健康長寿とグライコミクスGlycomics for healthy aging
東京都健康長寿医療センター研究所老化機構研究チーム・プロテオームTokyo Metropolitan Institute of Gerontology ◇ 〒173–0015 東京都板橋区栄町35–2 ◇ 35–2 Sakae-cho, Itabashi-ku, Tokyo 173–0015, Japan
発行日:2020年6月25日Published: June 25, 2020
糖鎖を網羅的に解析するグライコミクスは,近年の分析技術の進歩により飛躍的な発展を遂げ,N結合型糖鎖プロファイル(N-グライコーム)がさまざまな健康状態の変化を反映して変動することがわかってきた.そこで筆者らは「健康長寿」に特徴的なN-グライコームがあると仮定し,N-グライコミクスにより健康長寿のマーカー探索や長寿のメカニズム解明に迫れるのではないか,と考えている.そして,健康長寿を達成したと考えられる超百寿者(105歳以上)に特徴的な血しょう糖鎖を調べ,健康長寿との関連について解析している.本稿では,血しょうおよび血清N-グライコームとその変動因子について概観した後,筆者らの最近の研究を紹介する.そして,糖鎖の網羅的な解析により何を明らかにできるか議論したい.
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
ヒト血しょうに含まれるタンパク質は,アルブミンを除くほとんどすべてが糖タンパク質である1).これらの糖タンパク質のアスパラギン残基に結合した糖鎖(N結合型糖鎖)を網羅的に調べるN-グライコミクスは,近年の質量分析装置の高感度化,高精度化により,技術面で飛躍的な進歩を遂げてきた.血しょうN-グライコミクスによって明らかになるN結合型糖鎖プロファイル(血しょうN-グライコーム)は,疾患だけでなく生活習慣や加齢などのさまざまな健康状態の変化を反映することが示唆されている2, 3).そのため,健康診断への応用やさまざまなバイオマーカーとしての利用など,健康な超高齢社会の実現に向けた新しいツールになるものと期待されている.
ところで,日本は世界でも例をみない速度で高齢化が進行し,2019年にはついに100歳以上の人口が7万人を超えるまでに至った.そして「老後破産」や「下流老人」といった言葉に注目が集まり,社会保障や福祉制度のありかたへの関心が高まっている昨今,この超高齢社会をいかに維持していくかは,世界が注目するところとなっている.この困難な課題の解決策の一つと考えられるのは「健康寿命を延ばすこと」である.多くの高齢者が肉体的にも精神的にも健康を維持し,productive agingを実現することは,超高齢社会を健全に保つための喫緊の課題である.「健康でいられる時間をできるだけ長くしたい」というのは,今や個人の願いであるだけでなく,社会全体の課題となっている.
そこで本稿では,血しょうN-グライコミクスを「健康長寿の研究」に役立てようと取り組んでいる筆者たちの研究について紹介したい.健康長寿を達成したと考えられる日本人超百寿者(105歳以上)の血しょうN-グライコームを解析し,超百寿者に特徴的な糖鎖ならびにそのキャリアタンパク質を明らかにすることから,健康長寿のメカニズムの解明に向けアプローチしている.
まず初めに,健康状態が安定している場合の血しょうN-グライコームについて紹介する.Gornikらは,12人の健康な人から数日間にわたって7回採血を行い,個人ごとに血しょうN-グライコームのばらつきを調べた.その結果,12人の変動係数(coefficient of variation:CV)は平均して5.6%であり,健康状態に変化のない期間においては血しょうN-グライコームは安定していると報告した4).また,5人の健康なボランティアの血しょうN-グライコームを1.5年から6年にわたって縦断的に調べたHennigらも,季節の変化などには影響を受けず,生活習慣等の変化がない限り個人のN-グライコームはほとんど変化しないことを報告している5).これらの報告から,生活習慣や健康状態に変化がなければ,血しょうN-グライコームに変動はないものと考えられる.
しかし,ひとたび健康状態が変化すると,血しょうおよび血清N-グライコームのパターンは変化する.疾患による糖鎖変化についてはこれまでにもかなり報告があるが,最もよく研究されているのは炎症性疾患とがんである.
