膜タンパク質の動的構造解析Molecular Movies of Membrane Proteins
京都大学大学院医学研究科Department of Cell Biology. Graduate School of Medicine, Kyoto University ◇ 606–8501 京都市左京区吉田近衛町 ◇ Yoshida-konoe, Sakyo-ku, Kyoto 606–8501, Japan
発行日:2020年4月25日Published: April 25, 2020
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
膜タンパク質の構造生物学はいま大きな転換期を迎えている.近年,電子顕微鏡の検出器の進歩により,多くの膜タンパク質の構造が結晶を用いずに解かれるようになった.2019年に電子顕微鏡を使って解かれた膜タンパク質構造は276個で,これに対しX線を使って解かれたものは457個であり(http://www.wwpdb.org)1),この数は数年のうちには逆転するだろうと予想されている.これまで膜タンパク質の構造解析の最大のボトルネックは結晶化であったが,これは解消されつつある.電子顕微鏡による構造解析については筆者の専門ではないし,多くのすぐれた総説もあるのでそちらを参考にされたい(たとえば,小誌88巻4号2)).本稿では,膜タンパク質研究にいまだ結晶を使ってメリットのある分野,X線自由電子レーザーを用いた動的構造解析について解説したい.我々はこの手法を“高速分子動画法”と呼んでいる.
図1に高速分子動画法についてまとめた.X線自由電子レーザーはこれまでのX線光源とはまったく異なった新しいタイプの光源である.放射光が数十psのパルスの中に105個程度のフォトンを有するのに対し,日本のX線自由電子レーザーSACLA(SPring-8 angstrom compact free electron laser)においては,10 fs程度のパルス中に1011個以上のフォトンを有している.これは1回の10 fsパルスだけでタンパク質結晶からの回折像を得ることができることを意味している.タンパク質に非常に強いX線を照射した場合はX線により電子がけり出され,原子がプラズマ化し,クーロン力によって爆発することが知られている.しかしながら原子核には質量があるため,数十fsの間は元の位置より大きく動くことはできない.また,この時間は実際に原子間の結合が切れる時間にも対応している.つまり10 fsの間に回折像が得られれば,X線の照射によって化学結合が切断される前の構造を得ることができると考えられる3).これは動いている被写体に対してストロボライトで撮影し,一瞬の静止画を切り出すのとよく似ている.ただ1枚のX線回折像は基本的に分子の投影図の情報に近いものしか有していないため,いろいろな方向からとった回折像を組み合わせることにより,三次元再構成を行う.このような構造解析の方法をシリアルフェムトセカンド結晶解析法(SFX)と呼んでいる4)(この最初の論文ではfemtosecond X-ray protein nanocrystallographyと呼ばれている).SFX自身は,タンパク質の静的な構造を切り出す手法にすぎない.これに対して,まず何らかの方法でタンパク質結晶の中で構造変化または化学反応を開始させ,その一定時間後にX線自由電子レーザーを照射し,その構造を決めるポンプ・プローブ実験を行う.ポンプ(励起)とプローブ(検出)の間の時間を変えることにより,反応開始から一定時間後の一連の構造を得ることができ,これを並べたタイムラプスイメージ(パラパラ漫画のようなものを想像していただくとよい)を作ることができる.我々はこの方法を“高速分子動画法(Molecular Movies)”と呼んでいる.本法は,現時点では結晶中で可能な反応・変化だけしか追えないという限界があるが,空間分解能としては原子分解能(つまりオングストロームレベル)で,時間分解能としては数十fs程度で生体高分子の構造変化や化学反応を追うことのできる大変ユニークな方法である.
