相分離シャペロンとなるインポーチンβファミリーImportinβs function as chaperone for phase-separating protein
立命館大学・生命科学部College of Life Sciences, Ritsumeikan University ◇ 滋賀県草津市野路東1–1–1 ◇ 1–1–1 Noji-higashi, Kusatsu, Shiga Japan
発行日:2020年4月25日Published: April 25, 2020
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
近年,細胞内の相分離が大きな注目を集めている.液-液相分離(liquid-liquid phase separation:LLPS)とも呼ばれるこの現象が,細胞内で重要な役割を担っていることが次々と明らかとなってきた.ここでいう生物学的相分離とは,散らばった分子が集まり,主に液滴が作られることによって分離する現象である.凝集と似ているが,大きな違いはその柔軟性にある.相分離によって形成されたものは流動的であり,さまざまな因子の微妙な変化によって可逆的に変化する.細胞は,相分離による区画化を巧みに使いこなし,高次機能の発現に役立てていると考えられている1).これまでに細胞内顆粒として観察されていた核小体やRNA顆粒などは相分離による産物であり,膜のないオルガネラ(membraneless-organelle)と呼ばれ,あらためて脚光を浴びている.本稿では,現在最も研究が進められているRNA顆粒の形成に関与するFused in Sarcoma(FUS)タンパク質の相分離と,その制御因子である核内輸送受容体インポーチンβファミリーについて紹介する.インポーチンβファミリーは,細胞質中のタンパク質を核内へと輸送するタンパク質であるが,相分離性のタンパク質と結合することで相分離を抑制する「相分離シャペロン」としての機能を持つことが新たに明らかとなった.
近年の相分離ブームの火付け役となった研究対象がRNA結合タンパク質FUSである.FUSは脂肪肉腫関連遺伝子として発見されたが,その後,重篤な神経変性疾患である筋萎縮性側索硬化症(amyotrophic lateral sclerosis:ALS)に関わることが明らかとなった2).FUSは,全長526アミノ酸から構成されるが(図1A),RNA結合モチーフ(RRM)およびZnフィンガーモチーフ(ZnF)を除く,7割以上の領域が単独では特定の構造を持たない天然変性領域からなる.また,その天然変性領域の配列は限られたアミノ酸から構成された低複雑性ドメイン(low-complexity domain:LC)であるという特徴を持つ.N末端領域はセリン,チロシン,グリシン,グルタミン残基が豊富に存在することからSYGQ-rich領域,それ以外の領域にはアルギニン-グリシン-グリシンの繰り返しが多く存在することから,RGG-repeatと呼ばれる.LCは数種類のアミノ酸に富んでることに加え,決まった繰り返しパターンを持つことが多い.SYGQ-rich領域では,チロシン残基の前後にグリシンまたはセリンが並んだ[S/G]Y[S/G]の繰り返しが多くみられる.多くのタンパク質がLCを持つことが認識されていたが,その天然変性領域の機能は謎に包まれてきた.
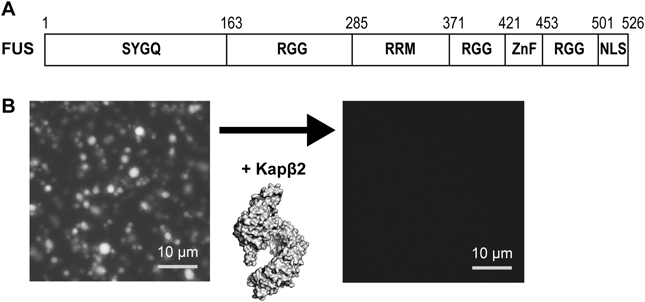
(A)FUSのドメイン構造.SYGQ:SYGQ-rich領域,RGG:RGG-repeat, RRM:RNA結合モチーフ,ZnF:Znフィンガーモチーフ,NLS:核移行シグナル.(B)相分離によるFUSの液滴はKapβ2の添加により消失する.