炎症性疾患に関しては,クローン病や潰瘍性大腸炎などの炎症性腸疾患6),血流感染症7),慢性腰痛8),慢性膵炎9),リウマチ10)などで調べられている.これら炎症性疾患での糖鎖変化としては,三分岐・四分岐の高分岐糖鎖の増加やシアル酸含有糖鎖の増加,また高マンノース型糖鎖の減少などの変化があげられている.これらの報告の多くは,N結合型糖鎖をタンパク質から切り離して解析したものである.一方,糖ペプチド解析を行ったり,二次元電気泳動を利用したりすることで糖タンパク質として解析している報告もある.それらの報告では,血流感染症や慢性膵炎により,免疫グロブリンG(IgG),免疫グロブリンAの他,α1-酸性糖タンパク質,ハプトグロビン,フェツイン,トランスフェリンといった急性期タンパク質の糖鎖が変化している7, 9).炎症の急性期に血液中に増加する急性期タンパク質が,タンパク質の発現量増加だけでなく,糖鎖の変化も伴うことは興味深い.
また,がんでは膵がん9),直腸がん11, 12),膀胱がん13)などで解析されており,コアフコシル化糖鎖(膵がん),β1,6-分岐糖鎖,α2,6結合のシアル酸含有糖鎖などの増加が報告されている.
さらに,糖尿病でも血しょうおよび血清N-グライコームが変化する14).糖尿病にはI型とII型があるが,両方に共通した変化として高分岐糖鎖が増加することが報告されている14).また,II型糖尿病ではα2,6結合のシアル酸含有糖鎖が増加し,α2,3結合のシアル酸含有糖鎖が減少する15).ゲノムワイド関連解析(GWAS)によって,シアル酸転移酵素遺伝子(ST6GAL1)がII型糖尿病の発症リスクアレル候補としてあげられており16),II型糖尿病におけるこれらの変化はI型とII型を区別できる特徴かもしれない.
その他にもダウン症17)や無症候性脳梗塞18)における血しょうおよび血清N-グライコーム変化が調べられている.ダウン症では,健常者に比べてガラクトース含有糖鎖やα2,3結合のシアル酸含有糖鎖が減少し,コアフコース含有糖鎖が増加した17).また,無症候性脳梗塞では二分岐・コアフコース含有糖鎖が減少し,三分岐・α1,3-フコース含有糖鎖が増加した18).これらの糖鎖変化は,診断のバイオマーカーとなるかもしれない.
疾患に伴う健康状態の変化でなくても,N-グライコームは,飲酒や喫煙などの生活習慣や加齢により変化することが知られている3, 19–21).特に,IgGの糖鎖の加齢変化については,暦年齢だけでなく,生物学的年齢とも相関することが報告されている22).Kristicらは四つのコホートの18歳から98歳まで約5000人のIgGの糖鎖を調べ,加齢によって増加する1種類の糖鎖と減少する2種類の糖鎖のピーク面積から「糖鎖年齢」(GlycanAge)と定義したパラメーターを算出した.このGlycanAgeは暦年齢とよい相関を示したが(相関係数は0.64~0.70),年齢と相関する生理学的なパラメーター[肺活量,収縮期血圧,body mass index(BMI)など]ともよい相関を示し,糖鎖が生物学的年齢のバイオマーカーとなることを示唆した22).そこで,IgG以外のタンパク質に結合する糖鎖も生物学的年齢を反映すると仮定すると,健康長寿を達成した人の血しょうN-グライコームを調べることで,暦年齢を超えた健康長寿の共通メカニズムに迫ることが可能かもしれない.
まず,健康長寿にとって重要な因子と考えられるInflammagingについて紹介する.Franceshiは免疫老化の研究から,加齢により亢進する慢性炎症が高齢者の身体機能の低下や死亡につながることを明らかにし,Inflammagingの概念を提唱した23, 24).慢性炎症の亢進には,老化細胞が分泌するsenescence-associated secretory phenotype(SASP)の関与が指摘されている25, 26).SASPとは,老化細胞が分泌するさまざまな因子であり,炎症およびがんの発生や進展を促進する作用を持つ.具体的には,炎症性サイトカイン,ケモカイン,細胞外マトリクス分解酵素,ロイコトリエンなどが含まれる25–27).