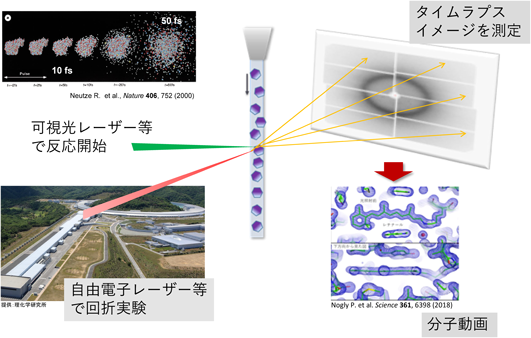
図2に我々が使用している高速分子動画法用の回折装置を示す.この装置は理化学研究所によって開発されDAPHNIS(diverse application platform for hard X-ray diffraction in SACLA)と呼ばれる5).ヘリウムで置換された回折チェンバーと高速読み出し可能なmpCCD(複数のCCDを組み合わせて読み出しを早くしたもの)で構成されている.回折チェンバーの上部にはサンプル導入のためのインジェクターと呼ばれる装置がつけられている6).目的によって異なったインジェクターがあるのだが,膜タンパク質の場合は,高粘度溶液用インジェクターと呼ばれるものを用いることが多い(図3).近年,膜タンパク質の結晶はその多くが脂質キュービック相(LCP)と呼ばれる粘稠な媒体中で作られる.LCP中の結晶を回折チェンバーに導入するためには,圧力をかけてゆっくりと絞り出すようにノズルの先から出す必要があり,それがまっすぐ吐出されるように,外側からガイドするガスを送り,さらに下からポンプで引く形をとっている.一見,非常に困難なことを行っているように感じられるが,実は粘度の低い液体を吐出するのに比べて流速をとてもゆっくりとすることが可能なため,サンプルの消費量を抑えることができ,SFXの実験に非常に適したシステムとなっている.このインジェクターで微結晶(通常数マイクロメートル程度)を導入することにより,結晶はランダムな向きで自由電子レーザーのビームで照射されることになる.通常1万個程度の結晶からデータを集めることにより,三次元再構成に必要なデータを得ることができる.


右はサンプルのストリームの拡大図.
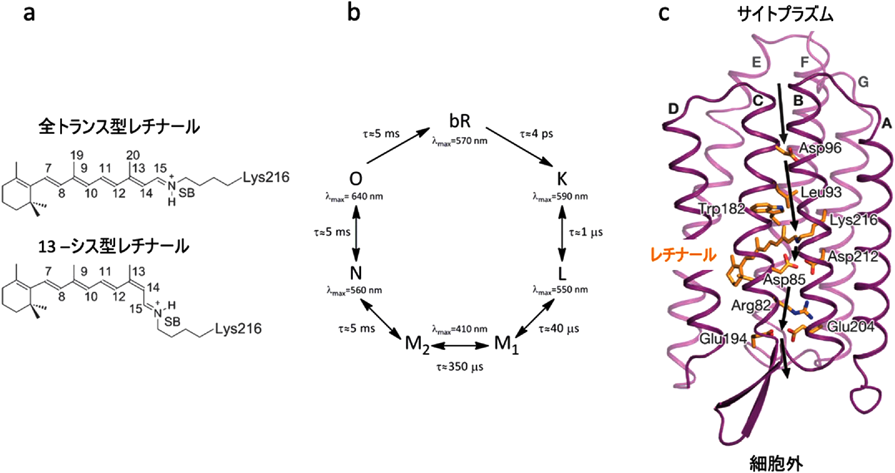
我々が最初にこのシステムのターゲットに選んだのは,光駆動性プロトンポンプ,バクテリオロドプシン(bR)である.bRは図4aにあるように,結合したレチナールが光照射により全トランス型から13-シス型に異性化した後に,図4bのような光サイクルを経て元の基底状態(bR状態)に戻る.その間に水素イオンを細胞膜の内側から外側に輸送する(図4c).bRはこれまでクライオトラップや変異酵素を用いた各種の実験でその中間体について調べられているが,これらの中間体は人為的に作製されたもので実際の中間体とは異なるため,プロトンポンプの機構については現在でも論争がある.高速分子動画法を用いて構造変化を実時間観察し,この問題の決着をつけるべく実験を行った7).実験には図5aに示したような装置を用いた.