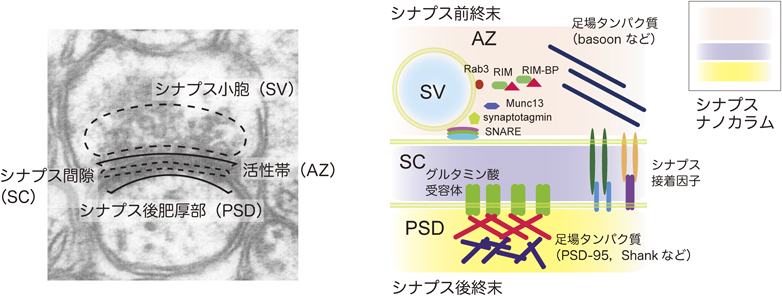
哺乳類の脳は1000億を超える神経細胞から構成されているといわれている.神経細胞は,信号伝達を担う細胞接着構造であるシナプスを介して互いに接続して回路を形成し,脳機能に必要な情報処理を行う.シナプスの多くは,軸索末端と樹状突起との間に形成される.シナプスの軸索末端側をシナプス前終末,樹状突起側をシナプス後終末と呼ぶ.シナプス前終末では,電位依存性Ca2+チャネルの活性化によるCa2+の流入が,神経伝達物質を内包するシナプス小胞(synaptic vesicle:SV)と細胞膜との融合を促進して,神経伝達物質が放出される.放出された神経伝達物質は,シナプス間隙の中を拡散した後,シナプス後終末に存在するグルタミン酸受容体などの神経伝達物質依存的なイオンチャネルに結合して活性化する.その結果,イオン透過によって電気ポテンシャルが変化し,化学信号は再び電気信号に変換される.シナプスは,放出される神経伝達物質の種類の違いにより,興奮性と抑制性の2種類に大別される.そのバランス調節は神経回路の正常な動作に不可欠である.
シナプス前終末の構造は,蓄積したSVと神経伝達物質を放出するために必要な分子群(SNAREやsynaptotagmin, Munc13等の膜融合関連因子,電位依存性Ca2+チャネル,BassoonやPiccoloなどの巨大な足場タンパク質等)が集合した活性帯(active zone:AZ)に特徴づけられる(図1).一方,シナプス後終末の構造は,グルタミン酸受容体などのさまざまな神経伝達物質受容体とPSD-95やShank2などの足場タンパク質が集合したシナプス後肥厚部(postsynaptic density:PSD)に特徴づけられる.近年の超解像顕微鏡による解析では,SV, AZ, PSDが整列して配置された「ナノカラム」と呼ばれる機能ユニットを形成していることが示されており,正常な信号伝達に必要であると考えられている1, 2).シナプス間隙を介して整列するには,シナプス前終末と後終末が物理的に橋渡しされる必要性があるが,その役割をシナプス接着分子が担うと考えられる.シナプス接着分子には,免疫グロブリン(immunoglobulin:Ig)スーパーファミリーやカドヘリンファミリー,ロイシンリッチリピート(leucine-rich repeat:LRR)タンパク質ファミリー等に属する多様な分子が含まれるが,本稿では,主に最近になって立体構造が明らかになったシナプス接着分子について概説する.前半は,発生過程でシナプスの分化誘導を行うシナプスオーガナイザーと呼ばれる接着分子群3–5)について,後半は,てんかん関連リガンド-受容体複合体LGI1-ADAM226)について概説する.LGI1-ADAM22は,厳密には接着分子とは異なるが,シナプス間隙をまたいで信号伝達の調節を担う重要な分子として紹介したい.
シナプスオーガナイザーは,細胞外ドメイン(extracellular domain:ECD)を介してシナプス間隙をまたいだ,いわゆる「トランスシナプティック」な相互作用を生じることで,シナプス形成を誘導する.シナプス標的(神経細胞の接続先)の選択や興奮性と抑制性のバランス調節で重要な役割を担う.バランス調節の破綻は自閉症や知的障害などの神経発達障害に関係していると考えられており,シナプスオーガナイザーをコードする遺伝子の異常と神経発達障害との関連を示唆する論文が数多く報告されている.
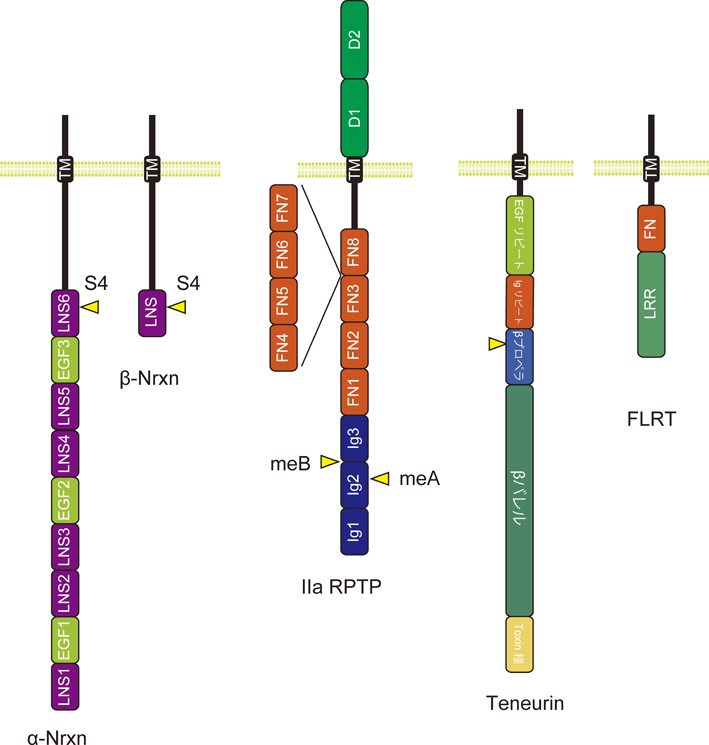
シナプス前終末の主なシナプスオーガナイザーとして,Neurexin(Nrxn)5)やIIa型受容体チロシンホスファターゼ(receptor protein tyrosine phosphatase:RPTP)3, 4),Teneurin(Ten)7, 8)の3種類のタンパク質ファミリーが知られている(図2).
