糖鎖立体構造解析技術の開発要素Techniques for analyzing 3D structure and dynamics of glycan
東北医科薬科大学Tohoku Medical and Pharmaceutical University ◇ 宮城県仙台市青葉区小松島4–4–1 ◇ 4–4–1 Komatsushima, Aoba-ku, Sendai, Miyagi 981–8558, Japan
発行日:2020年6月25日Published: June 25, 2020
真の生命像を俯瞰的に捉えるためには,未解明問題が多く残っている糖鎖の研究を積極的に推進することが急務である.糖鎖の生理的機能を解明するためには,生化学的手法(酵素による糖鎖の切断),分子生物学的手法(糖転移酵素のノックアウト)などさまざまな手法が考えられるが,中でも糖鎖の化学構造を決定し,その立体構造・運動性や相互作用様式を調べるアプローチは,糖鎖の機能を視覚的に理解することを促し,説得力がある.いわゆる構造生物学的な考えであるが,糖鎖を対象とする場合は,他の生体高分子と異なる特異な性質を十分理解する必要がある.本稿では糖鎖の立体構造・運動性・相互作用について概説し,糖鎖の立体構造や相互作用について筆者が普段よく質問されることを中心にQ&A形式でまとめ,最後に立体構造解析技術の現状と展望を述べたい.
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
生体分子としての糖鎖を核酸やタンパク質など他の生体高分子と比較すると,共通点も存在するが,相違点がきわだっている.核酸もタンパク質も糖鎖も,構成ユニット(ビルディングブロック)がつながって高分子ができ上がっているという点では共通であるが,その構成ユニットとその結合様式を比較すると糖鎖の特徴を理解することができる(図1).核酸の場合は,ヌクレオチドがホスホジエステル結合で,タンパク質の場合はアミノ酸がペプチド結合(アミド結合)でつながっており,結合様式は一義的である.一方で,糖鎖の場合は,構成ユニット(主に単糖)間の水酸基(ヒドロキシ基)どうしが脱水縮合してグリコシド結合を形成しており,その結合様式は立体化学・結合部位の点から実に多様である.まさにこの点が,糖鎖の化学構造の多様性を生み出していることと密接に関係し,糖鎖は多機能な情報分子としての潜在能力を秘めている.別の観点からいえば,核酸やタンパク質はその結合様式の特徴から直鎖構造しか形成することができないが,糖鎖は構成ユニット中に水酸基を複数持つことから,分岐構造(枝分かれ構造)を形成することが可能である.この分岐構造については,糖鎖と糖鎖結合分子(レクチン,抗体など)との相互作用を理解するときに,特に注意を払うべき点である.というのは,複数の分岐はしばしば同じ構造ユニットを有しており,1本の糖鎖あたり複数のタンパク質相互作用部位を有するからである.

糖鎖は核酸やタンパク質と異なり,分岐構造の形成が可能である.
糖鎖は遊離の状態,もしくはタンパク質や脂質と結合した状態(複合糖質)として生体内で存在している.タンパク質と糖鎖の結合様式も複数存在すること,また脂質自身も脂質の鎖長の違いなどで構造多様性を有することから,複合糖質としてはいっそう多様性を増すことになる.核酸の場合でもリボースやデオキシリボースといった糖がその構成ユニットとして入り込んでおり,核酸も糖質と無縁ではない.細胞質や核内にもリン酸化と同じように糖修飾が見いだされ,多くは競合する関係にある.このように糖あるいは糖鎖は生体内に広く分布しており,さまざまな形で見いだされる.したがって糖鎖・糖質を対象とする糖鎖生物学は,さまざまな研究分野に浸透するべきものであろう.
糖鎖に限らず,生体分子の世界を見たいと思う.分子の世界を手に取るように見ることができれば,その分子の働きを容易に理解することができるであろう.しかし,原子レベルの分解能で分子を見るには,特殊な方法を用いる必要がある.その方法については後述するとして,ここでは糖鎖の立体構造の表現方法について述べたい.
糖鎖の立体構造を明らかにするということは,糖鎖を構成する各原子の座標(x, y, z)を明らかにするということと同義である.座標を明らかにすることができれば,適当なソフトウェアを用いることにより,糖鎖を立体構造表示することができ,分子の世界をのぞき込むことが可能になる.一方で,糖鎖の立体構造を比較したり,運動性を評価したりする際に,糖鎖の立体構造を表現するパラメータを定義すると便利である.そのパラメータが,単糖間のグリコシド結合周りの二面角である.都合がよいことに,糖鎖を構成する単糖はほとんどの場合環状構造となっており,その環状構造は通常エネルギー的に安定ないす形構造を占めている.そのため,糖鎖の全体的な立体構造を捉える場合は,環状構造部分は固定して考えてよく,自由度のあるグリコシド結合周りの二面角のみを調べればよいことになる.これはタンパク質主鎖のコンホメーションを考えるときに,主鎖二面角を表示したラマチャンドラン・プロットを用いることと似ている.
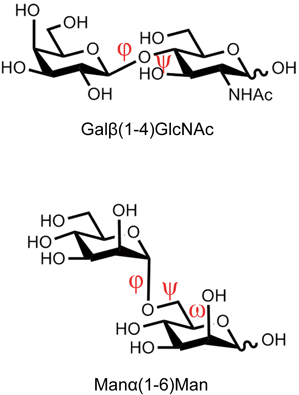
糖鎖のグリコシド結合周りの二面角は,二面角を構成する連続した四つの原子の座標から規定される(図2).たとえば六員環の単糖(i)と単糖(i−1)が1–4結合でつながっている場合,以下の二つの二面角φ(ファイ)とψ(プサイ)を用いることがIUPAC-IUB合同委員会により推奨されている1).
