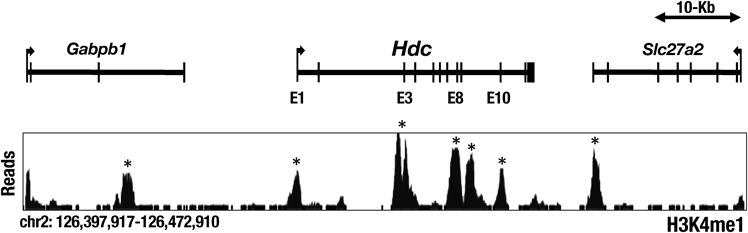
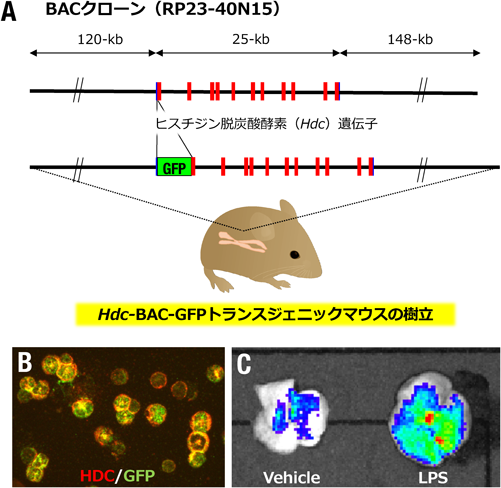
敗血症性肺傷害時の肺に集積するヒスタミン産生能を持つ好中球の役割The role of histamine-producing neutrophils that accumulate in septic lung
東北医科薬科大学医学部医化学教室Division of Medical Biochemistry, Tohoku Medical and Pharmaceutical University ◇ 〒983–8536 仙台市宮城野区福室1–15–1 ◇ 1–15–1 Fukumuro, Miyagino-ku, Sendai 983–8536, Japan
受付日:2020年1月24日Received: January 24, 2020
発行日:2020年4月25日Published: April 25, 2020