大脳皮質局所神経回路の遺伝学的解析Genetic dissection of local neural circuitry in the cortex
マックスプランクフロリダ神経科学研究所Max Planck Florida Institute for Neuroscience ◇ One Max Planck Way, Jupiter, Florida, USA 33458
発行日:2020年4月25日Published: April 25, 2020
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
大脳皮質神経回路が,どのような原理で発達し,どのような構造を持つのかを明らかにすることは,大脳高次機能の構造的基盤を理解するための必須課題である.また,大脳皮質構築原理から得られる情報は,脳疾患の病因,病態の解明,さらに治療法の開発に資すると考えられており,大脳皮質神経回路の発達,構造面からの研究は,医学的見地からも大きな役割を担っている.近年の遺伝学的標識技術,イメージング技術の爆発的進歩に伴い,神経回路構造がこれまでにない解像度で解析されるようになり,神経回路の解剖学的研究は,新たな局面を迎えつつある.さらに,最近の単一細胞トランスクリプトーム解析法の発達によって,大脳皮質神経細胞タイプの驚くべき多様性が明らかになり,それに連動する神経回路種の多様性,特異性が予見され,その実体を明らかにする研究の進展が期待されている.しかしながら,各々の細胞タイプにおける結合特異性,大脳皮質内空間配置に関する研究は,技術的制約のため,いまだ緒についたばかりといっても過言ではない.特に,大脳皮質内の神経演算において重要な役割を果たす,抑制性介在神経細胞(抑制性細胞)と興奮性投射細胞(興奮性細胞)の間に形成される局所神経回路の構造的知見はほとんど明らかになっていない.本稿では,最近,我々が開発した,ある特定の種類の興奮性細胞に入力を送る抑制性細胞をサブタイプ特異的に標識するための遺伝学的手法を紹介する.さらに,この手法を発展させ,展開しうる,局所神経回路研究に関する今後の展望を述べる.
大脳皮質神経細胞の約8割を占める興奮性細胞は,グルタミン酸を神経伝達物質として用い,後シナプス細胞を電気的に興奮させる作用を持つ.興奮性細胞は,一般に,長い軸索を投射し,層,カラム,領野といった大脳皮質の機能単位における演算結果を離れた脳部位に伝達し,皮質内,皮質下の異なる神経回路システムへの信号伝達に主要な役割を果たしている.大脳皮質内の興奮性細胞の大部分を占める錐体細胞は,錐体形の細胞体,脳表面へ向けて伸ばされた尖端樹状突起,細胞体の周りに局所的に形成される基底樹状突起,1本の長い軸索からなる基本形態を有する.さらに,樹状突起は,生化学的,生物物理学的に特殊な性質を備えた細胞内区画であるスパインと呼ばれる小さな棘状突起を無数に持つ.興奮性入力は,このスパイン上に特異的にシナプスを形成し,限局的に興奮性シナプス伝達を行う.
興奮性細胞は,上述の基本的特性を保持しながらも,大脳皮質の領野,層における位置,軸索投射先に依存して,細胞形態,電気生理学的特性,遺伝子発現などにおいて違いを示すことが過去の限られた研究から示唆されてきた.単一細胞トランスクリプトーム解析による最近の体系的研究は,異なる層,領野に存在する,多様な興奮性細胞サブタイプ群の存在を明らかにし1),この見解を強力に支持した.さらに,細胞タイプ特異的遺伝子操作,解剖学的解析,行動中の動物における神経活動記録を組み合わせた研究は,運動野で異なる遺伝子発現を示す二つの異なる5層興奮性細胞サブタイプが,皮質下への異なる軸索投射パターンを示すだけでなく,運動コントロール時に異なるタイミングで発火することを示した2).これらの知見は,興奮性細胞が,遺伝学的,形態学的,機能的に異なるサブタイプを有することを示すとともに,各々のサブタイプは特有の回路メカニズムにより制御されていることを示唆している.実際,視覚野第5層において,視蓋投射錐体細胞は,近隣の視蓋投射錐体細胞からよりも高い確率で脳梁投射錐体細胞から入力を受けることが示されている3).