Nrxnは,内部プロモーターの存在によってαとβの2種類のアイソフォーム(α-Nrxnおよびβ-Nrxn)が発現する.α-NrxnのECDは,上皮成長因子(epidermal growth factor:EGF)様のドメインが二つのlaminin/Nrxn/sex-hormone-binding globular(LNS)ドメインに挟まれたLNS-EGF-LNSが3回繰り返されたドメイン構造を持つ.一方,β-NrxnのECDは一つのLNSのみで構成される.β-NrxnのLNSは,α-Nrxnの6番目のLNSに対応する.この6番目のLNSが,シナプス後終末のシナプスオーガナイザーと相互作用する.Nrxnと相互作用するシナプス後終末のシナプスオーガナイザーとして,イオンチャネル型グルタミン酸受容体ファミリーのGluD2, LRR膜貫通タンパク質(LRR transmembrane proteins:LRRTM)とNeuroligin(Nlgn)が知られている9–11).GluD2は,分泌タンパク質Cbln1を介してNrxnと相互作用する.また,ごく最近になって,GluD1がCbln4を介してNrxnと相互作用し,抑制性のシナプス形成を誘導することが報告されており12),GluD2-Cbln1-Nrxnと同様の複合体を形成することが予想される.
IIa型RPTPは,脊椎動物ではPTPδ, PTPσ, LARの3種類がある.一方,無脊椎動物はLAR 1種類のみを持つ.IIa型RPTPのECDは,三つのIgドメイン(Ig1~3)と四つもしくは八つのIII型フィブロネクチン(fibronectin:FN)ドメイン(FN1~8)がN末端側から順に並んだ形で構成される(図2).IIa型RPTPは,シナプス後終末のシナプスオーガナイザーであるインターロイキン1受容体アクセサリータンパク質(interleukin-1 receptor accessory protein:IL-1RAcP)やIL-1RAcP様1(IL-1RAcP-like 1:IL1RAPL1),TrkC, SlitおよびTrk様(Slit- and Trk-like:Slitrk)タンパク質ファミリー,synaptic adhesion-like molecule 3および5(SALM3およびSALM5),ネトリンGリガンド-3(Netrin G ligand 3:NGL-3)と相互作用してシナプス形成を誘導する13–19).NGL-3を除いたこれらのシナプスオーガナイザーは,IIa型RPTPのIgドメインと相互作用する.NGL-3は,FNドメインと相互作用すると考えられている.
Tenは,進化の過程で細菌の毒素と真核生物の膜受容体が融合して生じたと考えられているユニークな受容体である20, 21).2000残基を超える巨大なTenのECDは,EGFリピート,Igリピート,βプロペラ,βバレル,Toxin様の五つのドメインで構成される(図2).Tenと相互作用するシナプス後終末のシナプスオーガナイザーとして同定されているのは,接着Gタンパク質共役型受容体であるlatrophilin(Lphn)のみである22–24).TenとLphnによるシナプス形成の誘導には,シナプス前終末に局在する共受容体であるfibronectin leucine-rich repeat transmembrane protein(FLRT)も必要である25, 26).FLRTのECDは,LRRドメインとFNドメインの二つのドメインで構成され(図2),LRRドメインがLphnと相互作用する.
3. 選択的スプライシングによるシナプスオーガナイザー間の相互作用の調節
シナプスオーガナイザーの多くは,選択的スプライシングによって生じるNrxnやIIa型RPTPのバリアントのそれぞれに対して異なる結合能を示す3–5).たとえば,シナプス後終末のシナプスオーガナイザーとの結合能を制御するNrxnのスプライス部位は,α-Nrxnとβ-Nrxnに共通のLNSドメインに存在する4番目のスプライス部位(S4;図2)であり,Cbln1-GluD2はS4に挿入のあるバリアント(+S4)と選択的に結合する11).一方,LRRTMは,S4に挿入のないバリアント(−S4)と選択的に結合する27, 28).同様なIIa型RPTPのスプライス部位は,Ig2内およびIg2とIg3の境界に存在する(図2).この2か所に短いエクソン(ミニエクソン)に対応するペプチド[ミニエクソンペプチドA(mini-exon peptide A:meA)および,ミニエクソンペプチドB(mini-exon peptide B:meB)]が挿入される.最もバリエーションが豊かなPTPδの場合は,9残基,6残基,3残基のmeAが挿入されたバリアント(A9, A6, A3)とmeAが欠失したバリアント(A−)が存在する.meBについては,挿入されたバリアント(B+)と欠失したバリアント(B−)の二つのバリアントが存在する.これらの組合わせで合計8種類のバリアントが発現していることになる.IL1RAPL1は,A9/B+やA6/B+, A9/B−のバリアントに結合するのに対して,IL-1RAcPは,A9/B+, A6/B+, A3/B+のバリアントに強く結合する14, 16, 29).また,Slitrkは,meAとは無関係に,B+のバリアントのみに選択的に結合する30, 31).これらの結合能は,実際のシナプス誘導活性と正の相関を示す.SALM5も,B+のバリアントに対してより強い結合を示す32, 33)が,逆にシナプス誘導活性を抑制するという報告もある34).一方,SALM3では,meBがシナプス誘導活性に必要であることが報告されている35).Tenにもスプライスバリアントが存在し,ホモ二量体化とヘテロ二量体化の制御に関わっていると提案されている22, 23, 36).しかし,バリアント選択的な結合相手は同定されておらず,また,スプライス部位は,Lphnとの相互作用部位と空間的に離れて位置しており37, 38),議論の余地が残されている.