さてNMR法により得られるパラメータ,特に3Jカップリング定数を用いて糖鎖のグリコシド結合周りの二面角を明らかにすることができる.3Jカップリング定数は以下のKarplusの式によって二面角の情報に変換できるからである.
NMR法では主に水素原子を観測対象とするため,3Jカップリング定数と二面角を関連づける場合,以下のような水素原子を含めた二面角の定義が用いられることもある.
X線結晶構造解析から得られた糖鎖の像は水素原子の情報が含まれていないことが多いので,その場合は前者の水素原子を使わない定義を用いることになる.論文間で二面角の定義が異なることもあり,データを比較して議論する際には二面角の定義を確認する必要がある.
多くの場合,グリコシド結合周りの二つの二面角φとψで糖鎖のコンホメーションを描写することが可能であるが,一部の結合(1-6結合など)についてはφとψに加えて,ω(オメガ)など追加の二面角が必要になる(図2下).
六員環の環状糖のリング部分は先に述べたようにいす形がエネルギー的に安定であり,多くの場合,環状構造はいす形を占めていると考えてよい.しかし,酵素反応の中間体(遷移状態)としてのリガンド糖鎖,あるいは硫酸化を過度に受けた糖残基などはいす形から逸脱することがあるので注意を促したい2).また核酸を構成するリボース,デオキシリボースなど五員環は,六員環よりも環状構造は柔らかい.通常,糖の環状構造(パッカリング)はCremerとPopleによって提案された三つのパラメータによって表現される3).六員環の場合,いす形(C)2個に加えて舟形(B)6個,ねじれ舟形(S)6個,封筒形(E)12個,半いす形(H)12個の合計38個に分類される.
パッカリングの重要性を示す一例として,抗インフルエンザウイルス薬をあげることができる.リレンザやタミフルはシアル酸の誘導体であるが,その環状構造内に炭素–炭素間の二重結合が導入されている.これは二重結合の導入によりいす形を崩し,ウイルス由来のノイラミニダーセ(シアリダーゼ)の基質の遷移状態を模倣している.もう一つの例として細菌のラムノース転移酵素をあげたい.ラムノースを付加する糖転移酵素EarPは,細菌の伸長因子EF-Pの特定のArg残基(Arg32)をラムノース化する4).EarPの結晶構造解析からラムノースはいす形で結合していたが5),想定されるSN2反応(立体化学の反転を伴う反応)を進行させるためには,いす形では反応が進まず,ねじれ舟形のようないす形とは異なる形になることが予想された.実際に分子動力学計算を行うと,いす形ではないラムノースの形が安定な形として見いだされた.これらのことから,酵素反応は非いす形を経て反応が進むことが考えられた5).他に,ERマンノシダーゼにおいてもその結晶構造からリガンド糖鎖の非いす形のコンホメーションが議論されている6).
最近になり,環状構造を持たない直鎖状のユニット(リビトール)が哺乳動物の糖鎖に存在することも見いだされた7, 8).このリビトール内および連結部分のコンホメーションを表現する場合には通常より多くの二面角が必要になる.新しいメンバーであるリビトールの安定なコンホメーションを実験および計算の両面から調べることは今後の課題である9, 10).
さて糖鎖の立体構造と相互作用については,タンパク質や核酸の場合と比較して圧倒的に情報量が少ないことから,統計的に調べることが困難な状況である.たとえばPDBを用いて糖鎖の像が得られている糖タンパク質の立体構造を調べた場合,その数は2015年6月の時点で重複を除くと100を超えていない11).限られた例数で我々の糖鎖の立体構造・相互作用についての理解は十分ではないが,それでも糖鎖を対象とした先駆的な研究報告も多数存在する.そこで本節では,筆者がこれまで糖鎖の立体構造や相互作用に関して受けた質問を中心に疑問点をあげて,過去の論文を参照しつつQ&A形式で回答してみたい.
糖タンパク質を結晶化しているにもかかわらず,糖鎖の像が得られていない場合がある.これは大きく分けて二つの場合が考えられる.一つは,結晶化の効率を上げるために,何らかの方法で糖タンパク質の糖鎖をあらかじめ取り除いている場合である.糖鎖をグリコシダーゼで除く,あるいは糖鎖付加がそもそもできない大腸菌で糖タンパク質を発現している場合などがある.もう一つは,糖鎖が結合しているにもかかわらず電子密度像として得られていない場合である.これは主に糖鎖のグリコシド結合周りの二面角の自由度に由来すると考えられる.タンパク質の結晶構造解析において,N末端やC末端,あるいはループ部分など比較的運動性の高いと思われる領域の電子密度像が得られない事象と似ている.このような場合,糖鎖の全体像はわからないが,糖鎖が部分的に像を与えることもしばしばみられる.Asnに結合しているN-アセチルグルコサミン1~2残基はよく観測されるものの,それ以降の電子密度像が得られていないケースである.これはAsnに結合している糖鎖の根本付近は比較的堅く,自由度が比較的乏しいことに対応していると考えられる.糖鎖を切断しているからその像が見えないのか,それとも糖鎖は存在するけれど,その自由度が高いために像が得られていないのかは,立体構造のデータだけからは判断できない場合が多く,元の論文をひもとく必要がある.
