糖鎖合成の新展開New development of oligosaccharide synthesis
大阪大学大学院理学研究科化学専攻Department of Chemistry, Graduate School of Science, Osaka University ◇ 〒560–0043 大阪府豊中市待兼山町1–1 ◇ 1–1 Machikaneyama, Toyonaka, Osaka 560–0043
発行日:2020年6月25日Published: June 25, 2020
糖鎖機能解明のために必要な糖鎖は,化学合成により調製されてきた.糖鎖合成は,糖と糖を連結するグリコシル化反応を繰り返し,単糖から糖鎖へと構築する.この手法を駆使することで,分枝構造や,アミノ酸に結合したさまざまな糖鎖を合成することができる.
また,糖鎖は,糖転移酵素と糖ヌクレオチドを用いる酵素反応を利用することで合成できる.しかし,糖鎖が持つすべての結合様式に相当する糖転移酵素を用意するのは困難なので,化学合成で一部糖鎖骨格を構築し,その他を酵素で合成する酵素化学法が発展した.また,鶏卵から単離したヒト複合型二分枝糖鎖を出発原料に,市販のグリコシダーゼや,糖転移酵素を利用することで,多様な構造の糖鎖を合成することもできる.本稿ではこれらを詳細に紹介する.
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
細胞表層,血液中のタンパク質表層には,糖が連なった糖鎖が存在する.生体内の糖鎖は,グルコース,マンノース,ガラクトース,N-アセチルグルコサミン,フコース,N-アセチルシアル酸などが連結したものであり,さまざまな結合様式でつながることで糖タンパク質型,糖脂質型,さらには硫酸化糖鎖に分類される.
タンパク質に結合した糖鎖は,結合部位であるアミノ酸側鎖の種類によってN型,O型の二つに分けられる.N型糖鎖は,Asn-X-Ser/Thr(Xはプロリン以外のアミノ酸)のアスパラギン側鎖の窒素原子に糖鎖がN-グリコシル化されている1).N型糖鎖は,3種類に分けることができ,複合型1,ハイマンノース型3,その両方の糖鎖様式を持つハイブリッド型2が存在する(図1).ハイマンノース型糖鎖は,マンノースを九つ持ち,複合型は,末端からシアル酸,ガラクトース,N-アセチルグルコサミンからなり,さらに,二,三,四分枝と多様な構造を示す2).糖脂質は,ゴルジ体で主に合成されるが,シアル酸,グルコース,ガラクトース,N-アセチルガラクトサミン,フコースなどで構成される.
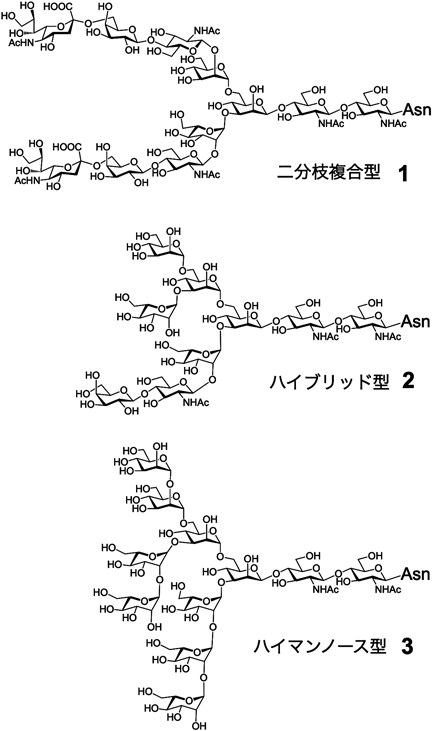
タンパク質上のハイマンノース型糖鎖は,小胞体内でのタンパク質フォールディングを促進する機能を有している3).小胞体で,リボソームによりペプチドが合成される際,グルコース三つ,マンノース九つからなる糖鎖(糖鎖3にグルコースが三つ付加)がオリゴ糖転移酵素により付加する.そして,この糖鎖にさまざまなシャペロンやフォールディングセンサー酵素が相互作用することでタンパク質のフォールディングが促進される.このハイマンノース型糖鎖3は,ゴルジ体で一部,酸性のシアリル糖鎖へと変換され,複合型糖鎖1となり,細胞表層,体液中でのタンパク質の構造安定性,溶解性,寿命,抗原性に関与するようになる.ゴルジ体では,マンノシダーゼ,糖転移酵素が作用し,糖鎖末端からシアル酸,ガラクトース,N-アセチルグルコサミンからなる糖鎖が構築される.このシアル酸,ガラクトース,N-アセチルグルコサミンからなる糖鎖が2組の場合は,二分枝N型糖鎖となる.この分枝様式は三,四分枝も存在する2).