1)Nrxn
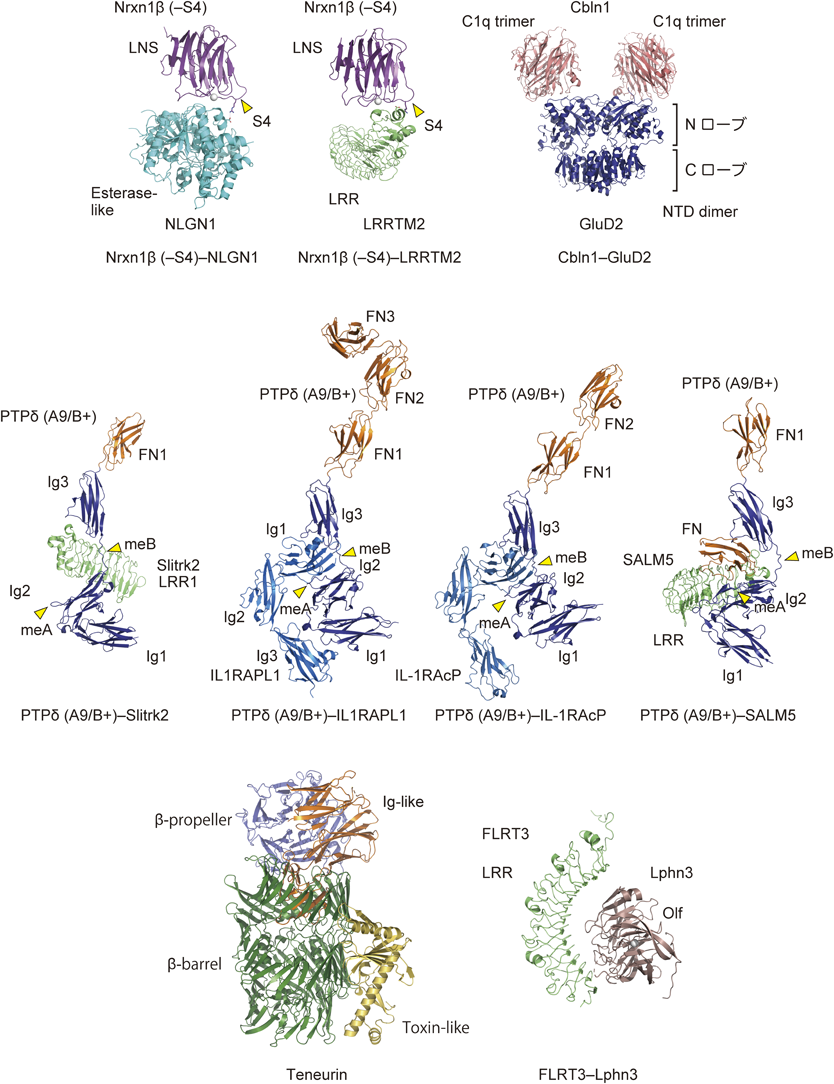
NrxnとNlgnとの相互作用様式は,主にβ-NrxnのLNSドメインとNlgnのコリンエステラーゼ様ドメインの複合体の立体構造から明らかにされている(図3;上段左)39–41).複数の研究グループから,異なるアイソフォームやスプライスバリアントの立体構造が報告され,さまざまな組合わせでの親和性も測定されている42).明確なON/OFFの制御ではない(基本的にどの組合わせでもシナプス誘導は生じる)ので,親和性の差をどのように捉えるかによって解釈は異なるが,興奮性と抑制性のシナプスのバランス調節への寄与などが考えられている.
Nrxn-Cbln1-GluD2複合体については,英国MRCのAricescuと慶應大の柚崎の共同グループが,Cbln1とGluD2のN末端ドメイン(NTD)の複合体を模した融合体の結晶構造を決定して,Cbln1とGluD2との相互作用様式を明らかにしている(図3;上段右)43).Cbln1は,補体の構成要素C1qに代表されるタンパク質ファミリー(C1qファミリー)に属しており,C1qファミリーに特徴的な3回対称軸を持つ球状の三量体を形成する.イオンチャネル型グルタミン酸受容体のN末端ドメインの立体構造は二枚貝の貝殻(clamshell)にたとえられ,NローブとCローブの二つの部位に分けられる.Cbln1とGluD2の融合体構造では,二量体化したGluD2 NTDの各プロトマーのNローブに,三量体化したCbln1が相互作用して,2 : 6のヘテロ八量体を形成している.+S4依存的な相互作用の詳細は依然として不明ではあるが,Nrxn1β(+S4)LNS-Cbln1複合体の電子顕微鏡負染色像は,Cbln1の球状のドメインよりもさらにN末端側の領域が,Nrxn1βのS4領域を認識することを示唆している.上述のGluD1-Cbln4-Nrxnの複合体は,Nrxn-Cbln1-GluD2と比較すると結合が弱いと推測されるが,類似の相互作用によって複合体を形成すると予想される.