基本的にDAPHNISにポンプ光用の可視レーザーをつけた装置で,図5bのようにポンプ光で励起後一定の時間にX線レーザーでデータをとるということを繰り返し,一つのタイムポイントのデータを集める.そして次にディレイの時間を変えて同じ実験を繰り返し,次のタイムポイントのデータを集める.これを繰り返し,我々は20 nsから1725 µsまでの13点のタイムポイントのデータを集めることに成功した(図6).分子動画法と他の動的構造解析手法との違いは,非平衡状態を実時間観察していることである.図6には構造の変化を示す差フーリエ図を示してあるが,レチナールで起こった構造変化は約100 µsでタンパク質の表面まで伝わると考えられる.これは基本的に熱振動の伝わる速さだと考えられる.この後だんだんと差フーリエは小さくなり,元の基底状態に戻る.順を追って構造変化をみていくと,20 ns後には,すでにレチナールは13-シス型になっていることがわかる(図7a).このときシッフ塩基を形成しているLys216の側鎖の結合の回転と,レチナールの平面構造からねじれを引き起こす.290 ns後には,レチナールが平面構造に戻ると同時にLys216全体が動いており,それに伴いヘリックスが動き,またシッフ塩基に水素結合していた水分子が消失している.さらに760 ns後の構造をみると左側のヘリックスが動き始めると同時に,シッフ塩基とThr89をつなぐ水分子W452が現れているのがわかる(図7b).Thr89はさらにAsp85に水素結合しており,シッフ塩基からAsp85へのプロトンの移動に重要な役割を果たしていると考えられる.36 µs後には先ほど現れた水分子は消失しており,それと同時にThr89-Asp85の水素結合が切れ,Asp85の側鎖が回転を始める(図7c).Arg82がコンホメーションを変えるとともに新しい水分子W451が現れ,これがさらに水分子の鎖を通じて水素イオンを細胞膜の外に輸送するのに用いられると考えられる.
[Nango, E. et al. (2016) Science, 354, 6319より.]
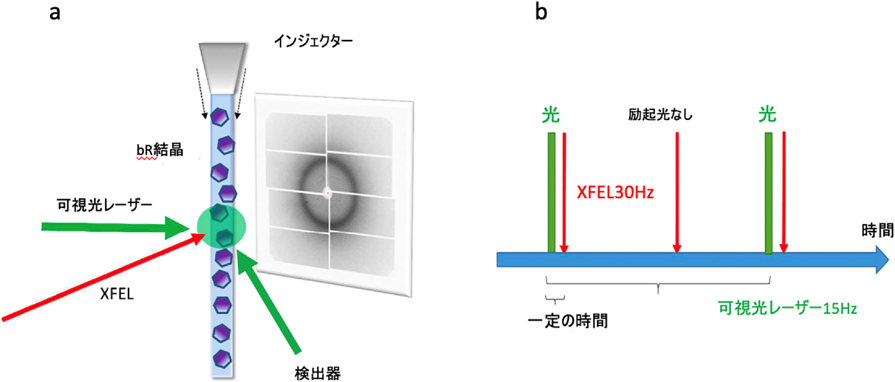
(a)実験のセットアップ.(b)パルスシークエンス.[Nango, E. et al. (2016) Science, 354, 6319より.]
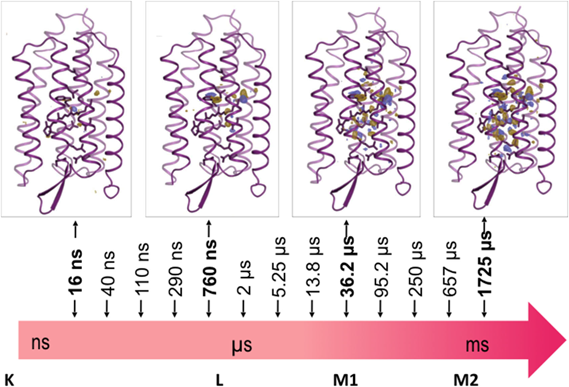
K, L, M1, M2は,図4bに示した状態に対応する.[Nango, E. et al. (2016) Science, 354, 6319より.]