近年の研究により相分離におけるLCの重要性が明らかとなってきた.FUSは細胞内においてRNA顆粒と呼ばれるRNAとタンパク質からなる構造体に含まれること知られていたが,その機能は明らかとされていなかった.2012年,McKnight研究室のグループがFUSの相分離によるヒドロゲル化がRNA顆粒の形成に重要であることを明らかとした3).特に,N末端領域のSYGQ-rich領域が相分離に重要であることを示し,アミロイド様のクロスβポリマーを形成することが示された.2017年には固体NMR法により立体構造が決定され,SYGQ-rich領域の作るクロスβポリマー構造が原子レベルで明らかとなった4).通常のアミロイド線維よりも疎水的な相互作用が少ないことから,柔軟性を持つことが示唆された.この他,生体高分子の相分離研究のトップを走るHyman研究室やRosen研究室をはじめとするさまざまなグループによりFUSの相分離に関する研究が行われ,FUSは相分離研究のモデルタンパク質となっている5, 6).
FUSはC末端にある核移行シグナル(nuclear localization signal:NLS)によって,主に核に局在する.FUSのNLSはプロリン-チロシン残基を持つPY-NLSに分類されるものであり,Karyopherin β2(Kapβ2)によって認識される.Kapβ2は核内輸送受容体であるインポーチンβファミリーの一つであり,PY-NLSを持つタンパク質を核内へと輸送することが知られている.ヒトではおよそ20種類のインポーチンβファミリーが存在し,それぞれ特有のNLSを認識し,核内へと輸送基質を運搬する働きを持つ7).
核-細胞質間の生体高分子のやり取りは核膜上に存在する核膜孔複合体(nuclear pore complex:NPC)を通して行われる.円筒状のNPC内部は核膜孔タンパク質(nucleoporin:Nup)のフェニルアラニン-グリシン配列に富んだFG-repeatで満たされており,40 kDa以上の生体高分子は自由に行き来ができない.FG-repeatもLCの一種であり,相分離による分子バリアを作ることで,不要な生体高分子の出入りを防ぐと考えられている8).インポーチンβファミリーはNPCを通過できる性質を持っており,インポーチンβファミリーと結合した生体高分子が選択的にNPCを通過できる仕組みとなっている.
FUSのALSに関連するアミノ酸変異を伴う遺伝子変異はPY-NLS領域に集中している.病態として,細胞質でのFUSの沈着がみられることから,Kapβ2によるFUSの核輸送の破綻と疾患との関連が示唆される.
FUSは高い自己会合性を示すことから,その自己会合性による凝集が疾患を招くと考えられ研究されてきた.分散状態と凝集状態の中間ともいえる相分離状態が明らかになったことで,FUSの新たな側面がみえてきた.FUSは細胞質ではストレス顆粒と呼ばれるRNA顆粒の一種の主要なコンポーネントとなることが知られている.ストレス顆粒は,ストレス時に一過的に形成され,mRNAを蓄え翻訳を抑制することで細胞を守ると考えられている.つまり,FUSの自己会合による相分離は細胞の防御応答に役立っており,ストレス顆粒がうまく解消されないことにより,異常な凝集体へと状態変化することが問題である考えられるようになってきた.疾患に関連するFUSのアミノ酸変異が,Kapβ2との相互作用に関わるPY-NLSに集中していることから,我々は,FUSを細胞質から核内へと輸送するKapβ2がFUSの相分離を制御する鍵となると考えた.まず,相分離状態のFUSに対してどのように働くかについて調べた.精製したタンパク質を用い,テストチューブ内でのFUSの液滴解析を行った.FUSの液滴を形成させてから,Kapβ2を加えたところ,速やかにFUSの液滴は消失した(図1B).Kapβ2以外のインポーチンβであるインポーチンα/β1やKap121では,部分的に抑制するもののKapβ2ほどの抑制能は持たなかった.しかし,FUSのNLSをインポーチンα/β1とKap121がそれぞれ認識する特有のNLS配列に置き換えたところ,高い相分離抑制能を示した.その他の検証実験の結果からも,相分離抑制にはNLSを介した強い相互作用が重要であることが示された9).