一方で,例外的にほとんど完全に糖鎖の電子密度像が得られることがある.代表的な例として,抗体のFc領域に結合している糖鎖があげられる12).1対の二本鎖複合型糖鎖が結合しているが,一部の糖残基を除き,全体像を観察することができる.これは糖鎖がFc分子の比較的内部に埋もれており,糖鎖とポリペプチド鎖が広範囲で分子内相互作用しており,糖鎖の運動性が比較的抑制されていることと関係している13, 14).
糖鎖の立体構造を知ることは,その機能をよく理解するための大事なステップである.たとえば,糖タンパク質糖鎖の立体構造を知るためには,糖タンパク質のままで結晶構造解析を行えばよいことになる.しかしながら,結晶が得られたとしても,先に述べた理由により糖鎖の電子密度像を得られない場合が多い.その場合は,糖鎖の立体構造に関する情報を積極的に得るためには他の手法に依存することになる.ありうる方法の中で溶液NMR法は,水溶液中における生体分子の立体構造・運動性・相互作用に関する情報を原子レベルで提供する方法である.これまで糖鎖・糖タンパク質のNMR解析がなされてきたが,立体構造を決定した報告は多くなく,その背景には糖鎖のNMR解析の難しさがある.これにはいくつか理由がある.第一に糖鎖の立体構造を規定するために利用できるNMRパラメータの少なさである.立体構造を制限するための十分な情報が得られないため,立体構造を一義的に規定するに至らない場合が多い.距離情報を提供する水素原子数が少ないこととも関係する.第二に本質的な問題として,糖鎖は一つの形をとっているのではなく,多くの場合複数のコンホマー間をすばやく遷移していることがあげられる.立体構造を規定する情報が少ない上に,1形でないとすると,その立体構造の描写は著しく困難になる.複数の安定構造からなる糖鎖の立体構造の描写を行うためには,(準)安定コンホマーを一つ一つ明らかにするととともに,その占有数(エネルギー差)や遷移速度を明らかにする必要がある.筆者らは,複数のレクチンを使って一つの共通の糖鎖と結合させ,各糖鎖-レクチン複合体の結晶構造解析を行うことにより,糖鎖の一つ一つのコンホマーを明らかにすることを試みた.その方法により,これまで一過的にしか捉えることのできなかった糖鎖の折れ曲がり構造が実験的に可視化された15).
糖鎖の立体構造を実験的に決定することが難しいため,分子動力学計算などの計算化学的な手法による糖鎖の立体構造の解明は重要な位置を占めている.分子動力学計算によって明らかにされた糖鎖の動的振る舞いをながめていると,静的なスナップショットをながめていただけでは出てこないような新しいインスピレーションも湧いてくる.ただ,実験を行う立場から気になる点は,計算によって得られた立体構造やダイナミクスは,どの程度本来の水溶液中の状態を反映しているのか,言葉を変えると計算結果はどの程度信頼できるか,である.この問いに答えるのは容易ではない.その理由として,実験的に得られた糖鎖の立体構造やダイナミクスの情報がそもそも多くないことがあげられる.それでも,これまでの先駆的な研究において計算データとNMRデータの比較がなされており,報告によると(準)安定構造と各構造の占有数は計算化学によっておおよそ再現することができるという16, 17).しかし,現状でも,どの力場を用いるか,水分子をどう扱うか,計算時間は十分か,実験結果と計算結果が対応しているかなど常に注意を払う必要がある.筆者の経験から,(N-アセチル基のない)グルコサミンやリビトールリン酸,硫酸化を受けた糖鎖などに対して力場のパラメータが設定されているかどうかを計算時に確認する必要がある2).
糖鎖は柔軟な構造をしているため,複数の安定な構造をすばやく遷移しているが,タンパク質と相互作用する際に,その中のどのコンホマーが結合時に選択されるかは,糖鎖構造生物学の関心事の一つである.複数のコンホマーの存在と各コンホマーの占有率は通常,分子動力学計算などの手法により明らかにされるが,興味深いことにその占有数比はPDB中のリガンド糖鎖の各立体構造の出現頻度とよく一致する.すなわち,1分子を対象とした計算により得られたエネルギー的に安定なコンホメーションは,タンパク質と糖鎖の複合体の結晶構造中で統計的に頻度高くみられるコンホメーションと一致する18).したがって,「糖鎖はタンパク質と相互作用するときにコンホメーションが変化するか?」の質問に対する答えとしては,「糖鎖のコンホメーションは変化しない」が最も妥当な答えになる.もちろん,糖鎖のタンパク質結合部分は運動性の低下を伴うので,そのイメージを持ちながらの答えであり,例外も存在する.筆者らのグループが最近行った糖鎖とタンパク質の相互作用の例では,タンパク質の結合に伴い,遊離の糖鎖では観測されなかったコンホメーションでタンパク質と結合することが計算化学により示された19).
通常単糖とタンパク質の一価(1対1)の相互作用は弱く,解離定数にしておおよそmM程度である場合が多い.なぜ相互作用が弱いのか.それは,結合自由エネルギーの内訳にヒントが隠されている.解離定数Kd(結合定数K)は結合自由エネルギーΔGと関係し,結合自由エネルギーはエンタルピー項ΔHとエントロピー項TΔSに分けることができる. ここでRは気体定数,Tは絶対温度である.糖鎖とタンパク質の相互作用の多くの場合,ΔHがΔGに大きく寄与しており,TΔSは通常結合に不利に働いている.このΔHとTΔSがほぼ相殺し,そのわずかな差がΔGとなっている.せっかくΔHで相互作用に貢献してもΔSが大きく足を引っ張っているのである.一例として,抗糖鎖抗体と糖鎖(ポリシアル酸DP~100)との相互作用の場合,ΔH=−80.1 kcal/mol, TΔS=−72.7 kcal/molであり,ΔHとTΔSがほぼ相殺してΔG=−7.4 kcal/mol(Kd=3.5 μM)となっている20).