大脳皮質神経細胞の2割を占め,GABAを神経伝達物質として用いる抑制性細胞は,通常,局所的かつ密なネットワークを後シナプス細胞と形成し,その電気的抑制効果によって,シナプス,細胞,回路レベルで大脳皮質内の神経活動パターン,バランスを制御している.抑制性細胞には,parvalbumin(PV),somatostatin(SOM),vasoactive intestinal peptide(VIP)の発現で分類され,互いにほぼ重複しない三つのサブグループが存在し,それぞれ,形態,電気生理学的特性,結合特異性,機能が異なることが知られている.大部分のPV細胞は,後シナプス細胞の細胞体を神経支配し,細胞全体における電気信号のゲインを決定するのに対し,SOM細胞は,樹状突起を神経支配し,局所的な神経活動制御に関与する.また,VIP細胞は,主に,PV細胞やSOM細胞といった他の抑制性細胞を神経支配し,抑制性細胞が神経支配する興奮性細胞において脱抑制作用を引き起こすことが知られている.
これまで,上述の三つのサブグループは,異なる抑制性細胞サブタイプを代表する単純化されたモデルとして,発達メカニズム,回路機能の解析に利用されてきた.しかしながら,各々のサブグループは,形態,電気生理学的特性,遺伝子発現において多様な細胞タイプを含む,不均一な細胞集団であることが知られている4).最近の研究において,軸索の皮質層分布が異なっているSOM細胞サブタイプは,マウスのヒゲの動きに対して異なる電気活動応答を示すことが明らかにされた5).このことは,抑制性細胞の特異的サブタイプが,神経回路機能において特殊な役割を担っていることを示唆している.発達メカニズム,回路構造,回路機能を特異的サブタイプごとに明らかにしていくことが,今後,ますます重要な課題になってくるものと思われる.
興奮性細胞と抑制性細胞の間で織りなされる大脳皮質神経回路の配線図を明らかにすることは,長年にわたる,神経科学における重要課題の一つである.この問題を解明するために,いくつかのアプローチが用いられてきた.脳スライス内の二つ以上の細胞で同時にパッチクランプを行い,電気生理学的結合性を同定した後,色素注入を行い,細胞形態を可視化する方法は,異なる層に位置する興奮性細胞へ入力する抑制細胞の形態,電気生理学的特性に関して多くの情報をもたらした6, 7).しかしながら,この方法には,一つの興奮性細胞に対して,解析可能な入力細胞の数に限りがあることや,脳スライスを用いるため,一部の神経結合は保全されないという内在的欠点が存在する.前者の問題は,入力細胞にチャネルロドプシンを発現し,皮質を分割する微小領域を順次,隙間なく光刺激し,興奮性細胞から応答を記録することで部分的には解決される8, 9).光遺伝学と組み合わせたこの方法で,一つの興奮性細胞に対する入力細胞の大まかな空間分布が得られるものの,1細胞レベルでの結合性は不明となる.また,スライスを利用することに起因する後者の問題は,解決されないままである.近年,高効率電子顕微鏡法を用いたコネクトミクスは,細胞形態,ならびにシナプスレベルでの結合性を可視化し,大脳皮質の配線図解明に多大な貢献をしている10).しかしながら,この方法は,多大な労力と時間を要すること,また,細胞タイプを定義する遺伝子発現の情報を同時解析するのは困難であるという欠点を持つ.さらに,上述のすべての既存アプローチは,遺伝子発現解析,回路機能解析などへの汎用性が低いという欠点を持つ.