生合成における糖鎖配列の制御は,遺伝子制御ではなく糖転移酵素の基質特異性により行われ,また修復機能もないため構造が不均一となる.また,N型糖鎖は,前述のように二~四分枝となるため,さらに構造多様性が生じ,どの構造の糖鎖が機能を発揮しているか確定することが困難となる.
生物科学的な実験を行い糖鎖機能を理解するには糖鎖試料が必要であるが,生体試料から単離するには微量のため,さまざまな実験に用いることができない.一般には,研究対象として興味深いタンパク質を単離し,その後糖鎖を酵素で切り離すことで,その糖鎖を質量分析等で構造解析することができる.特に,抗体をはじめ比較的量が確保できるタンパク質(血中濃度が5~10 pg/mL以上)4)の糖鎖は,単離後,核磁気共鳴装置,質量分析装置等で構造決定することができると同時に,次の生物科学的実験に利用することができる.しかし,興味深い活性を示すサイトカインなどは,0.5 pg/mL程度しか血中には存在しておらず4),そこから糖鎖を単離しても前述の糖鎖の不均一性という問題が重なり,質量分析装置でもその構造解析は困難を極める.このような場合は,チャイニーズハムスターの卵巣細胞(CHO細胞)等で糖タンパク質を大量に発現し,そこから糖鎖構造を決定することで糖鎖と活性の関係の研究を実施することができる.しかし,天然型から単離した糖タンパク質とCHO細胞で調製した糖タンパク質の糖鎖組成が同じとはいえない問題もある.
以上の理由から糖鎖を化学的に合成する必要性があり,また,高純度のものを得ることができれば生物科学的実験に活用することができる.
糖鎖を化学合成するには,糖と糖を連結するグリコシル化反応を繰り返し,単糖から糖鎖へと構築する(図2).このグリコシル化反応の研究は1893年にFischer5)が実施し,その後100年を経て数多くのすばらしい反応が見つかってきた.また,酵素を有機試薬のように利用した糖鎖合成も活用されるようになってきた.以下ではその糖鎖合成について経緯を一部ではあるが紹介する.
糖は,アルデヒドと水酸基が分子内アセタールを形成した構造で,そのアセタール炭素位をアノメリックといい,ここに陽電荷(カチオン)5を化学的に発生させ,別の糖水酸基に結合させることをグリコシル化という(図2).このカチオンをアノメリック位に発生させるためには,チオグリコシド,イミダート基,ハロゲン(フッ素,塩素,臭素)などの官能基がアノメリックに結合した糖供与体(ドナー)4を調製する必要がある.そして,それぞれ該当する活性化法で,この官能基を脱離させることでカチオン5を発生させる.この5はオキソカルベニウムイオン6と共鳴構造をとる.この際,反応に用いる溶液中に水がわずかにでも存在するとこのカチオンと即座に反応するため,目的とするグリコシル化反応は起こらない.この不要な水分を除去するために,原料の乾燥は必要不可欠である.さらに水を強力に吸着するモレキュラーシーブスを反応溶液中に加えてアルコールと反応させることで,グリコシド体7(β結合)または8(α結合)を得ることができる.α選択的にグリコシル化反応を制御する方法はいまだ確立されていないが,基質や条件によって制御することが可能である.一方,β選択的グリコシル化反応は,糖の2位にエステル系保護基(11)を導入しておくことで,12のようなアシロキソニウムイオン中間体を経てβ選択的にグリコシド体13を得ることが可能である.