ΔSの解釈は単純ではないが,水分子の再配置21)や糖鎖のコンホメーションの柔軟性の喪失22)によるものと考えられている.ΔSを人為的に小さくすることができれば,高親和性の糖鎖リガンドも原理的に開発することが可能である.自然界では,その弱い相互作用を克服するために多価での相互作用が利用されているようである.IgG抗体が二価で働いているように,糖鎖側およびタンパク質(レクチン)側も多価になっている場合が多く,みかけの親和性(avidity)は一価の相互作用の場合の親和性と比べて格段に強くなる.一般にその見かけの親和性は,それぞれ一価の結合定数の掛け算に匹敵する. こうして多価の相互作用が実現されたときにはμM程度の親和性となり,実効的には問題のない結合力となる.
タンパク質の半数以上は糖鎖修飾を受けているという見積もりがあり,その糖鎖の役割は何なのか,タンパク質の立体構造に与える影響はどの程度なのか,糖鎖生物学の課題の一つである.タンパク質に結合している糖鎖の機能を大きく二つに分けると,糖鎖そのものがレクチンで認識されてシグナルとして働く場合(分子間相互作用)と,糖鎖がタンパク質の安定化に寄与している場合(分子内相互作用)になる23).ここで問題としたいのは,糖鎖とタンパク質の分子内相互作用がタンパク質の立体構造に及ぼす影響である.糖タンパク質から糖鎖を取り除くと,そのタンパク質は細胞外へ分泌されなくなる,溶解度が減少する,立体構造形成がうまくいかなくなる,生理活性が喪失・低下するなどの報告がある.このことから糖鎖はタンパク質の立体構造や物理化学的性質に影響を与えているといえる.具体的に糖鎖がタンパク質の立体構造にどのような影響を及ぼすかは,タンパク質ごと,糖鎖修飾部位ごとで議論することが必要になるが,Asnに糖鎖修飾を受けたタンパク質とそうでないタンパク質を統計的に比較すると,糖鎖修飾によりAsnの側鎖コンホメーションの自由度が制限される傾向にあるという23–25).また糖鎖の欠損により,一般的にタンパク質の運動性(揺らぎ)が増す傾向にあるようである26, 27).ただ現状では報告例が多くなく,今後のさらなる解析が必要になる.
糖鎖を含めた生体分子の立体構造解析を行う場合には,構造生物学的な手法が必要になる.原子レベルの分解能を提供する手法といえばX線結晶構造解析が王道であるが,溶液NMR法,固体NMR法に加えて最近では電子顕微鏡による解析が雑誌を賑わす存在になった.現在PDB中には約159,000個の立体構造情報が登録されており(2019年12月15日現在),そのうちX線は141,000個と圧倒的に多く,次にNMRの13,000個,次に電子顕微鏡の4,000個である.電子顕微鏡により決定された立体構造数の伸びは著しい(図3).ここでは各手法の特徴と糖鎖に関連する最近の進展をまとめたい.
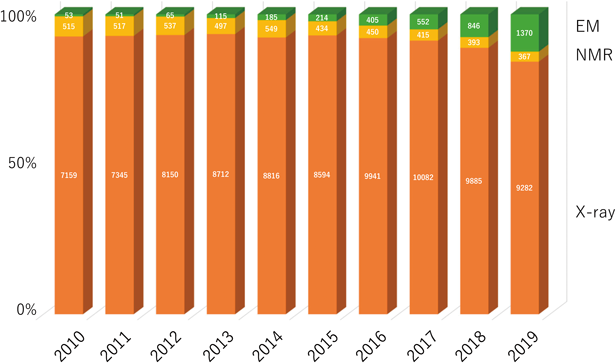
各年に登録された立体構造全体をそれぞれ100%とし,各解析手法[X線結晶構造解析(X-ray),NMR,電子顕微鏡(EM)]の割合を示した.棒グラフ中に示している数字は実際の立体構造数である.近年,電子顕微鏡によって解析された立体構造の割合が増加している.
最近になり,生体高分子のクライオ電子顕微鏡(以下クライオ電顕)による観察例は増加の一途をたどっている.膜タンパク質やタンパク質線維,ウイルス,細胞など適用範囲はさまざまである.試料の調製の改善が進んだこともあるが,コンピュータに制御されたデータの取得とデータ処理の進展によるところが大きい.古典的な電子顕微鏡では,固定化や染色などが必要であり,そのステップで生体試料にダメージを与えることもしばしばであったが,クライオ電顕では薄い氷の中に閉じ込められた試料を可視化するため,生理的条件下により近い条件で測定することが可能である.ランダムに配向した分子の単粒子解析を行うことにより,3次元構造をおおよそ原子レベルの分解能で構築することができる.対称性の高い球状のウイルスなどは,電顕による構造解析に適した試料といえる.
糖タンパク質を対象とする場合,クライオ電顕は魅力的である.結晶化の必要がなく,糖鎖の不均一性や柔軟性は深刻な問題にはならない.仮に糖鎖が複数のコンホメーションからなっていたとしても,適切な分類により解決できるかもしれない.