ある特定の神経回路を細胞タイプ特異的に,再現性高く,高効率で可視化する方法は,大脳皮質局所神経回路の発達,構造,機能を解明する上できわめて有用である.また,特定の局所神経回路を構成する細胞において,チャネルロドプシン,GCaMPなどの遺伝学的ツールを発現できるような汎用性の高い方法も求められている.このような遺伝学的アプローチの確立を目指し,最近,我々は,狂犬病ウイルス(rabies virus:RV)による逆行性単一シナプス標識法11)と,2種類の組換え酵素(FlpとCre)の共発現に依存する交差的標識法12)を組み合わせた,intersectional monosynaptic tracing(iMT)法を開発した13)(図1).この遺伝学的アプローチでは,Flpとcyan fluorescent protein(CFP)を有する狂犬病ウイルス(RV-CFP-Flp),抑制性細胞サブクラス特異的Creマウス,Flp/Cre依存的red fluorescent protein(RFP)リポーターマウスが主要な遺伝学的材料となる.まず,TVA(偽型化された狂犬病ウイルス表面のEnvAを認識する受容体),rabies glycoprotein(RG)(狂犬病ウイルスの逆行性経シナプス移動に必要なタンパク質),H2BYFP(一次感染細胞のマーカーとなる核移行型YFP)を目的の興奮性細胞に発現させる.ここでは,上顆粒細胞層と下顆粒細胞層をそれぞれ一次感染細胞にするために,異なる胎生期(胎生15日と12日)で子宮内エレクトロポレーションを行った.次に,EnvAで偽型化され,かつ,RGを欠損したウイルス遺伝子を持つEnvA-RV-ΔRG-CFP-FlpをTVA発現興奮性細胞に一次感染させる.狂犬病ウイルスの特性により,ウイルス粒子が一次感染興奮性細胞からシナプスを介し逆行性に入力細胞へ伝播するが,入力細胞は外来RGを発現していないので,RG欠損型の狂犬病ウイルスはそれ以上伝播することができない.このようにして,一次感染興奮性細胞へ直接入力を送る細胞だけが狂犬病ウイルスを取り込むことになる.この遺伝学的操作を,PV-Cre, Flp/Cre依存的リポーターを有するマウスにおいて行うと,一次感染興奮性細胞に入力を送るPV細胞のみがFlp(狂犬病ウイルス由来)とCre(PV-Cre由来)を共発現し,RFPで標識されることが期待される.実際,2/3層興奮性細胞に入力するPV細胞を検出することを目的にした実験において,RFP標識された入力細胞のPV発現,電気生理学的特性の検証により,iMT法は高い特異性を持つことが実証された.また,iMT法は,2/3層興奮性細胞に入力するSOM細胞,VIP細胞も高い特異性で検出することが示された.ここで注目すべきことは,興奮性細胞に入力を送るVIP細胞はごく少数であるにもかかわらず,iMT法は,このVIP細胞サブタイプを標識することができたということである.この結果は,iMT法が,比較的少数のサブタイプに属する入力細胞も高い効率で検出することができることを示唆している.
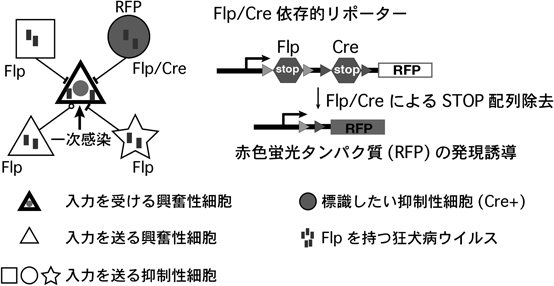
ウイルス由来のFlpと遺伝子改変動物由来のCreが共存することにより,興奮性細胞に入力を送る標的の抑制神経細胞のみで蛍光タンパク質の発現が誘導される.
次に,我々は,iMT法を用い,「異なる興奮性細胞サブタイプは,異なる抑制性入力の回路構造を持つか」という問いに答えるための先駆的実験を行った.その結果,2/3層興奮性細胞は,同層からのみならず深層(4–6層)からも幅広くPV細胞からの入力を受けるのに対し,4–6層興奮性細胞は,主に同層のPV細胞から局所入力を受けることがわかった(図2).また,運動野と前部体性感覚野の2/3層興奮性細胞に入力するSOM細胞の軸索分布を比較したところ,前部体性感覚野1層におけるSOM軸索密度は,運動野に比べて有意に低いことが明らかになった.すべてのSOM細胞を標識した場合,1層の軸索密度は,これらの領野で変わらないことから,1層に軸索投射する特徴的な形態を示し,SOM陽性サブグループに属することで知られるマルチノッチ細胞の2/3層興奮性細胞に対する結合性が,運動野と前部体性感覚野で異なる可能性を示唆している.これらの結果は,層,領野の異なる興奮性細胞は,それぞれ特異的な抑制入力の回路構造を持つという見解を支持している.