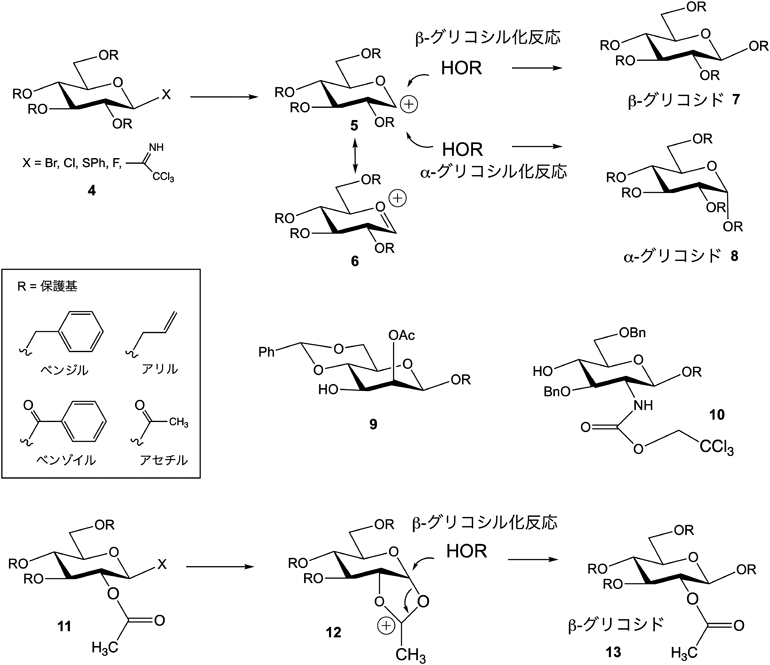
一方,カチオンを反応させる他方の糖の水酸基は複数個あるので,決まった位置の水酸基と反応させるには,保護基を利用する必要がある.糖は,2, 3, 4, 6位に水酸基を持つが,その1か所をグリコシル化するため,それ以外の水酸基にエステル系,エーテル系の保護基を導入する[たとえば9(3位のみが水酸基),または10(4位のみが水酸基)].エステル系保護基は糖鎖合成の最終工程で塩基によるけん化により,ベンジル,アリル基などのエーテル系は触媒を利用する反応により除去し,水酸基が遊離した天然型糖鎖へと変換することができる.
また,糖鎖を合成するには,保護基の脱着と糖をグリコシル化させるタイミングを綿密に計画する必要がある.糖鎖合成は各合成ステップごとにシリカゲルや順相のHPLCカラムで精製することが望ましい.特に糖鎖全長が合成できた際は,有機溶媒に可溶なため比較的精製が容易なので,この段階で行い,あとは定量的に保護基が脱保護できる条件を探すことで最終生成物を高純度で得られるようにする必要がある.
糖鎖の生合成は,糖転移酵素と糖ヌクレオチドが働くことで行われており,この反応をフラスコ内で行うことで糖鎖を合成することができる.糖転移酵素は,糖ヌクレオチドを糖供与体として用い,該当する糖受容体に糖を転移することで糖鎖を構築する.生体内糖鎖は,さまざまな結合様式を示し,グリコシド結合のα/β配置,ならびに受容体の種類ごとに異なる糖転移酵素が働く.したがって,酵素を用いて糖鎖を合成する場合,糖鎖が持つグリコシド結合の種類だけ糖転移酵素が必要となる.これまで単離された糖鎖関連酵素はCAZYというウエブサイトで報告されている(http://www.cazy.org).
糖鎖を酵素合成するには,糖鎖が持つすべての結合様式に相当する糖転移酵素を用意する必要があるが,現実的には困難である.この場合は,化学的グリコシル化と糖転移酵素反応を組み合わせて実施することが効率的である.すなわち,基本となる糖鎖骨格は化学合成で調製し,そして非還元末端側に酵素で糖鎖を伸長するのが一般的である.
また,鶏卵から単離した複合型二分枝糖鎖を出発原料に,市販のグリコシダーゼや,糖転移酵素を利用することで,多様な構造の糖鎖を合成することもできる.この方法で30種類以上の複合型糖鎖を容易に得ることができるようになっている6, 7).最近では,同上の複合型二分枝糖鎖の各水酸基を選択的に保護し,化学的グリコシル化反応により三分枝糖鎖の合成も報告されている8).
ここからは,これまで報告された糖鎖合成の代表的なものを紹介する.