糖鎖の像が得られている例をいくつか紹介する.ジカウイルスのクライオ電顕像が3.8 Åの分解能で2016年に報告されたが,大変興味深いことに,Asn154に結合している糖鎖の像が部分的ではあるものの観測されている(図4左)28).その糖鎖は,宿主側のC型レクチン受容体DC-SIGNとの結合を介しているということが想定されている.実際デングウイルスの糖鎖とDC-SIGNの相互作用のようすが低分解能ながらクライオ電顕により調べられており,糖鎖のオリエンテーションや立体構造の重要性を示している29).同様に糖鎖の重要性が指摘されているウイルスがHIVである.これまでいくつかの報告がなされているが,Bjorkmanらの論文では詳細にエンベロープ糖タンパク質の糖鎖の構造について述べており,結晶構造とクライオ電顕から得られた糖鎖の像を比較している30).残念ながらクライオ電顕像の結果からは,その分解能の低さから糖鎖全体のコンホメーションを議論することは妨げられたが,高分解能の結晶構造のデータを用いて,低分解能の電子密度像への当てはめを行って議論することが可能であった.
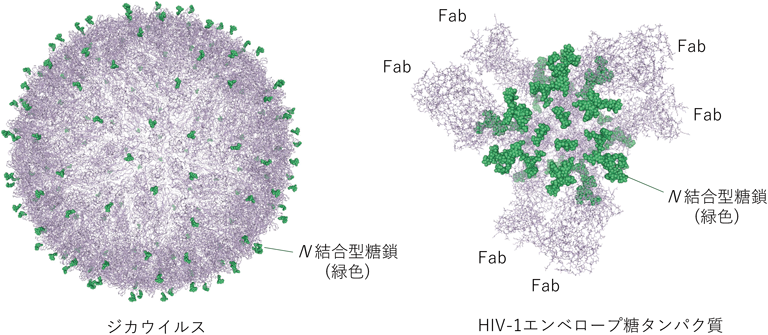
いずれの図においてもN結合型糖鎖を緑で示している.ジカウイルスのAsn154に結合しているN結合型糖鎖の像は部分的に観測されている(GlcNAcβ1-4GlcNAcβ1-Asn).エンベロープ糖タンパク質(三量体)には2種類のFab(合計六つ)が結合している.立体構造表示はNGL viewerにて行った46).
またクライオ電顕像に限ったことではないが,結晶構造解析も含めて低分解能の糖鎖密度像を用いた解析の場合,糖鎖の立体化学の間違えや,環の不自然なゆがみなどを生み出すことがしばしばある31).実際PDB中の糖鎖構造には多くのエラーが存在しており,そのデータを用いた統計的な解析には注意を要する.コンホメーションや立体化学のエラーの検出や修正を行う方法も報告されており32–34),クライオ電顕により得られた低分解能の電子密度に正しく糖鎖を当てはめて構造精密化がなされている.
構造生物学的手法の中で最も実績があるのがX線結晶構造解析であり,高分解能の情報を提供する.これまで多くの糖タンパク質や糖鎖-タンパク質複合体などが原子レベルで解明されてきた.他の手法を用いる場合でも同様であるが,試料の調製が鍵となる.いかに良質な試料を調製できるかが結晶化・構造解析の成否を決めるといってもよい.
糖タンパク質の場合,多くの場合は糖鎖の像が得られないが,しばしば電子密度像が得られることがある.糖鎖とタンパク質が分子内で広範に相互作用している場合は,先に述べたようにほぼ完全な状態で糖鎖の電子密度像を得られることがある.たとえば極端な例として,HIV-1のエンベロープ糖タンパク質は高マンノース型糖鎖や複合型糖鎖が高密度で結合しているにもかかわらず,2種類のFabを結合させることによって結晶構造が得られている35)(図4右).通常であれば糖鎖を均一にするなどの方策をとるのが一般的であるが,これは糖鎖をnativeな状態で結晶化することに成功しており大変意義深い.一方で,糖タンパク質の結晶構造解析で注意しなければならないことは,結晶中では分子がパッキングされているため,生理的には観測されないような相互作用が強いられている可能性もあることである.糖分解酵素や糖転移酵素の結晶化を行ったところ,自身のAsnに結合している糖鎖が隣接分子の基質結合部位に収まって,基質を模倣している場合があった36, 37).その相互作用により結晶化が促進されている面もあるが,結晶中における糖鎖のコンホメーションや相互作用を議論するときは,それが生理的条件下・水溶液中でも起こるかどうかを常に意識しなければならない.
NMR法,特に溶液NMR法は,水溶液中における原子レベルの情報を提供する唯一の方法である.溶液NMR法は糖鎖の運動性を調べるのに特に適した方法である.運動性はNMRパラメータのうちの緩和時間が適しており,緩和時間を用いた糖鎖のダイナミクス研究も報告例が多数存在する13, 38, 39).一方で,糖鎖の立体構造を調べるためには,立体構造を規定するためのNMRパラメータとしてNOE(核オーバーハウザー効果)と3Jカップリング定数があげられるが,糖鎖は水素の密度が少ないこともあり,この二つだけでは不十分な場合が多い.そのため,追加で立体構造を規定するようなパラメータが必要になり,RDC(残余双極子カップリング)やPCS(擬コンタクトシフト)などを利用した糖鎖のコンホメーション解析が進んでいる40).特に多分岐糖鎖において,PCSなどを利用することにより,各分枝に由来する信号を区別して議論できるようになったことは大きなブレークスルーである41–43).筆者らも糖鎖のコンホメーションを解析するための新たなNMRパラメータとして,糖残基間の1H-13C異核NOEやグリコシド酸素をセレンに置換したセレン含有糖鎖における77Seカップリング定数の利用44)などを検討している.セレンに由来するNMR信号は化学シフトの範囲が広く,かつレクチンとの結合を鋭敏に反映するため(図5),生体溶液中(血清中など)での相互作用の特異的検出など今後の応用が期待される45).