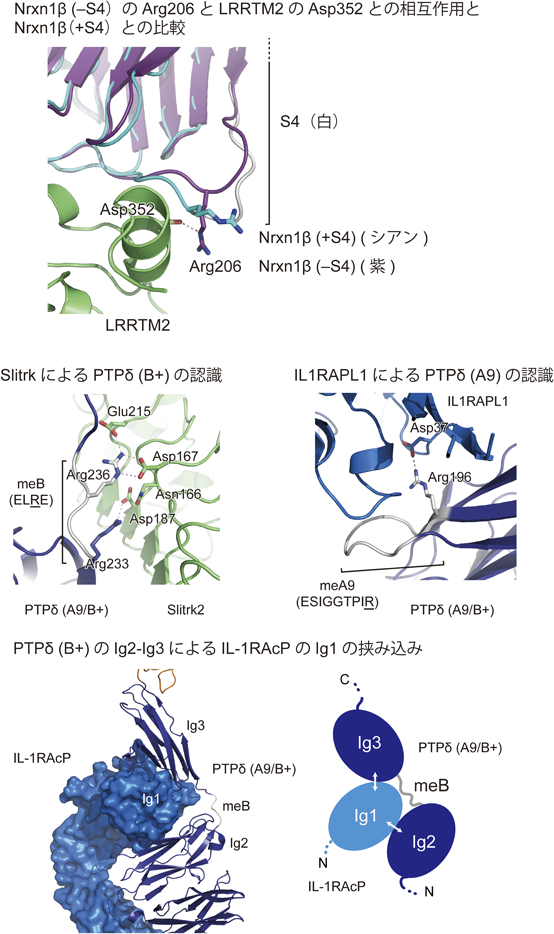
一方,筆者らは,–S4依存的なLRRTM1/2との相互作用機構を明らかにするために,Nrxn1β(−S4)LNS-LRRTM2 LRR複合体の結晶構造を決定した(図3;上段中)28).細胞外もしくは細胞膜上で機能するLRRには,リピート領域のN末端側とC末端側にジスルフィド結合を含むキャップ構造が存在して,リピート構造を安定化しているが,LRRを介したタンパク質間相互作用では,一般的にリピート領域が形成する凹面が利用される.しかし,Nrxn1β(−S4)とLRRTM2の相互作用では,珍しいことに,C末端側のキャップ構造が利用されている.この相互作用にはCa2+が必要であるが,NrxnとNlgnの相互作用と同様にCa2+が分子間インターフェイスに配位している.Nrxn1β(+S4)の立体構造との比較では,S4の挿入は相互作用に重要なNrxn1βのアルギニン残基のコンホメーションを変化させ,その結果として結合が阻害されることが示唆された(図4;上段).実際に,このアルギニン残基をアラニン残基に置換した変異体とNrxn1β(+S4)は,LRRTM2のECDに対して同程度の親和性で結合する.スプライスインサートの欠失によって,局所的なコンホメーションを変化させることで,親和性とシナプス形成の誘導能が調節されている.
2)IIa型RPTP
Nrxnが単独のドメイン(LNSドメイン)を介してシナプス後終末のシナプスオーガナイザーと相互作用するのに対して,IIa RPTPは,細長く伸びたECDの先端に位置する三つのIgドメインを介してSlitrkやIL-1RAcP, IL1RAPL1, SALM, TrkCと相互作用する.SlitrkのECDは二つのLRRドメイン(LRR1およびLRR2)で構成され,LRR1がIIa型RPTPとの結合とシナプス誘導能を担う.韓国KAISTのKimと延世大のKoの共同グループは,PTPδ(A9/B+)-Slitrk1 LRR1複合体の結晶構造を,また,筆者らはPTPδ(A9/B+)-Slitrk2 LRR1複合体の結晶構造(図3;中段左)を決定して,meBインサートの3残基目と1残基前のアルギニン残基がSlitrkにより特異的に認識されていることを明らかにした(図4;中段左)30, 31).B–のスプライスバリアントは,Slitrkと結合せず,シナプス形成誘導能も持たない.meBのアミノ酸配列は,シンプルに目印としての役割を果たしている.