高齢者においては慢性炎症が亢進するため,図1Aに示すように炎症性サイトカイン(IL-6, TNF-αなど),プロスタグランジン,ロイコトリエンなどを中心とする炎症性ネットワークシステムと,抗炎症性サイトカイン(IL-10, TGF-β)やコルチゾルなどを中心とする抗炎症性ネットワークシステムとのバランスが崩れることが,Inflammagingをもたらすと考えられている24).
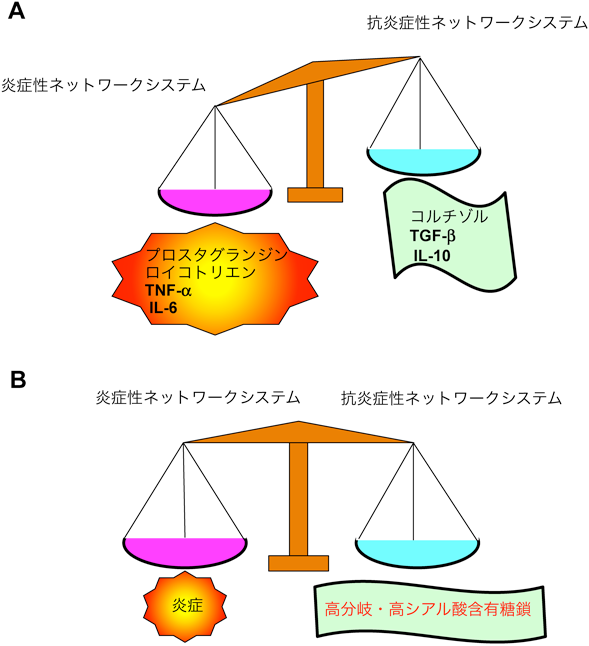
(A)一般高齢者 文献24より改変引用,(B)超百寿者.
日本人の高齢者においても,Inflammagingは余命を規定する重要な因子であるという.新井らは,85~99歳,100~104歳,105歳以上の三つの年齢群において,造血能,肝機能,腎機能,脂質・糖代謝,白血球テロメア長,炎症指標(サイトメガロウイルス抗体価,CRP, IL-6, TNF-αの4因子から求めたZスコア)といった老化関連バイオマーカーと余命との関連を調べた.その結果,炎症指標がすべての年齢群において余命を規定する唯一の要因であった28).さらに,炎症指標はすべての年齢群で,日常生活動作(activities of daily living:ADL)や認知機能とも有意な関連を示した28).つまり,高齢者にとってInflammagingを抑えることは,健康長寿にとってきわめて重要であると考えられる.
橋本らは最近,110歳以上の超百寿者の血液中に,特殊なT細胞であるCD4陽性キラーT細胞が多いことを報告した29).健康長寿にはInflammagingを抑えるだけでなく,免疫応答全体を維持することが重要なのであろう.今後の展開を期待したい.
筆者らは,治療中の疾患を有さない,すなわち健康な超百寿者(平均106.7歳)の血しょうN-グライコミクスを行い,健康長寿に特徴的なN結合型糖鎖を調べた.血しょう糖タンパク質からペプチド:N-グリコシダーゼF(PNGase F)を用いて,N結合型糖鎖を切り出し,液体クロマトグラフィー質量分析法(LC-MS/MS)を用いて分析したところ,1人あたり50種類の糖鎖が同定できた.それらについて,相対ピーク面積比を算出し,多変量解析を行って若齢(平均30.2歳),老齢(平均71.6歳)のN-グライコームと比較した.その結果,三分岐・四分岐(高分岐)でシアル酸を含む糖鎖が超百寿者に多く,逆に二分岐糖鎖が超百寿者に少ないことが明らかになった(図2)30).