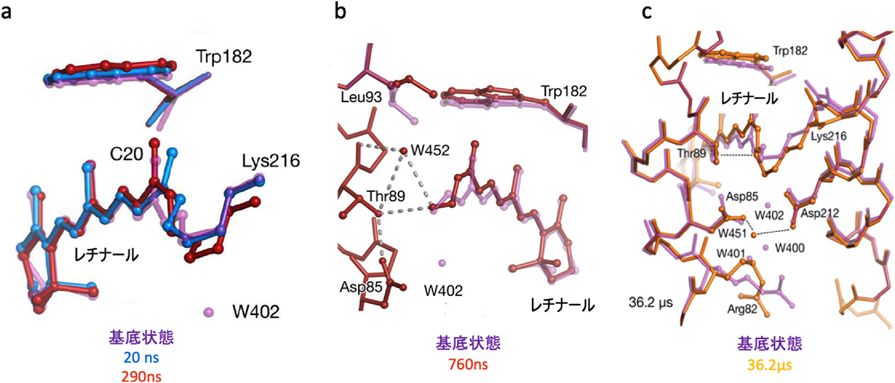
基底状態と反応開始より20 ns後,290 ns後,760 ns後および36.2 µs後の構造を比較した.[Nango, E. et al. (2016) Science, 354, 6319より.]
我々はさらにbRの20 ns以前の構造変化を捉えることを試みた8).この実験は,共同研究者のスイスのポール・シェーラー研究所(PSI)のグループとともにアメリカの自由電子レーザーLCLSを用いて行われた.我々はこの実験により反応開始から0–142 fs後(この時間の間の構造が混ざっているという意味)および457–547 fs後の構造および10 ps後の構造を解析することに成功した(図8).10 ps後の構造は前述の20 ns後の構造と基本的に同じK状態で,すでにレチナールが13-シスのコンホメーションをとっている.今回はそれに達する以前のI状態およびJ状態に相当する構造の情報が得られた.I状態ではレチナール上で電荷の分離が起き,シッフ塩基近傍に負電荷が存在する形となり,この電荷により,レチナールの二重結合が回転可能な状態(J状態)になり異性化が起こると考えられている.我々のデータではIおよびJ状態においてシッフ塩基に水素結合していた水分子(W402)とそれに水素結合するAsp212の側鎖が移動することが観測された.これは移動した電荷および結合の回転に伴う立体障害の影響であると考えられ,K状態では水分子の位置もAsp212の側鎖の位置も基底状態に戻っていた.このような動きはこれまでまったく予想・観察されていなかったものであり,高速分子動画法の卓越した位置・時間分解能によって初めて観測可能となったものである.
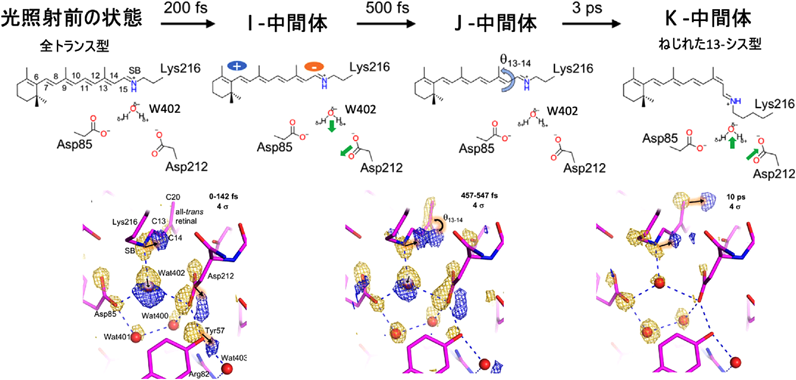
[Nogly, P. et al. (2018) Science, 361, 6398より.]