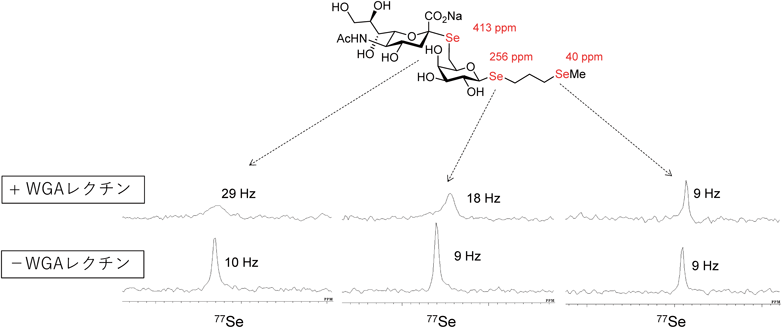
各NMRシグナルの線幅(半値幅)をHz単位で図中に示している.WGAレクチンの添加により,特定のセレンシグナルの線幅の広幅化が観測されている.一方でWGAレクチンの結合に関与していない部分に由来するセレンシグナルの線幅はほとんど変化していない.このことから,WGAレクチンの相互作用部位を同定することができる.
糖鎖の立体構造からその機能を理解する「糖鎖構造生物学」は,これからより普及していく必要がある.主役はタンパク質であることは間違いないが,その機能調節をドラスティックに行っているのは脇役の糖鎖である.受容体の機能を糖鎖に着目すると,新しい側面が見えてくることもある.そのためには,糖鎖を対象とした構造生物学的手法の開発が必要であり,一つの方法だけではなく,計算化学的手法も含めた複数の手法の組合わせが必要になる.今後異分野のますますの連携が必要になる.また新しい技術開発は新しい知見をもたらすのが常であり,構造生物学分野のさらなる技術開発も必要である.糖鎖の立体構造・運動性・相互作用をより理解することができれば,医薬品の開発につながることも期待される.いままでにない視点からのアプローチも可能になるため,筆者も医薬品の開発において「糖鎖構造生物学分野」から貢献することを意識している.
1) IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). (1983) Symbols for Specifying the Conformation of Polysaccharide Chains Recommendations 1981. Eur. J. Biochem., 131, 5–7.
2) Singh, A., Tessier, M.B., Pederson, K., Wang, X., Venot, A.P., Boons, G.J., Prestegard, J.H., & Woods, R.J. (2016) Extension and validation of the GLYCAM force field parameters for modeling glycosaminoglycans. Can. J. Chem., 94, 927–935.
3) Cremer, D. & Pople, J.A. (1975) General definition of ring puckering coordinates. J. Am. Chem. Soc., 97, 1354–1358.
4) Lassak, J., Keilhauer, E.C., Furst, M., Wuichet, K., Godeke, J., Starosta, A.L., Chen, J.M., Sogaard-Andersen, L., Rohr, J., Wilson, D.N., et al. (2015) Arginine-rhamnosylation as new strategy to activate translation elongation factor P. Nat. Chem. Biol., 11, 266–270.
5) Sengoku, T., Suzuki, T., Dohmae, N., Watanabe, C., Honma, T., Hikida, Y., Yamaguchi, Y., Takahashi, H., Yokoyama, S., & Yanagisawa, T. (2018) Structural basis of protein arginine rhamnosylation by glycosyltransferase EarP. Nat. Chem. Biol., 14, 368–374.
6) Karaveg, K., Siriwardena, A., Tempel, W., Liu, Z.J., Glushka, J., Wang, B.C., & Moremen, K.W. (2005) Mechanism of class 1 (glycosylhydrolase family 47) alpha-mannosidases involved in N-glycan processing and endoplasmic reticulum quality control. J. Biol. Chem., 280, 16197–16207.
7) Manya, H., Yamaguchi, Y., Kanagawa, M., Kobayashi, K., Tajiri, M., Akasaka-Manya, K., Kawakami, H., Mizuno, M., Wada, Y., Toda, T., et al. (2016) The Muscular Dystrophy Gene TMEM5 Encodes a Ribitol β1,4-Xylosyltransferase Required for the Functional Glycosylation of Dystroglycan. J. Biol. Chem., 291, 24618–24627.
8) Kanagawa, M., Kobayashi, K., Tajiri, M., Manya, H., Kuga, A., Yamaguchi, Y., Akasaka-Manya, K., Furukawa, J.I., Mizuno, M., Kawakami, H., et al. (2016) Identification of a Post-translational Modification with Ribitol-Phosphate and Its Defect in Muscular Dystrophy. Cell Rep., 14, 2209–2223.
9) Klein, R.A., Hartmann, R., Egge, H., Behr, T., & Fischer, W. (1994) The aqueous solution structure of the tetrasaccharide-ribitol repeat-unit from the lipoteichoic acid of Streptococcus pneumoniae strain R6 determined using a combination of NMR spectroscopy and computer calculations. Carbohydr. Res., 256, 189–222.
10) Hatcher, E., Guvench, O., & Mackerell, A.D. Jr. (2009) CHARMM Additive All-Atom Force Field for Acyclic Polyalcohols, Acyclic Carbohydrates and Inositol. J. Chem. Theory Comput., 5, 1315–1327.