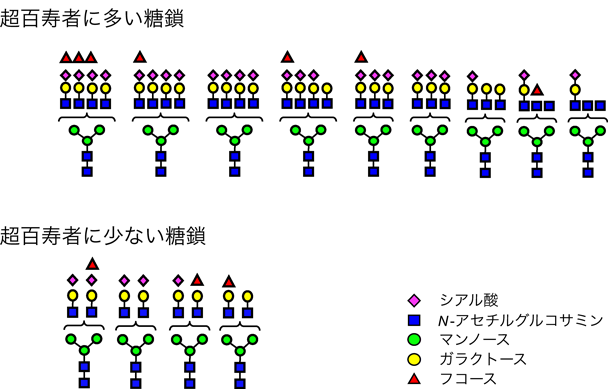
文献30より改変引用.
高分岐でシアル酸を多く含む糖鎖は,上述したように炎症に伴って増加する.そこで,超百寿者の血液中の炎症マーカーやSASPとしても知られる炎症性サイトカイン(IL-6やTNF-α)を測定した.その結果,若齢や老齢の対照群より超百寿者で,これらの炎症マーカーが増加していることが明らかになった30).この結果は,健康な超百寿者も,加齢によって亢進するInflammagingから逃れているわけではないことを示している.そこで筆者らは,健康な超百寿者では,高分岐・高シアル酸含有糖鎖の増加を特徴とするN-グライコームの変化によって,Inflammagingが亢進しても“炎症バランス”が崩れず安定に保たれるのではないかと考えた(図1B).
しかし,「超百寿者」に特徴的な糖鎖が「健康長寿」に特徴的な糖鎖とはいい切れず,その生物学的な役割や意義は慎重に調べる必要がある.そこで次に,糖鎖が結合しているキャリアタンパク質に着目しさらに解析を進めた.
キャリアタンパク質を同定するためには,糖鎖をタンパク質から切り離すことなく,糖ペプチドとして解析するN-グライコプロテオミクスを行う必要がある.
そこで,複合型のN結合型糖鎖を含む糖タンパク質をレクチン(ECA, Erythrina cristagalli)を用いて濃縮後,酵素消化し,LC-MS/MSを用いて若齢(21~38歳),老齢(71~74歳),超百寿(106~107歳)のN-グライコプロテオミクスを行った.それぞれのタンパク質量で補正した糖ペプチドのピーク面積比について多変量解析を行ったところ,ハプトグロビンの207Asnと211Asnを含むペプチド(NLFL207NHSE211NATAK)に結合した高分岐・高シアル酸含有糖鎖が,超百寿者に多いことが明らかになった31).次に,207Nと211N,それぞれの部位特異的な糖鎖修飾を詳細に調べるため,グルタミルエンドペプチダーゼ(Glu-C)を利用して207Asnを含むペプチド(NLFL207NHSE)と211Asnを含むペプチド(211NATAK)に消化し,207と211それぞれに結合する糖鎖を調べた.その結果,超百寿者に特徴的なハプトグロビンの部位特異的な糖鎖の組合わせは図3に示すものであった31).以上より,超百寿者に特徴的な糖鎖のキャリアタンパク質の一つは,ハプトグロビンであることが明らかになった.
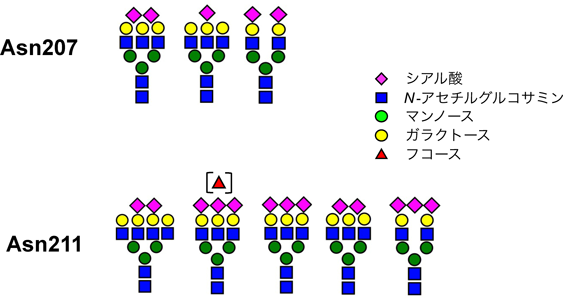
文献31より改変引用.
ハプトグロビンは,炎症によって増加する急性期タンパク質で,ヘモグロビンと結合し,肝臓でのヘモグロビンの分解に関与することが知られている.また,ハプトグロビンの糖鎖はさまざまな疾患バイオマーカーとなっており,膵がん,肝がん,肝硬変などにより,フコースやシアル酸が増加することが知られている32–34).そこで担がんや肝疾患の影響を反映したものであるか,その関連性を探った.超百寿者の肝機能や病歴について調べたところ,いずれも肝機能は正常で肝がんなどの病歴もなく,ここで観察された超百寿者の特徴的なハプトグロビンの糖鎖変化は,これらの罹患によるものではないと推測された31).