今回bRの例で示したように,高速分子動画はこれまで解明の進んでいなかった膜タンパク質の動的過程の解明に対する非常に強力なツールである.ここに示した例だけでなく,すでに光合成における水分解過程の中間構造を示すことにも成功しており9),また視覚をつかさどる光受容体ロドプシンや光遺伝学のツールの機構解明の実験も行われ,これらの成果も期待されている.我々はこの高速分子動画法の適用をより多くのタンパク質に拡げるために,新学術領域“高速分子動画法によるタンパク質非平衡状態構造解析と分子制御への応用(高速分子動画)”を立ち上げた(http://www.molmovies.med.kyoto-u.ac.jp).本領域では光反応性でないタンパク質に対しても高速分子動画法を適用できるような技術を開発することを目指している.たとえば光でスイッチできるリガンドをデザインして光感受性でない受容体を活性化したり,光以外の刺激(溶液の混合,温度ジャンプなど)で反応を開始したりするシステムを開発することを目指している.この目的のためにタンパク質工学やケミカルバイオロジーを駆使する.またタンパク質の動的解析だけでなく,新規機能性タンパク質や化合物の開発を行い,光遺伝学や光薬理学のツールとして供することを目指している.多くの光遺伝学のツールは膜タンパク質であり,医薬の50%以上は膜タンパク質をターゲットにしていることを考えると,これらの新規機能性タンパク質・化合物は基礎生物学や創薬にとって重要なツールになりうる.高速分子動画法を核として膜タンパクの動的構造に関する理解を深め,それを応用にもつなげていきたいと考えている.
1) Berman, H.M., Henrick, K., & Nakamura, H. (2003) Announcing the worldwide Protein Data Bank. Nat. Struct. Biol., 10, 980.
2) 光岡 薫,Christoph Gerle (2016) 低温電子顕微鏡法を用いた単粒子解析の最近の進展と膜タンパク質への応用,生化学,88, 532–536.
3) Neutze, R., Wouts, R., van der Spoel, D., Weckert, E., & Hajdu, J. (2000) Potential for biomolecular imaging with femtosecond X-ray pulses. Nature, 406, 752–757.
4) Chapman, H.N., Fromme, P., Barty, A., White, T.A., Kirian, R.A., Aquila, A., Hunter, M.S., Schulz, J., DePonte, D.P., Weierstall, U., et al. (2011) Femtosecond X-ray protein nanocrystallography. Nature, 470, 73–77.
5) Tono, K., Nango, E., Sugahara, M., Song, C., Park, J., Tanaka, T., Tanaka, R., Joti, Y., Kameshima, T., Ono, S., et al. (2015) Diverse application platform for hard X-ray diffraction in SACLA (DAPHNIS): application to serial protein crystallography using an X-ray free-electron laser. J. Synchrotron Radiat., 22, 532–537.
6) Mizohata, E., Nakane, T., Fukuda, Y., Nango, E., & Iwata, S. (2018) Serial femtosecond crystallography at the SACLA: breakthrough to dynamic structural biology. Biophys. Rev., 10, 209–218.
7) Nango, E., Royant, A., Kubo, M., Nakane, T., Wickstrand, C., Kimura, T., Tanaka, T., Tono, K., Song, C., Tanaka, R., et al. (2016) A three-dimensional movie of structural changes in bacteriorhodopsin. Science, 354, 1552–1557.
8) Nogly, P., Weinert, T., James, D., Carbajo, S., Ozerov, D., Furrer, A., Gashi, D., Borin, V., Skopintsev, P., Jaeger, K., et al. (2018) Retinal isomerization in bacteriorhodopsin captured by a femtosecond X-ray laser. Science, 361.
9) Suga, M., Akita, F., Sugahara, M., Kubo, M., Nakajima, Y., Nakane, T., Yamashita, K., Umena, Y., Nakabayashi, M., Yamane, T., et al. (2017) Light-induced structural changes and the site of O=O bond formation in PSII caught by XFEL. Nature, 543, 131–135.

京都大学大学院医学研究科分子細胞情報学教授.農学博士.
1963年生まれ.91年東京大学大学院農学系研究科博士課程修了.農学博士.2000年より英国インペリアルカレッジ生命科学科教授(~15年).07年より京都大学大学院医学研究科教授,12年より理化学研究所放射光科学研究センターSACLA利用技術開拓グループ・グループディレクターを併任.
研究テーマと抱負最近は膜タンパク質構造解析と自由電子レーザーを使った時間分割構造解析を行っています.大事だけれど,人がやりたがらない研究をしたいと思っています.
ウェブサイトhttp://cell.mfour.med.kyoto-u.ac.jp/
趣味自転車を組み立てるのとヒルクライムのレースに出るのが好きです.

This page was created on 2020-03-05T09:08:47.504+09:00
This page was last modified on 2020-04-15T11:28:33.000+09:00
このサイトは(株)国際文献社によって運用されています。