一方,筆者らはPTPδ(A9/B+)-IL1RAPL1複合体とPTPδ(A9/B+)–IL-1RAcP複合体の結晶構造(図3;中段中)から,IL1RAPL1がmeAインサートのC末端に位置するアルギニン残基を認識すること(図4;中段右),また,IL1RAPL1やIL-1RAcPと相互作用する2番目と3番目のIgドメインの適切な空間配置をmeBインサートが制御していることを示した(図4;下段)29).Ig1とIg2は,密なドメイン間相互作用によってV字形の構造ユニットを形成することで単一のドメインのように振る舞うが,それに続くIg3は,meBの有無に応じてIg1-Ig2のユニットに対して相対的な配置が変化する.この空間配置を制御することで,PTPδのIg1-Ig2とIg3は,IL1RAPL1とIL-1RAcPのL字形構造の先端に位置するIgドメインを挟み込むように相互作用することが可能となる(図4;下段).meAは,Slitrkとの相互作用におけるmeBと同様に,目印としての役割を果たすのに対して,meBは,挟み込みによる認識に関与する二つのドメインの空間配置を制御する役割を担う.実際に,meBの配列を変えたPTPδ変異体では,IL1RAPL1やIL-1RAcPに対する親和性はほとんど変化しないが,長さを2倍や3倍にした変異体では,親和性が低下する.PTPδとの結合に関してmeBへの依存度が高いIL-1RAcPの方が,IL1RAPL1よりも低下の度合いは大きい.
同様な挟み込みによる認識は,PTPδとSALM5との相互作用でもみられる.SALMのECDは,LRR, Ig, FNの三つのドメインで構成され,LRR-IgがIIa RPTPとの相互作用を担う.筆者らと北京大学のLiuの研究グループは,それぞれ独立にPTPδとSALM5 LRR-Igの複合体の結晶構造を決定した(図3;中段右)32, 33).PTPδ Ig2とSALM5 LRRの相互作用に加えて,PTPδのIg1-Ig2とIg3がSALM5 Igを挟み込むように認識していた.meBの有無で,PTPδとSALM5の結合親和性は1桁近く変化することから,IIa RPTPとSALM5によるシナプス形成誘導もmeBの選択的スプライシングによって調節される可能性が示唆される.
3) Ten
TenのECDは,EGFリピートを介して二量体化するが,EGFリピート以外の領域での二量体間の相互作用はなく,各プロトマーは独立に運動していると考えられる.シカゴ大のAraçとスタンフォード大のSüdhof, Skiniotisの共同グループは,EGFリピートを除いて単量体化したTen2 ECD(ΔEGF)の立体構造を低温電子顕微鏡(cryo-EM)単粒子解析によって決定している38).また,オックスフォード大のSeiradakeのグループは,より高分解能の立体構造をX線結晶構造解析によって決定している(図3;下段左)37).Tenは,一般的な1回膜貫通型の受容体と異なり,C末端側が細胞外に露出したII型の膜タンパク質であり,また,真核生物の他の膜受容体との相同性はない.しかし,バレルとToxin様の両ドメインは,細菌のTc毒素のBCコンポーネントと相同である.Tenとその相互作用相手であるLphnの複合体の立体構造は,現在までのところは報告されていないが,TenのバレルとToxin様ドメインの周辺で相互作用すると予想されている.スプライス部位はβプロペラドメイン内に存在していることから,βプロペラドメインを介して相互作用する未同定のシナプスオーガナイザーの存在も予想されているが,筆者の知る限りでは,それを実験的に支持する報告はない.
上述したように,シナプス形成誘導において,TenはFLRTと共依存的に機能する.LphnのECDは,レクチン,Olfactomedin(Olf),Horm/GPCR autoproteolysis-inducing(GAIN)の三つのドメインで構成され,FLRTとの結合は,Olfドメインとの相互作用を介して行われる.前述のTen2 ECD(ΔEGF)のcryo-EM構造を決定した研究グループとラトガス大のComolettiのグループが,それぞれ独立にFLRT3 LRRとLphn3 Olfの複合体の結晶構造を報告している44, 45).球状のLphn Olfは,FLRT3 LRRの凹面に収まるように相互作用する(図3;下段右).
4. シナプスオーガナイザーによるシナプス分化誘導の分子機構
シナプスオーガナイザーのトランスシナプティックな相互作用は,シナプス前終末でのAZの構築とSVの蓄積,シナプス後終末でのPSDの構築を促進して,シナプス形成を誘導すると考えられる.メガダルトン(MDa)級の大きさを持つAZやPSDは,これらを構成する足場タンパク質が高密度に凝縮したものであると考えられる.近年,生体分子の凝縮が液–液相分離(liquid-liquid phase separation)によって起きることがさまざまな細胞機能で示されてきているが,シナプスにおいても,AZやPSDを構成する因子群が相互作用依存的に液-液相分離を生じて凝縮することを香港科技大のZhangのグループが報告している46–50).シナプスで生じる分子の凝縮に類似した現象は,ガラス基盤で支持した脂質膜上で再現されており,また,再現された現象を阻害する要因が,神経細胞においても同様の効果を示すことも確かめられている.このような実験系では,細胞外での相互作用の要素は含まれていないことから,AZやPSDの構築は細胞外での相互作用がなくても開始される可能性が考えられる.シナプス形成が誘導される前の段階でAZやPSDを構築する準備は始まっており,そこに細胞外でのトランスシナプティックな相互作用が加わることで,上述の「ナノカラム」が構築され,シナプスとしての機能が獲得されるのかもしれない.
シナプス形成には,膜に平行な面に沿って広がるシナプスオーガナイザーのクラスタリングが必要であることが示唆されているとともに,一部のシナプスオーガナイザーでは二量体化が必要なことも示されている32, 33, 39–41, 51).接着状態での分子全体の立体構造と会合状態,それらの生理的意義と使い分けなど,多くの課題が残されている.また,本稿では詳細にはふれなかったが,糖鎖(ヘパラン硫酸鎖)がシナプス形成の誘導に重要な役割を担っていることも明らかになっており,その機能メカニズムの理解も必要である52).