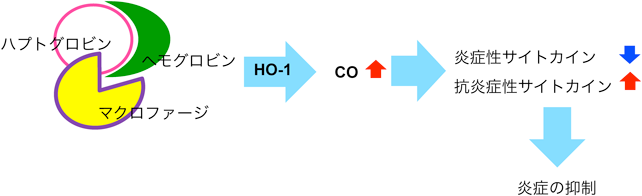
一方,ハプトグロビンは,抗炎症性シグナルにも関与していることが知られている(図4).すなわち,ハプトグロビンはヘモグロビンと結合した後,マクロファージ表面のCD163と結合し,マクロファージに取り込まれる.マクロファージ内では,ヘムオキシゲナーゼ1(HO-1)によってヘモグロビンが分解されるが,その際に発生する一酸化炭素(CO)は,炎症性サイトカイン(IL-1βやTNF-αなど)の産生を抑制し,抗炎症性サイトカイン(IL-10など)の産生を促進することによって,炎症を抑制する35, 36).つまり,炎症の急性期に増加するハプトグロビンは,このような抗炎症性シグナルによって,炎症に傾いたバランスを元に戻す働きがあると考えられる.筆者らは,血しょう中のハプトグロビンが,超百寿者群では若齢対照群に比べて多いことを報告しているが37),もし超百寿者に特徴的なハプトグロビンの糖鎖が,何らかのメカニズムでこの抗炎症性シグナルを促進するなら,キャリアタンパク質とこれら特徴的な糖鎖の両方の増加によって効果的にInflammagingが抑制され,健康長寿がもたらされるのかもしれない.
糖鎖の網羅的な解析法であるグライコミクスは,健康状態を反映するグライコームの変化をハイスループットに検出し,バイオマーカーとなる糖鎖変化を見いだす手法として,非常に有用である.近年,シアル酸の結合様式特異的な化学誘導体化法を利用し,質量分析法によりα2,3結合とα2,6結合を容易に区別できるようになっており38),より詳細な網羅的グライコミクスと新たなバイオマーカー開発への展開が期待される.しかし,グライコーム変化の生物学的な意味について考察することは難しく,キャリアタンパク質の情報を同時に取得することが重要である.今後,シアル酸結合様式特異的な化学誘導体化法などの前処理法を含めた高感度・高精度な分析手法の開発によってグライコプロテオミクスのハイスループット化が進み,さらにグライコプロテオミクスとゲノミクスやプロテオミクスなど他のオミクス情報とを統合させる,いわゆるBig Data Scienceへ展開することにより,健康長寿に関わる糖鎖変化とそのメカニズムの全貌が明らかにされるだろう.健康な超高齢社会が糖鎖バイオマーカーによってもたらされる日が一日も早く来ることを期待したい.
本稿で紹介した研究は,共同研究者である橋井則貴博士(国立医薬品食品衛生研究所),新井康通博士,広瀬信義博士(慶應義塾大学百寿総合研究センター)の協力のもとに行ったものであり,深く感謝いたします.また,本研究成果の一部は日本学術振興会科学研究費補助金による支援を受けたものです.ここに謝意を表します.
1) Clerc, F., Reiding, K.R., Jansen, B.C., Kammeijer, G.S., Bondt, A., & Wuhrer, M. (2016) Human plasma protein N-glycosylation. Glycoconj. J., 33, 309–343.
2) Dotz, V. & Wuhrer, M. (2019) N-glycome signatures in human plasma: associations with physiology and major diseases. FEBS Lett., 593, 2966–2976.
3) Miura, Y. & Endo, T. (2016) Glycomics and glycoproteomics focused on aging and age-related diseases—Glycans as a potential biomarker for physiological alterations. Biochim. Biophys. Acta, 1860, 1608–1614.