11) Suga, A., Nagae, M., & Yamaguchi, Y. (2018) Analysis of protein landscapes around N-glycosylation sites from the PDB repository for understanding the structural basis of N-glycoprotein processing and maturation. Glycobiology, 28, 774–785.
12) Deisenhofer, J. (1981) Crystallographic refinement and atomic models of a human Fc fragment and its complex with fragment B of protein A from Staphylococcus aureus at 2.9- and 2.8-A resolution. Biochemistry, 20, 2361–2370.
13) Yamaguchi, Y., Kato, K., Shindo, M., Aoki, S., Furusho, K., Koga, K., Takahashi, N., Arata, Y., & Shimada, I. (1998) Dynamics of the carbohydrate chains attached to the Fc portion of immunoglobulin G as studied by NMR spectroscopy assisted by selective 13C labeling of the glycans. J. Biomol. NMR, 12, 385–394.
14) Yamaguchi, Y. & Barb, A.W. (2019). Glycobiology.
15) Nagae, M., Kanagawa, M., Morita-Matsumoto, K., Hanashima, S., Kizuka, Y., Taniguchi, N., & Yamaguchi, Y. (2016) Atomic visualization of a flipped-back conformation of bisected glycans bound to specific lectins. Sci. Rep., 6, 22973.
16) Nishima, W., Miyashita, N., Yamaguchi, Y., Sugita, Y., & Re, S. (2012) Effect of bisecting GlcNAc and core fucosylation on conformational properties of biantennary complex-type N-glycans in solution. J. Phys. Chem. B, 116, 8504–8512.
17) Re, S., Miyashita, N., Yamaguchi, Y., & Sugita, Y. (2011) Structural diversity and changes in conformational equilibria of biantennary complex-type N-glycans in water revealed by replica-exchange molecular dynamics simulation. Biophys. J., 101, L44–L46.
18) Imberty, A. (1997) Oligosaccharide structures: theory versus experiment. Curr. Opin. Struct. Biol., 7, 617–623.
19) Nagae, M., Mishra, S.K., Neyazaki, M., Oi, R., Ikeda, A., Matsugaki, N., Akashi, S., Manya, H., Mizuno, M., Yagi, H., et al. (2017) 3D structural analysis of protein O-mannosyl kinase, POMK, a causative gene product of dystroglycanopathy. Genes Cells, 22, 348–359.
20) Nagae, M., Ikeda, A., Hane, M., Hanashima, S., Kitajima, K., Sato, C., & Yamaguchi, Y. (2013) Crystal structure of anti-polysialic acid antibody single chain Fv fragment complexed with octasialic acid: insight into the binding preference for polysialic acid. J. Biol. Chem., 288, 33784–33796.
21) Lemieux, R.U., Delbaere, L.T., Beierbeck, H., & Spohr, U. (1991) Involvement of water in host-guest interactions. Ciba Found. Symp., 158, 231–245, discussion, 245–238.
22) Carver, J.P. (1993) Oligosaccharides: How can flexible molecules act as signals? Pure Appl. Chem., 65, 763–770.
23) Nagae, M. & Yamaguchi, Y. (2012) Function and 3D structure of the N-glycans on glycoproteins. Int. J. Mol. Sci., 13, 8398–8429.
24) Lütteke, T. (2009) Analysis and validation of carbohydrate three-dimensional structures. Acta Crystallogr. D Biol. Crystallogr., 65, 156–168.
25) Petrescu, A.J., Milac, A.L., Petrescu, S.M., Dwek, R.A., & Wormald, M.R. (2004) Statistical analysis of the protein environment of N-glycosylation sites: implications for occupancy, structure, and folding. Glycobiology, 14, 103–114.
26) Lee, H.S., Qi, Y., & Im, W. (2015) Effects of N-glycosylation on protein conformation and dynamics: Protein Data Bank analysis and molecular dynamics simulation study. Sci. Rep., 5, 8926.
27) Joao, H.C., Scragg, I.G., & Dwek, R.A. (1992) Effects of glycosylation on protein conformation and amide proton exchange rates in RNase B. FEBS Lett., 307, 343–346.
28) Sirohi, D., Chen, Z., Sun, L., Klose, T., Pierson, T.C., Rossmann, M.G., & Kuhn, R.J. (2016) The 3.8 Å resolution cryo-EM structure of Zika virus. Science, 352, 467–470.
29) Pokidysheva, E., Zhang, Y., Battisti, A.J., Bator-Kelly, C.M., Chipman, P.R., Xiao, C., Gregorio, G.G., Hendrickson, W.A., Kuhn, R.J., & Rossmann, M.G. (2006) Cryo-EM reconstruction of dengue virus in complex with the carbohydrate recognition domain of DC-SIGN. Cell, 124, 485–493.
30) Gristick, H.B., Wang, H., & Bjorkman, P.J. (2017) X-ray and EM structures of a natively glycosylated HIV-1 envelope trimer. Acta Crystallogr. D Struct. Biol., 73, 822–828.
31) Agirre, J., Davies, G.J., Wilson, K.S., & Cowtan, K.D. (2017) Carbohydrate structure: the rocky road to automation. Curr. Opin. Struct. Biol., 44, 39–47.
32) Frenz, B., Ramisch, S., Borst, A.J., Walls, A.C., Adolf-Bryfogle, J., Schief, W.R., Veesler, D., & DiMaio, F. (2019) Automatically Fixing Errors in Glycoprotein Structures with Rosetta. Structure, 27, 134–139.