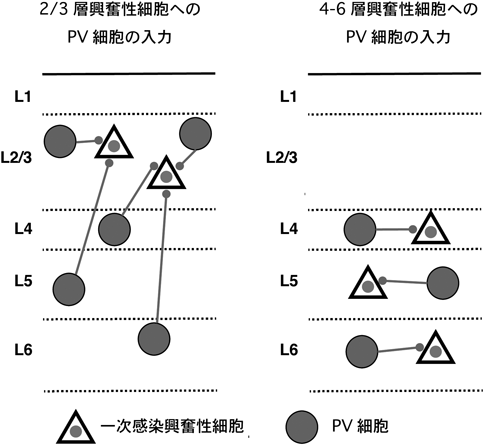
2/3層興奮性細胞は,同層からのみならず深層(4–6層)からも幅広くPV細胞からの入力を受けるのに対し,4–6層興奮性細胞は,主に同層のPV細胞から局所入力を受ける.
iMT法の開発により,特定の興奮性細胞サブタイプへ入力を送る抑制性細胞の大脳皮質内空間分布や軸索投射パターンをサブタイプ特異的に解析することが可能になった.今後,一次感染細胞となる興奮性細胞サブタイプをより限局することにより,異なる抑制性局所神経回路モデュールが発見されることが期待できる.これらの異なる抑制性局所神経回路モデュールを構成する細胞の遺伝子発現を解析することにより,細胞タイプと結合性の関係が明らかになり,より詳細な局所神経回路構造の理解へとつながるものと思われる.また,異なる興奮性細胞サブタイプが,分子,細胞レベルで,どのように特異的な抑制性入力を構築するかといった問題は,まったく未知の課題である.さらに,特異的抑制性入力がどのように興奮性細胞サブタイプの情報処理に寄与しているかを明らかにすることも重要な課題である.
iMT法は,組換え酵素依存的な遺伝子発現を基盤としているため,多様な遺伝学的ツールとの互換性が高く,上記のさまざまな問題を解明する上で有望な方法である.現時点でiMT法の汎用性を阻む大きな原因は,狂犬病ウイルスの毒性である.逆行性経シナプス標識で一般に利用されている狂犬病ウイルスは,感染後,約1週間を過ぎたあたりから毒性を示し始めるため,長期にわたって感染細胞を生理学的条件下で保持することができない.しかしながら,最近,Tripodiらのグループは,狂犬病ウイルス遺伝子にPEST配列を付加し,狂犬病ウイルスを短期間で自己不活化させることにより,その毒性を顕著に低減する方法(self-inactivating RV:SiRV)を開発した14).この効果を否定する報告もなされたが15),我々の研究室では,良好な結果が得られており,今後,iMT法にSiRVを取り入れ,詳細な検討を重ねる予定である.iMT法を改良することにより,近い将来,大脳皮質局所神経回路の多面的研究が躍進することを期待している.
iMT法の確立に向けて多大な尽力をしたMichael Yetman博士をはじめとする共同研究者の方々に謝意を表します.iMT法の確立に関する研究は,JSTさきがけによる支援を受けました.また,iMT法を応用した研究は,National Institute of Mental Health(MH115917)の支援により行われています.
1) Tasic, B., Yao, Z., Graybuck, L.T., Smith, K.A., Nguyen, T.N., Bertagnolli, D., Goldy, J., Garren, E., Economo, M.N., Viswanathan, S., et al. (2018) Shared and distinct transcriptomic cell types across neocortical areas. Nature, 563, 72–78.
2) Economo, M.N., Viswanathan, S., Tasic, B., Bas, E., Winnubst, J., Menon, V., Graybuck, L.T., Nguyen, T.N., Smith, K.A., Yao, Z., et al. (2018) Distinct descending motor cortex pathways and their roles in movement. Nature, 563, 79–84.
3) Brown, S.P. & Hestrin, S. (2009) Intracortical circuits of pyramidal neurons reflect their long-range axonal targets. Nature, 457, 1133–1136.