4) Gornik, O., Wagner, J., Pucic, M., Knezevic, A., Redzic, I., & Lauc, G. (2009) Stability of N-glycan profiles in human plasma. Glycobiology, 19, 1547–1553.
5) Hennig, R., Cajic, S., Borowiak, M., Hoffmann, M., Kottler, R., Reichl, U., & Rapp, E. (2016) Towards personalized diagnostics via longitudinal study of the human plasma N-glycome. Biochim. Biophys. Acta, 1860, 1728–1738.
6) Clerc, F., Novokmet, M., Dotz, V., Reiding, K.R., de Haan, N., Kammeijer, G.S.M., Dalebout, H., Bladergroen, M.R., Vukovic, F., Rapp, E., et al.; IBD-BIOM Consortium (2018) Plasma N-glycan signatures are associated with features of inflammatory bowel diseases. Gastroenterology, 155, 829–843.
7) Joenvaara, S., Saraswat, M., Kuusela, P., Saraswat, S., Agarwal, R., Kaartinen, J., Jarvinen, A., & Renkonen, R. (2018) Quantitative N-glycoproteomics reveals altered glycosylation levels of various plasma proteins in bloodstream infected patients. PLoS One, 13, e0195006.
8) Trbojevic-Akmacic, I., Vuckovic, F., Vilaj, M., Skelin, A., Karssen, L.C., Kristic, J., Juric, J., Momcilovic, A., Simunovic, J., Mangino, M., et al. (2018) Plasma N-glycome composition associates with chronic low back pain. Biochim. Biophys. Acta, 1862, 2124–2133.
9) Sarrats, A., Saldova, R., Pla, E., Fort, E., Harvey, D.J., Struwe, W.B., de Llorens, R., Rudd, P.M., & Peracaula, R. (2010) Glycosylation of liver acute-phase proteins in pancreatic cancer and chronic pancreatitis. Proteomics Clin. Appl., 4, 432–448.
10) Reiding, K.R., Bondt, A., Hennig, R., Gardner, R.A., O’Flaherty, R., Trbojevic-Akmacic, I., Shubhakar, A., Hazes, J.M.W., Reichl, U., Fernandes, D.L., et al. (2019) High-throughput serum N-glycomics: Method comparison and application to study rheumatoid arthritis and pregnancy-associated changes. Mol. Cell. Proteomics, 18, 3–15.
11) de Vroome, S.W., Holst, S., Girondo, M.R., van der Burgt, Y.E.M., Mesker, W.E., Tollenaar, R., & Wuhrer, M. (2018) Serum N-glycome alterations in colorectal cancer associate with survival. Oncotarget, 9, 30610–30623.
12) Doherty, M., Theodoratou, E., Walsh, I., Adamczyk, B., Stockmann, H., Agakov, F., Timofeeva, M., Trbojevic-Akmacic, I., Vuckovic, F., Duffy, F., et al. (2018) Plasma N-glycans in colorectal cancer risk. Sci. Rep., 8, 8655.
13) Ferdosi, S., Ho, T.H., Castle, E.P., Stanton, M.L., & Borges, C.R. (2018) Behavior of blood plasma glycan features in bladder cancer. PLoS One, 13, e0201208.
14) Rudman, N., Gornik, O., & Lauc, G. (2019) Altered N-glycosylation profiles as potential biomarkers and drug targets in diabetes. FEBS Lett., 593, 1598–1615.
15) Dotz, V., Lemmers, R.F.H., Reiding, K.R., Hipgrave Ederveen, A.L., Lieverse, A.G., Mulder, M.T., Sijbrands, E.J.G., Wuhrer, M., & van Hoek, M. (2018) Plasma protein N-glycan signatures of type 2 diabetes. Biochim. Biophys. Acta, 1862, 2613–2622.
16) Mahajan, A., Taliun, D., Thurner, M., Robertson, N.R., Torres, J.M., Rayner, N.W., Payne, A.J., Steinthorsdottir, V., Scott, R.A., Grarup, N., et al. (2018) Fine-mapping type 2 diabetes loci to single-variant resolution using high-density imputation and islet-specific epigenome maps. Nat. Genet., 50, 1505–1513.