5.てんかん関連リガンド-受容体複合体LGI1-ADAM22
興奮性シナプス伝達と抑制性シナプス伝達のバランス調節の破綻は,自閉症や知的障害などの神経発達障害にとどまらず,より劇的な症状として致命的な痙攣を伴うてんかん発作を引き起こす.てんかんの原因遺伝子産物のほとんどは,膜電位を制御するイオンチャネルであるが,非イオンチャネル型の原因遺伝子産物として,神経細胞の分泌タンパク質であるLGI1が知られている53).LGI1遺伝子の変異は,遺伝性てんかんの一つである常染色体優性外側側頭葉てんかん(autosomal dominant lateral temporal lobe epilepsy:ADLTE)を引き起こす54–56).ヒト以外の動物でも,LGI1とてんかん病態との関連が遺伝子改変動物により示されている.たとえば,LGI1のホモノックアウトマウスは,重篤なてんかん発作の繰り返しによって,生後3週間以内に死亡するが,この表現型はLGI1の入れ戻しによって回復する57).
LGI1は,膜結合型メタロプロテアーゼであるADAMファミリーに属するADAM22およびADAM23を受容体として複合体を形成する.この二つにADAM11を含めた3種類のメンバーでADAM22サブファミリーを構成する.ADAM22の細胞内領域は,シナプス後終末のPSDの中心的な構成分子である足場タンパク質PSD-95と相互作用し,AMPA型グルタミン酸受容体のアクセサリータンパク質Stargazinを介して,その機能を促進することが報告されている58, 59).また,シナプス前終末上のADAM23とシナプス後終末上のADAM22がLGI1を介してトランスシナプティックな相互作用をすることで,正常なシナプス伝達を維持する可能性が示唆されている.このモデルは,上述のナノカラムのコンセプトとも合致する.LGI1に対する自己抗体は,痙攣を伴う辺縁系脳炎を引き起こすが,自己抗体とLGI1との結合は,LGI1とADAM22との相互作用を阻害することも報告されている60, 61).
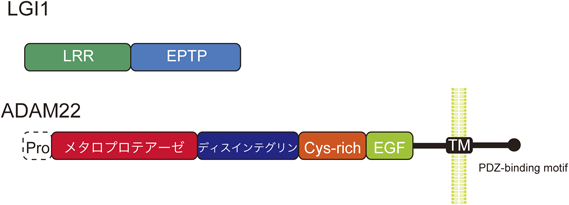
てんかんの原因となるLGI1のミスセンス変異の多くは,LGI1の分泌異常を引き起こす62).これまでの報告から筆者が把握している29種類の変異のうち,23種類で分泌能が調べられており,そのうちの20種類で分泌不全が確認されている.LGI1は,N末端側のロイシンリッチリピート(LRR)とC末端側のエピテンピンリピート(EPTP)の二つのドメインで構成されているが(図5),分泌不全を示す変異は両方のドメインに偏りなく見いだされている.後述するLGI1の立体構造に基づくと,構造の安定化に寄与するジスルフィド結合を担うシステイン残基やドメイン構造内部の疎水性残基など,フォールディング異常を引き起こす可能性の高い変異が多くみられる63).分泌不全は,LGI1のフォールディング異常に起因し,小胞体品質管理によって排除されていると考えられる.実際に,フォールディング異常の改善効果を持つケミカルシャペロンを投与することで,分泌異常を示すE383A変異を持つてんかんモデルマウスの生存期間を延ばすことができる62).
1)LGI1とADAM22の相互作用
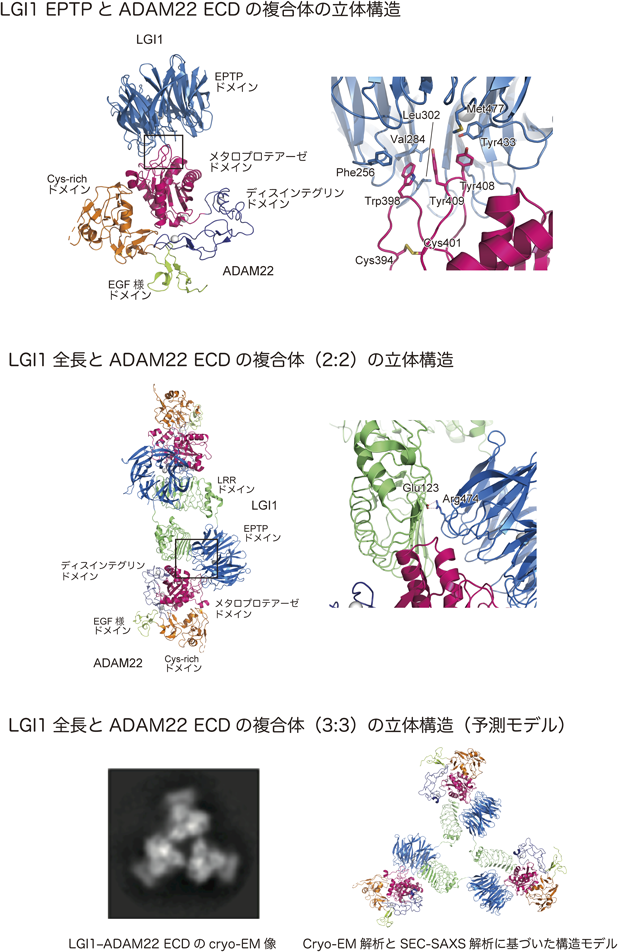
ADAM22のECDは,メタロプロテアーゼ様,ディスインテグリン,システインリッチ(Cys-rich),EGF様の四つのドメインで構成される(図5).このうち,メタロプロテアーゼ様ドメインがLGI1のEPTPドメインと相互作用する.この相互作用様式を明らかにするために,筆者らは,LGI1 EPTPとADAM22 ECDの複合体の結晶構造を決定した(図6;上段左)63).LGI1 EPTPは,七つのブレードで構成されるWD40リピート様のβプロペラ構造を有している.四つのβストランドが逆平行に並んだ各ブレードの構造は,典型的なWD40のブレードとはWDモチーフの位置が2残基ずれた非古典的なブレード構造であった.また,構造的な特徴として,内部にジスルフィド結合を持ち,中央の穴にカルシウムイオンが配位することで構造が安定化されている.