Kapβ2がFUSの相分離を抑制するためにはPY-NLSとの相互作用が重要であることが明らかとなったが,FUSのPY-NLSはC末端のわずか20アミノ酸程度の領域であり,相分離を促進するSYGQ-rich領域やRGG-repeatは残りの500残基の広域にわたって存在する.Kapβ2がFUSの相分離を抑制するためにPY-NLSとの結合だけで十分であるか,疑問が残った.そこでPY-NLSを持たないFUSに対して,Kapβ2は相分離抑制を行えるか検証を行った.その結果,PY-NLSを介して結合できない状態であったとしても過剰量のKapβ2を加えることでFUSの相分離を抑制する効果がみられた.
Kapβ2とFUSのPY-NLS以外の領域との相互作用について,さらに解析を進めた.Kapβ2とFUSとの複合体でのX線結晶構造解析を試みたところ,複合体での結晶化およびデータ収集には成功した.しかし,Kapβ2については全長を電子密度にあてはめモデリングできたのに対し,FUSにおいて電子密度が確認できたのはPY-NLS領域のみであった.したがって,X線結晶構造解析の結果からは,Kapβ2とFUSのPY-NLS以外の領域との相互作用は,弱いものであるか,過渡的であることが示唆された.そこで,弱い相互作用を検出するため,NMRによる解析を行った.同位体標識したFUSに対して,Kapβ2を加えた結果,複数のシグナルの摂動がみられた.Kapβ2が相互作用するFUSの領域は広域にわたっており,SYGQ-rich領域やRGG-repeatといったLCを含むさまざまな領域と相互作用することが明らかとなった.以上の結果を踏まえ,Kapβ2はPY-NLSを介した強い相互作用によって,その他の相分離に関与する領域との相互作用が可能となり,相分離を抑制するシャペロンとしての機能を果たすモデルを提唱した.
我々の研究グループの他に,3グループが同時に同様の研究成果を発表した10–12).我々がKapβ2とFUSとの相互作用にフォーカスして,物理化学的な手法による相互作用解析を行ったのに対し,他のグループはFUS以外のRNA結合タンパク質に対する相分離抑制能の検証,酵母やショウジョウバエを用いた細胞内での機能解析や,FUSのRGG-repeatのアルギニンのメチル化といった翻訳後修飾の影響についての結果を報告した.また,興味深いことは,このKapβ2の相分離シャペロンとしての機能がNPCを通過する際にも重要に思える点である.これまで,インポーチンβファミリーが選択的にNPCを通過するメカニズムは謎であったが,インポーチンβの相分離シャペロンとしての機能によりLCであるFG-repeatで満たされた核膜孔内部を通過することができるようになると考えられる13).インポーチンβファミリーは,これまで輸送タンパク質としての役割のみが知られてきたが,LCをほどく機能が重要な機能であることが明らかとなりつつある(図2).相分離を起こす研究は盛んに行われているが,それらを制御する因子についてはいまだ明らかにされていない点が多いため,今後の研究が期待される.
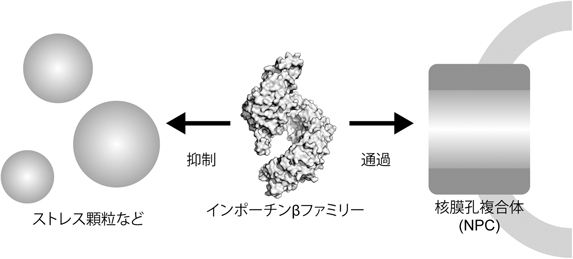
インポーチンβファミリーは相分離状態を解消させることで機能すると考えられる.
これらの研究は米国テキサス大学サウスウエスタンメディカルデンターのYuh Min Chook教授の研究室で行われたものです.共著者含め,ご協力いただいたすべての皆様に感謝申し上げます.
1) Banani, S.F., Lee, H.O., Hyman, A.A., & Rosen, M.K. (2017) Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol., 18, 285–298.
2) Vance, C., Scotter, E.L., Nishimura, A.L., Troakes, C., Mitchell, J.C., Kathe, C., Urwin, H., Manser, C., Miller, C.C., Hortobágyi, T., et al. (2013) ALS mutant FUS disrupts nuclear localization and sequesters wild-type FUS within cytoplasmic stress granules. Hum. Mol. Genet., 22, 2676–2688.