33) Agirre, J., Iglesias-Fernandez, J., Rovira, C., Davies, G.J., Wilson, K.S., & Cowtan, K.D. (2015) Privateer: software for the conformational validation of carbohydrate structures. Nat. Struct. Mol. Biol., 22, 833–834.
34) Lutteke, T. & von der Lieth, C.W. (2004) pdb-care (PDB carbohydrate residue check): a program to support annotation of complex carbohydrate structures in PDB files. BMC Bioinformatics, 5, 69.
35) Gristick, H.B., von Boehmer, L., West, A.P. Jr., Schamber, M., Gazumyan, A., Golijanin, J., Seaman, M.S., Fatkenheuer, G., Klein, F., Nussenzweig, M.C., et al. (2016) Natively glycosylated HIV-1 Env structure reveals new mode for antibody recognition of the CD4-binding site. Nat. Struct. Mol. Biol., 23, 906–915.
36) Kuhn, B., Benz, J., Greif, M., Engel, A.M., Sobek, H., & Rudolph, M.G. (2013) The structure of human α-2,6-sialyltransferase reveals the binding mode of complex glycans. Acta Crystallogr. D Biol. Crystallogr., 69, 1826–1838.
37) Tempel, W., Karaveg, K., Liu, Z.J., Rose, J., Wang, B.C., & Moremen, K.W. (2004) Structure of mouse Golgi alpha-mannosidase IA reveals the molecular basis for substrate specificity among class 1 (family 47 glycosylhydrolase) alpha1,2-mannosidases. J. Biol. Chem., 279, 29774–29786.
38) Berman, E., Walters, D.E., & Allerhand, A. (1981) Structure and dynamic behavior of the oligosaccharide side chain of bovine pancreatic ribonuclease B. Application of carbon 13 nuclear magnetic resonance spectroscopy. J. Biol. Chem., 256, 3853–3857.
39) Wyss, D.F., Choi, J.S., Li, J., Knoppers, M.H., Willis, K.J., Arulanandam, A.R., Smolyar, A., Reinherz, E.L., & Wagner, G. (1995) Conformation and function of the N-linked glycan in the adhesion domain of human CD2. Science, 269, 1273–1278.
40) Kato, K. & Yamaguchi, T. (2015) Paramagnetic NMR probes for characterization of the dynamic conformations and interactions of oligosaccharides. Glycoconj. J., 32, 505–513.
41) Usachev, K., Yamaguchi, Y., Takamatsu, M., Pavlova, N., Klochkov, V., Kurbangalieva, A., Murase, T., Shimoda, T., & Tanaka, K. (2017) Simple Gd3+-Neu5NAc complexation results in NMR chemical shift asymmetries of structurally equivalent complex-type N-glycan branches. Analyst (Lond.), 142, 2897–2900.
42) Canales, A., Boos, I., Perkams, L., Karst, L., Luber, T., Karagiannis, T., Dominguez, G., Canada, F.J., Perez-Castells, J., Haussinger, D., et al. (2017) Breaking the Limits in Analyzing Carbohydrate Recognition by NMR Spectroscopy: Resolving Branch-Selective Interaction of a Tetra-Antennary N-Glycan with Lectins. Angew. Chem. Int. Ed. Engl., 56, 14987–14991.
43) Fernandez de Toro, B., Peng, W., Thompson, A.J., Dominguez, G., Canada, F.J., Perez-Castells, J., Paulson, J.C., Jimenez-Barbero, J., & Canales, A. (2018) Avenues to Characterize the Interactions of Extended N-Glycans with Proteins by NMR Spectroscopy: The Influenza Hemagglutinin Case. Angew. Chem. Int. Ed. Engl., 57, 15051–15055.
44) Uzawa, J., Shimabukuro, J., Suzuki, T., Imamura, A., Ishida, H., Ando, H., & Yamaguchi, Y. (2018) J(77Se,1H) and J(77Se,13C) couplings of seleno-carbohydrates obtained by 77Se satellite 1D 13C spectroscopy and 77Se selective HR-HMBC spectroscopy. Magn. Reson. Chem., 56, 836–846.
45) Suzuki, T., Hayashi, C., Komura, N., Tamai, R., Uzawa, J., Ogawa, J., Tanaka, H.N., Imamura, A., Ishida, H., Kiso, M., et al. (2019) Synthesis and Glycan-Protein Interaction Studies of Se-Sialosides by 77Se NMR. Org. Lett., 21, 6393–6396.
46) Rose, A.S., Bradley, A.R., Valasatava, Y., Duarte, J.M., Prlic, A., & Rose, P.W. (2018) NGL viewer: web-based molecular graphics for large complexes. Bioinformatics, 34, 3755–3758.

東北医科薬科大学.博士(薬学).
1993年東京大学薬学部卒業.98年同大学院薬学研究科博士課程修了.98~2001年日本学術振興会特別研究員.01年東京大学薬学部助手.01~07年名古屋市立大学大学院薬学研究科講師.07~18年理化学研究所チームリーダー.19年同研究所研究員.19年4月より現職.
研究テーマと抱負構造に不均一性(曖昧さ)をもつ糖鎖が時に厳密に制御され機能している,そのギャップに生命の不思議さを感じつつ,糖鎖の機能を一つでも多く明らかにし,得られた知識を医薬品開発につなげることができればこの上ない喜びです.
趣味習い始めの合気道,年に1, 2回のゴルフ,車の運転練習を兼ねた日帰り温泉の旅.

This page was created on 2020-04-22T15:22:45.201+09:00
This page was last modified on 2020-05-28T08:26:55.000+09:00
このサイトは(株)国際文献社によって運用されています。