17) Borelli, V., Vanhooren, V., Lonardi, E., Reiding, K.R., Capri, M., Libert, C., Garagnani, P., Salvioli, S., Franceschi, C., & Wuhrer, M. (2015) Plasma N-glycome signature of Down Syndrome. J. Proteome Res., 14, 4232–4245.
18) Vilar-Bergua, A., Riba-Llena, I., Vanhooren, V., Dewaele, S., Libert, C., Penalba, A., Montaner, J., & Delgado, P. (2015) N-glycome profile levels relate to silent brain infarcts in a cohort of hypertensives. J. Am. Heart Assoc., 4, e002669.
19) Dall’Olio, F., Vanhooren, V., Chen, C.C., Slagboom, P.E., Wuhrer, M., & Franceschi, C. (2013) N-glycomic biomarkers of biological aging and longevity: A link with inflammaging. Ageing Res. Rev., 12, 685–698.
20) Ding, N., Nie, H., Sun, X., Sun, W., Qu, Y., Liu, X., Yao, Y., Liang, X., Chen, C.C., & Li, Y. (2011) Human serum N-glycan profiles are age and sex dependent. Age Ageing, 40, 568–575.
21) Knezevic, A., Polasek, O., Gornik, O., Rudan, I., Campbell, H., Hayward, C., Wright, A., Kolcic, I., O’Donoghue, N., Bones, J., et al. (2009) Variability, heritability and environmental determinants of human plasma N-glycome. J. Proteome Res., 8, 694–701.
22) Kristic, J., Vuckovic, F., Menni, C., Klaric, L., Keser, T., Beceheli, I., Pucic-Bakovic, M., Novokmet, M., Mangino, M., Thaqi, K., et al. (2014) Glycans are a novel biomarker of chronological and biological ages. J. Gerontol. A Biol. Sci. Med. Sci., 69, 779–789.
23) Franceschi, C., Bonafe, M., Valensin, S., Olivieri, F., De Luca, M., Ottaviani, E., & De Benedictis, G. (2000) Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci., 908, 244–254.
24) Franceschi, C., Capri, M., Monti, D., Giunta, S., Olivieri, F., Sevini, F., Panourgia, M.P., Invidia, L., Celani, L., Scurti, M., et al. (2007) Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev., 128, 92–105.
25) Freund, A., Orjalo, A.V., Desprez, P.Y., & Campisi, J. (2010) Inflammatory networks during cellular senescence: Causes and consequences. Trends Mol. Med., 16, 238–246.
26) Tchkonia, T., Zhu, Y., van Deursen, J., Campisi, J., & Kirkland, J.L. (2013) Cellular senescence and the senescent secretory phenotype: Therapeutic opportunities. J. Clin. Invest., 123, 966–972.
27) Wiley, C.D., Brumwell, A.N., Davis, S.S., Jackson, J.R., Valdovinos, A., Calhoun, C., Alimirah, F., Castellanos, C.A., Ruan, R., Wei, Y., et al. (2019) Secretion of leukotrienes by senescent lung fibroblasts promotes pulmonary fibrosis. JCI Insight, 4, e130056.
28) Arai, Y., Martin-Ruiz, C.M., Takayama, M., Abe, Y., Takebayashi, T., Koyasu, S., Suematsu, M., Hirose, N., & von Zglinicki, T. (2015) Inflammation, but not telomere length, predicts successful ageing at extreme old age: A longitudinal study of semi-supercentenarians. EBioMedicine, 2, 1549–1558.
29) Hashimoto, K., Kouno, T., Ikawa, T., Hayatsu, N., Miyajima, Y., Yabukami, H., Terooatea, T., Sasaki, T., Suzuki, T., Valentine, M., et al. (2019) Single-cell transcriptomics reveals expansion of cytotoxic CD4 T cells in supercentenarians. Proc. Natl. Acad. Sci. USA, 116, 24242–24251.