3) Kato, M., Han, T.W., Xie, S., Shi, K., Du, X., Wu, L.C., Mirzaei, H., Goldsmith, E.J., Longgood, J., Pei, J., et al. (2012) Cell-free formation of RNA granules: Low complexity sequence domains form dynamic fibers within hydrogels. Cell, 149, 753–767.
4) Murray, D.T., Kato, M., Lin, Y., Thurber, K.R., Hung, I., Mcknight, S.L., Tycko, R., Murray, D.T., Kato, M., Lin, Y., et al. (2017) Structure of FUS Protein Fibrils and Its Relevance to Self-Assembly and Phase Separation of Low- Article Structure of FUS Protein Fibrils and Its Relevance to Self-Assembly and Phase Separation of Low-Complexity Domains. Cell, 171, 1–13.
5) Lin, Y., Protter, D.S.W., Rosen, M.K., & Parker, R. (2015) Formation and Maturation of Phase-Separated Liquid Droplets by RNA-Binding Proteins. Mol. Cell, 60, 208–219.
6) Wang, J., Choi, J.M., Holehouse, A.S., Lee, H.O., Zhang, X., Jahnel, M., Maharana, S., Lemaitre, R., Pozniakovsky, A., Drechsel, D., et al. (2018) A Molecular Grammar Governing the Driving Forces for Phase Separation of Prion-like RNA Binding Proteins. Cell, 174, 688–699.e16.
7) Xu, D., Farmer, A., & Chook, Y.M. (2010) Recognition of nuclear targeting signals by Karyopherin-β proteins. Curr. Opin. Struct. Biol., 20, 782–790.
8) Hülsmann, B.B., Labokha, A.A., & Görlich, D. (2012) The permeability of reconstituted nuclear pores provides direct evidence for the selective phase model. Cell, 150, 738–751.
9) Yoshizawa, T., Ali, R., Jiou, J., Fung, H.Y.J., Burke, K.A., Kim, S.J., Lin, Y., Peeples, W.B., Saltzberg, D., Soniat, M., et al. (2018) Nuclear Import Receptor Inhibits Phase Separation of FUS through Binding to Multiple Sites. Cell, 173, 693–705.
10) Guo, L., Kim, H.J., Wang, H., Monaghan, J., Freyermuth, F., Sung, J.C., O’Donovan, K., Fare, C.M., Diaz, Z., Singh, N., et al. (2018) Nuclear-Import Receptors Reverse Aberrant Phase Transitions of RNA-Binding Proteins with Prion-like Domains. Cell, 173, 677–692.e20.
11) Hofweber, M., Hutten, S., Bourgeois, B., Spreitzer, E., Niedner-Boblenz, A., Schifferer, M., Ruepp, M.D., Simons, M., Niessing, D., Madl, T., et al. (2018) Phase Separation of FUS Is Suppressed by Its Nuclear Import Receptor and Arginine Methylation. Cell, 173, 706–713.e13.
12) Qamar, S., Wang, G.Z., Randle, S.J., Ruggeri, F.S., Varela, J.A., Lin, J.Q., Phillips, E.C., Miyashita, A., Williams, D., Ströhl, F., et al. (2018) FUS Phase Separation Is Modulated by a Molecular Chaperone and Methylation of Arginine Cation-π Interactions. Cell, 173, 720–734.
13) Schmidt, H.B. & Görlich, D. (2015) Nup98 FG domains from diverse species spontaneously phase-separate into particles with nuclear pore-like permselectivity. eLife, 4, 1–30.

立命館大学生命科学部助教.博士(理学).
1985年神奈川県に生る.2012年横浜市立大学大学院生命ナノシステム科学研究科博士後期課程修了.テキサス大学サウスウエスタンメディカルセンターポスドクを経て16年より現職.
研究テーマと抱負相分離性タンパク質と制御因子の構造機能解析.相分離における分子構造を解明することを目指している.
ウェブサイトhttp://www.ritsumei.ac.jp/lifescience/skbiot/matsumura/index.html(研究室ホームページ)
趣味ランニング,自転車.

This page was created on 2020-03-10T13:22:39.032+09:00
This page was last modified on 2020-03-31T17:39:18.000+09:00
このサイトは(株)国際文献社によって運用されています。