LGI1 EPTPのβプロペラ構造の周縁部に位置する疎水性のポケットが,ADAM22のメタロプロテアーゼドメインから突き出した二つのループ上の三つの芳香族残基と結合する(図6;上段右).この相互作用がLGI1 EPTPとADAM22の結合に不可欠であることは,部位特異的変異体を用いたプルダウン解析により確かめられた.この二つのループは,LGI1との結合によって,そのコンホメーションを変化させるが,一つのループの根元を安定化するようにジスルフィド結合が形成されている(図6;上段右のCys394とCys401).このジスルフィド結合を失う変異は,難治性のけいれんと深刻な知的障害を伴う重篤な脳症を引き起こすことが報告されている64).
LGI1 EPTPとの相互作用に関わるアミノ酸残基のほとんどは,ADAM23でも保存されており,それらの部位特異的変異体を用いたプルダウン解析の結果は,ADAM22と同様である.ADAM22とADAM23の配列保存性の高さと合わせて,ADAM23とADAM22は同様の相互作用様式でLGI1と結合することが示唆される.
2)LGI1とADAM22による高次会合体の形成
てんかん症状との関連が報告されているLGI1のミスセンス変異のうち,3種類の変異体R407C, S473L, R474Qは分泌能を維持している.R407C変異体については,S473L変異体やR474Q変異体とは異なり,遺伝子の入れ戻しによってLGI1をノックアウトしたてんかんモデルマウスの生存期間を野生型遺伝子と同等に回復できること,また,重篤な疾病変異を除外してあるゲノムデータベースであるgnomADに5例のR407C変異が登録されていることから,筆者らは,R407Cは異常を生じない変異であると結論づけた63).一方,S473L変異体は,ADAM22との結合能を失っていることが明らかになっている62).Ser473は,ADAM22との相互作用に直接関与はしていないが,相互作用面の近傍に位置しており,間接的に相互作用面へ影響を及ぼすことが示唆された.これに対して,R474Q変異は,ADAM22との相互作用にも顕著な異常はみられず,病態を生じるメカニズムは不明であった.残された可能性は,先に述べた,LGI1を介したADAM23とADAM22のトランスシナプティックな相互作用への影響であり,そのためには,LRRドメインも含めたLGI全長を含む複合体の会合状態を解析することが重要であると考えられた.
生体高分子の会合時の分子量は,会合状態を予測する上で重要な情報である.分子量を精度よく測定するために有効な手法の一つが,ゲル濾過クロマトグラフィーと多角度光散乱を組み合わせた解析(SEC-MALS)であり,この解析手法を用いてLGI1とADAM22との会合状態を解析した63).LGI1 EPTPとADAM22 ECDの複合体を解析した結果は分子量11万であり,1 : 1で複合体を形成していることを示していた.一方,LGI1全長とADAM22 ECDの複合体を解析した結果は,500 mM NaCl存在下で分子量27万,150 mM NaCl存在下で分子量36万であり,2 : 2もしくは3 : 3で高次の会合体を形成することを示していた.そして,興味深いことに,この高次の会合体の形成は,R474Q変異によって阻害される(分子量13万).
3)LGI1とADAM22複合体の立体構造
SEC-MALSで検出されたLGI1とADAM22の高次の会合体は,LGI1を介してトランスシナプティックに形成されるADAM23とADAM22の複合体に対応すると考えられる.筆者らは,試行錯誤の結果,LGI1全長とADAM22 ECDの複合体の結晶構造を決定した(図6;中段左)63).分解能は,通常のX線結晶構造解析で得られる分解能(<3 Å)には及ばなかった(7 Å)が,LGI1 LRRとLGI1 EPTP-ADAM22 ECDの高分解能構造を用いることで電子密度を解釈することができた.二つのLGI1分子が相互作用することで,二つのADAM22分子を橋渡しした全体構造をしており,長軸の長さ19 nmは,興奮性シナプスのシナプス間隙の大きさに対応する.ADAM22とADAM23がLGI1に同じように結合すると考えると,この結晶構造は,LGI1を介したADAM23とADAM22のトランスシナプティックな相互作用を反映した立体構造といえる.LRRモチーフは,一般的にタンパク質−タンパク質間相互作用に用いられるが,LGI1のLRRドメインは,単離された状態では会合能を示さない.明らかになった2 : 2の立体構造では,LGI1 LRRは,もう一方のLGI1 EPTPと相互作用するとともに,ADAM22とも相互作用している.そのために,LGI1のみでは二量体を形成できないと思われる.