30) Miura, Y., Hashii, N., Tsumoto, H., Takakura, D., Ohta, Y., Abe, Y., Arai, Y., Kawasaki, N., Hirose, N., & Endo, T.; SONIC (Septuagenarians, Octogenarians, Nonagenarians Investigation with Centenarians) (2015) Change in N-glycosylation of plasma proteins in Japanese semisupercentenarians. PLoS One, 10, e0142645.
31) Miura, Y., Hashii, N., Ohta, Y., Itakura, Y., Tsumoto, H., Suzuki, J., Takakura, D., Abe, Y., Arai, Y., Toyoda, M., et al. (2018) Characteristic glycopeptides associated with extreme human longevity identified through plasma glycoproteomics. Biochim. Biophys. Acta, 1862, 1462–1471.
32) Takahashi, S., Sugiyama, T., Shimomura, M., Kamada, Y., Fujita, K., Nonomura, N., Miyoshi, E., & Nakano, M. (2016) Site-specific and linkage analyses of fucosylated N-glycans on haptoglobin in sera of patients with various types of cancer: Possible implication for the differential diagnosis of cancer. Glycoconj. J., 33, 471–482.
33) Zhu, J., Chen, Z., Zhang, J., An, M., Wu, J., Yu, Q., Skilton, S.J., Bern, M., Ilker Sen, K., Li, L., et al. (2019) Differential quantitative determination of site-specific intact N-glycopeptides in serum haptoglobin between hepatocellular carcinoma and cirrhosis using LC-EThcD-MS/MS. J. Proteome Res., 18, 359–371.
34) Zhang, S., Jiang, K., Sun, C., Lu, H., & Liu, Y. (2013) Quantitative analysis of site-specific N-glycans on sera haptoglobin beta chain in liver diseases. Acta Biochim. Biophys. Sin. (Shanghai), 45, 1021–1029.
35) Thomsen, J.H., Etzerodt, A., Svendsen, P., & Moestrup, S.K. (2013) The haptoglobin-CD163-heme oxygenase-1 pathway for hemoglobin scavenging. Oxid. Med. Cell. Longev., 2013, 523652.
36) Otterbein, L.E., Bach, F.H., Alam, J., Soares, M., Tao Lu, H., Wysk, M., Davis, R.J., Flavell, R.A., & Choi, A.M. (2000) Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat. Med., 6, 422–428.
37) Miura, Y., Sato, Y., Arai, Y., Abe, Y., Takayama, M., Toda, T., Hirose, N., & Endo, T. (2011) Proteomic analysis of plasma proteins in Japanese semisuper centenarians. Exp. Gerontol., 46, 81–85.
38) Nishikaze, T., Tsumoto, H., Sekiya, S., Iwamoto, S., Miura, Y., & Tanaka, K. (2017) Differentiation of sialyl linkage isomers by one-pot sialic acid derivatization for mass spectrometry-based glycan profiling. Anal. Chem., 89, 2353–2360.

東京都健康長寿医療センター研究所老化機構研究チーム研究副部長.博士(薬学)
1989年東京大学大学院薬学系研究科修士課程修了.その後,昭和大学薬学部助手,放射線医学総合研究所(現・量子科学技術研究開発機構)科学技術特別研究員,東京都老人総合研究所研究員を経て,2012年より現職.
研究テーマと抱負老化や老化関連疾患で生じるタンパク質や糖鎖変化の解析から,バイオマーカー探索や病態メカニズムに関する研究を進めている.

東京都健康長寿医療センター研究所老化機構研究チーム研究員.博士(薬学).
2005年名古屋市立大学大学院薬学研究科博士後期課程中退.その後,武蔵野大学薬学部助手,名古屋市立大学大学院薬学研究科特任助教,京都大学大学院薬学研究科特定助教を経て,2013年より現職.
研究テーマと抱負質量分析法を基盤とする新規分析手法の開発と応用.
東京都健康長寿医療センター研究所シニアフェロー.
2019年より現職.その他については本誌89巻5号(2017),p. 612をご参照ください.

This page was created on 2020-04-17T14:46:26.676+09:00
This page was last modified on 2020-05-28T08:14:24.000+09:00
このサイトは(株)国際文献社によって運用されています。