternary complex of histone H3-H4 heterodimer with chaperon ASF1 and the replicative helicase subunit MCM2. Protein Cell, 6, 693–697.
85) Richet, N., Liu, D., Legrand, P., Velours, C., Corpet, A., Gaubert, A., Bakall, M., Moal-Raisin, G., Guerois, R., Compper, C., et al. (2015) Structural insight into how the human helicase subunit MCM2 may act as a histone chaperon together with ASF1 at the replication fork. Nucleic Acids Res., 43, 1905–1917.
86) Xu, M., Long, C., Chen, X., Huang, C., Chen, S., & Zhu, B. (2010) Partitioning of histone H3-H4 tetramers during DNA replication-dependent chromatin assembly. Science, 328, 94–98.
87) Evrin, C., Maman, J.D., Diamante, A., Pellegrini, L., & Labib, K. (2018) Histone H2A-H2B binding by polα in the eukaryotic replisome contributes to the maintenance of repressive chromatin. EMBO J., 37, e99021.
88) Petryk, N., Dalby, M., Wenger, A., Stromme, C.B., Strandsby, A., Andersson, R., & Groth, A. (2018) MCM2 promotes symmetric inheritance of modified histones during DNA replication. Science, 361, 1389–1392.
89) Gan, H., Serra-Cardona, A., Hua, X., Zhou, H., Labib, K., Yu, C., & Zhang, Z. (2018) The Mcm2-Ctf4-Polα axis facilitates parental histone H3-H4 transfer to lagging strands. Mol. Cell, 72, 140–151.
90) Bellelli, R., Belan, O., Pye, V.E., Clement, C., Maslen, S.L., Skehel, J.M., Cherepanov, P., Almouzni, G., & Boulton, S.J. (2018) POLE3-POLE4 is a histone H3-H4 chaperon that maintains chromatin integrity during DNA replication. Mol. Cell, 72, 112–126.
91) Yu, C., Gan, H., Serra-Cardona, A., Zhang, L., Gan, S., Sharma, S., Johansson, E., Chabes, A., Xu, R.-M., & Zhang, Z. (2018) A mechanism for preventing asymmetric histone segregation onto replicating DNA strands. Science, 361, 1386–1389.
92) Schlissel, G. & Rine, J. (2019) The nucleosome core particle remembers its position through DNA replication and RNA transcription. Proc. Natl. Acad. Sci. USA, 116, 20605–20611.
93) Kohzaki, H., Asano, M., & Murakami, Y. (2018) DNA replication machinery is required for development in Drosophila. Front. Biosci., 23, 493–505.
94) Jasencakova, Z., Scharf, A.N.D., Ask, K., Corpet, A., Imhof, A., Almouzni, G., & Groth, A. (2010) Replication stress interferes with histone recycling and predeposition marking of new histones. Mol. Cell, 37, 736–743.
95) Yang, J., Zhang, X., Feng, J., Leng, H., Li, S., Xiao, J., Liu, S., Xu, Z., Xu, J., Li, D., et al. (2016) The histone chaperon FACT contributes to DNA replication-coupled nucleosome assembly. Cell Rep., 14, 1128–1141.
96) Campos, E.I., Fillingham, J., Li, G., Zheng, H., Voigt, P., Kuo, W.-H.W., Seepany, H., Gao, Z., Day, L.A., Greenblatt, J.F., et al. (2010) The program for processing newly-synthesized histones H3.1 and H4. Nat. Struct. Mol. Biol., 17, 1343–1351.
97) Lin, S., Xu, Z., Leng, H., Zheng, P., Yang, J., Chen, K., Feng, J., & Li, Q. (2017) RPA binds histone H3-H4 and functions in DNA replication-coupled nucleosome assembly. Science, 355, 415–420.
98) Saredi, G., Huang, H., Hammond, C.M., Alabert, C., Bekker-Jensen, S., Forne, I., Reverón-Gómez, N., Foster, B.M., Mlejnkova, L., Bartke, T., et al. (2016) H4 K20me0 marks post-replicative chromatin and recruits the TONSL-MMS22L DNA repair complex. Nature, 534, 714–718.
99) Hammond, C.M., Strømme, C.B., Huang, H., Patel, D.J., & Groth, A. (2017) Histone chaperon networks shaping chromatin function. Nat. Rev. Mol. Cell Biol., 18, 141–158.
100) Dumit, V.I., Küttner, V., Käppler, J., Piera-Velazquez, S., Jimenez, S.A., Bruckner-Tuderman, L., Uitto, J., & Dengjel, J. (2014) Altered MCM protein levels and autophagic flux in aged and systemic sclerosis dermal fibroblasts. J. Invest. Dermatol., 134, 2321–2330.
101) Flach, J., Bakker, S.T., Mohrin, M., Conroy, P.C., Pietras, E.M., Reynaud, D., Alvarez, S., Diolaiti, M.E., Ugarte, F., Forsberg, E.C., et al. (2014) Replication stress is a potent driver of functional decline in aging haematopoietic stem cells. Nature, 512, 198–202.
102) Suzuki, Y., Yamaguchi, Y., Hanada, H., & Ishimi, Y. (2019) Changes in MCM2-7 proteins at senescence. Genes Genet. Syst., 94, 123–132.
103) Bai, G., Smolka, M.B., & Schimenti, J.C. (2016) Chronic DNA replication stress reduces replicative lifespan of cells by TRP53-dependent, microRNA-assisted MCM2-7 downregulation. PLoS Genet., 12, 1005787.
104) Harada, H., Nakagawa, H., Takaoka, M., Lee, J., Herlyn, M., Diehl, J.A., & Rustgi, A.K. (2008) Cleavage of MCM2 licensing protein fosters senescence in human keratinocyte. Cell Cycle, 7, 3534–3538.
105) Chuang, C.-H., Yang, D., Bai, G., Freeland, A., Pruitt, S.C., & Schimenti, J.C. (2012) Post-transcriptional homeostasis and regulation of MCM2-7 in mammalian cells. Nucleic Acids Res., 40, 4914–4924.
106) Labib, K., Diffley, J.F.X., & Kearsey, S.E. (1999) G1-phase and B-type cyclins exclude the DNA-replication factor Mcm4 from the nucleus. Nat. Cell Biol., 1, 415–422.
107) Johmura, Y., Shimada, M., Misaki, T., Naiki-Ito, A., Miyoshi, H., Motoyama, N., Ohtani, N., Hara, E., Nakamura, M., Morita, A., et al. (2014) Necessary and sufficient role for a mitosis skip in senescence induction. Mol. Cell, 55, 73–84.

薬学博士.
1975年静岡大学理学部卒業,79年金沢大学大学院薬学研究科修士課程修了,82年東京大学大学院薬学研究科博士課程修了,83~88年東京大学薬学部助手,88~2004年三菱化学生命科学研究所研究員,04~19年茨城大学理学部教授.
研究テーマと抱負1960年頃から現在までの生命科学の発展に寄与した主要な研究成果について,その原著論文を読むことで勉強しています.将来,それらについてまとめたものを発表したいと考えています.
趣味低山歩き,ゴルフ.

This page was created on 2020-03-04T11:06:11.79+09:00
This page was last modified on 2020-04-03T11:12:47.000+09:00
このサイトは(株)国際文献社によって運用されています。