てんかん発症の原因となるR474Q変異の周辺の構造に着目すると,Arg474は,もう一つのLGI1分子のGlu123と水素結合を形成していることが示唆された(図6;中段右).E123K変異もてんかんを発症する変異として同定されている.基本的には分泌異常による発症と考えられるが,高次会合体形成への影響も複合的に寄与している可能性もある.LGI1によるADAM22とADAM23の橋渡しにR474Q変異が影響を与えていることは,R474Q変異体を発現するLGI1ノックアウトマウスの脳抽出液を使ったプルダウン実験によって確かめられた.ADAM22もしくはADAM23をプルダウンすると,野生型のLGI1もR474Q変異体もともに共沈降する.これは,R474Q変異体がADAM22やADAM23との結合自体は阻害しないことを示す.一方,ADAM22をプルダウンしてADAM23の共沈降を調べた場合,もしくは,逆にADAM23をプルダウンしてADAM22の共沈降を調べた場合では,野生型とR474Q変異体で明確な違いがみられた.R474Qでは共沈降が起きなかったことから,立体構造から想定されるようなLGI1によるADAM22とADAM23のトランスシナプティックな橋渡しが,脳内の神経細胞でも阻害されていることが確かめられた.
4)LGI1とADAM22の高次会合体の構成
2 : 2のLGI1-ADAM22 ECD会合体の立体構造に基づいてトランスシナプティックな橋渡しのメカニズムが明らかになったが,SEC-MALSの解析では,細胞外環境に近いNa濃度(150 mM NaCl)存在下では,溶液中で3 : 3の会合体を形成することが示唆された.一方,R474Q変異体やEPTPのみのLGI1では,塩濃度による会合状態の変化はみられなかった.2 : 2と3 : 3の会合状態をSEC-MALS以外の方法で検証するために,筆者らは,分子の概形に関する情報が得られるX線小角散乱測定(small angle X-ray scattering:SAXS)を行った63).MALSと同様に,SECと組み合わせたSEC-SAXSという手法を用いた.得られた散乱カーブは2 : 2の立体構造とはよいフィッティングを示さず,SEC-MALSの場合と同様に3 : 3の会合体の可能性が示唆された.さらに会合状態を突き詰めるために,cryo-EMによる観察を行った.想定外に1 : 1に対応する粒子が多く観察された(観察した全粒子の60%)が,急速凍結の際に会合体が壊れている可能性もある.一方,35%の粒子が2 : 2の会合体に対応するものであり,結晶構造でみられた2 : 2の会合体が溶液中でも存在することが示された.3 : 3の会合体は,残りの5%の粒子でみられた(図6;下段左).割合は少なかったものの,最も明瞭な二次元投影像が得られたことから,構造的なゆらぎの少ない安定な状態として存在していることが示唆される.特徴的な3回対称を持った二次元投影像と結晶構造から抽出したLGI1 LRR/LGI1 EPTP/ADAM22 ECDの構造単位を利用すると,3 : 3の会合体モデルを作製することができる(図6;下段右).この会合体モデルから予測される散乱曲線は,実測した曲線とよくフィットしたことから,3 : 3の会合体の存在が裏づけられた.溶液散乱は径の大きな粒子の影響を反映しやすいこと,また,cryo-EMは急速凍結による複合体の解離が起きうることを考慮すると,溶液中あるいは細胞上での1 : 1, 2 : 2, 3 : 3の会合体の存在比を安易に予測することは避けなくてはならないが,これらの一連の実験は,3 : 3の会合体が安定な構造状態で存在することを強く示唆している.
5)LGI1とADAM22の高次会合体の機能
2 : 2の会合体の立体構造がトランスシナプティックな橋渡しを合理的に説明するのに対して,3 : 3の会合体の立体構造と機能は,合理性に欠ける印象を持つかもしれない.橋渡しの様式を変化させることで,シナプス前終末のAZやSVの密度とシナプス後終末のPSDの密度を調節するモデルや,同一膜上(いわゆる,シスと呼ばれる状態)で3 : 3の複合体を形成して,自己阻害状態となるモデルなどを想像することはできるが,これらのモデルの根拠となるような実験データは報告されていない.一方で,トランスシナプティックな橋渡しとは異なる機能を考慮する必要があるかもしれない.
近年,LGI1が軸索起始部(axon initial segment:AIS)と呼ばれる細胞体近くの軸索部位に集積して,ADAM22/23や電位依存性Kチャネル(Kv1)と共局在していることが報告されている65, 66).この共局在は,Kv1と相互作用する細胞接着因子TAG-1やCaspr2を介していると考えられている.AISは,活動電位の発生部位として,シナプス前終末での信号出力を決定する要因の一つであり,過剰な興奮はてんかん様の発作につながる可能性が考えられるが,Kv1の集積は,過興奮を抑制する方向で働くと考えられている.R474Q変異体は,ADAM22/23やKv1との局在を減じることから,AISの過興奮を引き起こしている可能性もある.3 : 3の会合体は,各プロトマーの配置からシスの様式で機能するのが合理的に思われるが,AISの膜上でKv1の集積を促進する役割を担っているのかもしれない.一方で,細胞接着因子TAG-1やCaspr2とADAM22/23との相互作用は弱い印象であり,Kv1とADAM22/23が共局在する別の分子機構が存在する可能性もある.