4) Huang, Z.J. & Paul, A. (2019) The diversity of GABAergic neurons and neural communication elements. Nat. Rev. Neurosci., 20, 563–572.
5) Munoz, W., Tremblay, R., Levenstein, D., & Rudy, B. (2017) Layer-specific modulation of neocortical dendritic inhibition during active wakefulness. Science, 355, 954–959.
6) Kawaguchi, Y. & Kubota, Y. (1996) Physiological and morphological identification of somatostatin- or vasoactive intestinal polypeptide-containing cells among GABAergic cell subtypes in rat frontal cortex. J. Neurosci., 16, 2701–2715.
7) Jiang, X., Shen, S., Cadwell, C.R., Berens, P., Sinz, F., Ecker, A.S., Patel, S., & Tolias, A.S. (2015) Principles of connectivity among morphologically defined cell types in adult neocortex. Science, 350, aac9462.
8) Pluta, S., Naka, A., Veit, J., Telian, G., Yao, L., Hakim, R., Taylor, D., & Adesnik, H. (2015) A direct translaminar inhibitory circuit tunes cortical output. Nat. Neurosci., 18, 1631–1640.
9) Katzel, D., Zemelman, B.V., Buetfering, C., Wolfel, M., & Miesenbock, G. (2011) The columnar and laminar organization of inhibitory connections to neocortical excitatory cells. Nat. Neurosci., 14, 100–107.
10) Motta, A., Berning, M., Boergens, K.M., Staffler, B., Beining, M., Loomba, S., Hennig, P., Wissler, H., & Helmstaedter, M. (2019) Dense connectomic reconstruction in layer 4 of the somatosensory cortex. Science, 366, eaay3134.
11) Wickersham, I.R., Lyon, D.C., Barnard, R.J., Mori, T., Finke, S., Conzelmann, K.K., Young, J.A., & Callaway, E.M. (2007) Monosynaptic restriction of transsynaptic tracing from single, genetically targeted neurons. Neuron, 53, 639–647.
12) Madisen, L., Garner, A.R., Shimaoka, D., Chuong, A.S., Klapoetke, N.C., Li, L., van der Bourg, A., Niino, Y., Egolf, L., Monetti, C., et al. (2015) Transgenic mice for intersectional targeting of neural sensors and effectors with high specificity and performance. Neuron, 85, 942–958.
13) Yetman, M.J., Washburn, E., Hyun, J.H., Osakada, F., Hayano, Y., Zeng, H., Callaway, E.M., Kwon, H.B., & Taniguchi, H. (2019) Intersectional monosynaptic tracing for dissecting subtype-specific organization of GABAergic interneuron inputs. Nat. Neurosci., 22, 492–502.
14) Ciabatti, E., Gonzalez-Rueda, A., Mariotti, L., Morgese, F., & Tripodi, M. (2017) Life-Long Genetic and Functional Access to Neural Circuits Using Self-Inactivating Rabies Virus. Cell, 170, 382–392.
15) Matsuyama, M., Jin, L., Lavin, T.K., Sullivan, H.A., Hou, Y., Lea, N.E., Pruner, M.T., Dam Ferdínez, M.L., & Wickersham, I.R. (2019) “Self-inactivating” rabies viruses are just first-generation, ΔG rabies viruses. bioRxiv, 550640.

マックスプランクフロリダ神経科学研究所リサーチグループリーダー.博士(理学).
大阪大学基礎工学部生物工学科卒.大阪大学大学院医学系研究科医科学修士課程修了.総合研究大学院大学修了.2012年より現職.
研究テーマと抱負大脳神経細胞の「多様性生成」,「回路形成,再編成」の分子メカニズムを理解すること.新しい技術を開発,導入しつつ,未開拓の分野を切り開いていきたい.
ウェブサイトhttps://chandeliercell.jimdo.com/
趣味アメリカで手に入る物を用いての日本食再現.美術,音楽鑑賞.動物園巡り.

This page was created on 2020-03-05T10:34:05.292+09:00
This page was last modified on 2020-03-31T15:09:42.000+09:00
このサイトは(株)国際文献社によって運用されています。