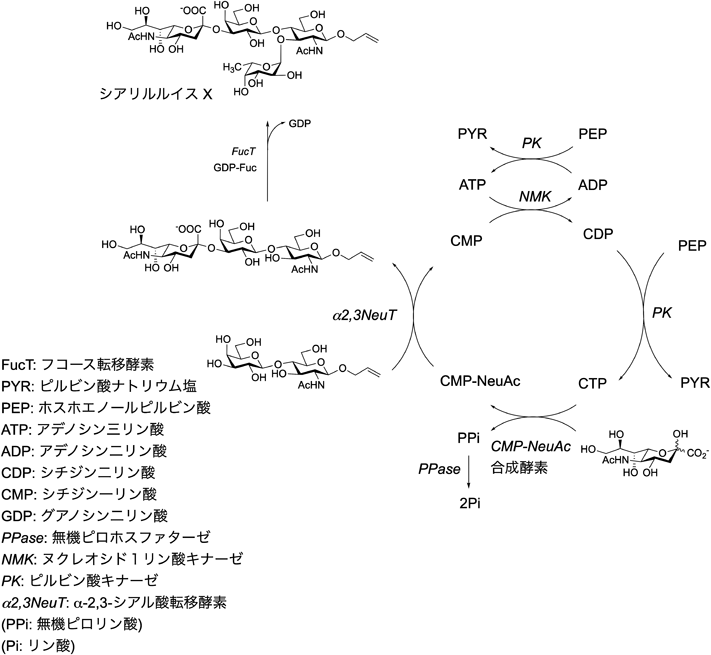
糖転移酵素を用いた糖鎖合成を系統的に実施したのはWongらのグループで,一つのフラスコ内にいくつもの酵素を入れて,生合成システムのような手法で抗炎症剤として期待されたシアリルルイスXの合成に成功した(図3)9).この方法は,糖ヌクレオチドとしてUDP-ガラクトース,CMP-シアル酸,GDP-フコースを複数の酵素で合成する過程と糖転移の過程を合わせている.ここでは,糖ヌクレオチドをリサイクルで合成する方法が利用されているが,そのメリットは,糖転移によって生じたヌクレオチド(CMP, UDP)がそれぞれシアル酸転移酵素,ガラクトース転移酵素の酵素阻害剤として作用することを抑え,反応が失速しないようにできることである.彼らは,CMP-シアル酸の再生法を記載している(図3右).その結果,UDP, CMPは触媒量で十分であり,該当する糖ヌクレオチドに再生するための酵素反応システムが取り入れられている.また,糖ヌクレオチドは高価なため,触媒量の糖ヌクレオチドでシアリルルイスXが合成できるメリットは非常に大きい.
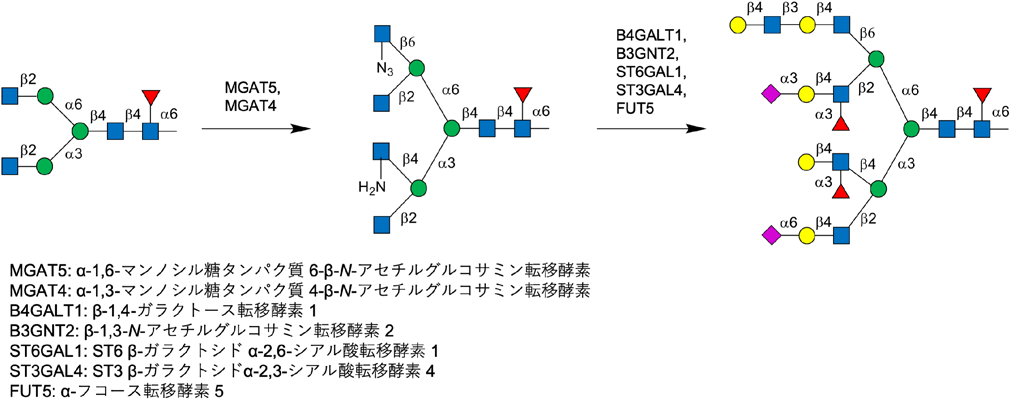
糖転移酵素は本来膜結合型酵素のため大腸菌での発現が困難であったが,さまざまな発現系が検討され,多くの糖転移酵素の膜外に位置する触媒ドメインの発現ができるようになるとともに,それを用いた糖鎖合成も発展した.しかし,すべての糖鎖結合様式を合成できるほどの糖転移酵素を用意することはできないので,基本骨格は化学合成で調製しておき,その非還元末端側の糖鎖を糖転移酵素で伸長するのが常套手段となっている.この手法を用いることで,最近,N型の多分枝糖鎖の誘導体が合成されている.Boonsらは,基本骨格となる糖鎖は化学的に合成し,そして分枝特異的に糖転移酵素を用いて糖鎖伸長することで分枝様式が非対称のN型糖鎖誘導体の合成に成功している(図4)10, 11).
シアル酸は,細胞ががん化した際に増加する糖鎖として注目され,その機能解明のため高純度シアリル糖鎖の合成は長く研究が続けられてきたが,その高効率的な立体選択的な合成法が最近確立された.シアル酸は,そのアノメリック位が2位となるケトース構造で,天然型はα型(図5: α-シアロシド)となる.しかし,熱学力的に不安定なα型のグリコシル化反応を実施するのは非常に困難で,これまでさまざまな検討がされてきた.そのような中,世界で最初にα選択的なシアリル化反応に成功したのは小川らのグループで,ガングリオシドGM3の合成を報告した12).また,3位に硫黄原子を付加させ反応中間体15をα型が生成しやすいものにする方法や,アセトニトリルを用いた溶媒効果によるα選択的な制御(17)など多くの例が報告された(図5)13).最近,田中らによる4, 5位間に環状構造のオキサゾリジノンを形成したシアリル供与体(18)でシアル酸ポリマーの合成が高効率的にできるようになっている14).
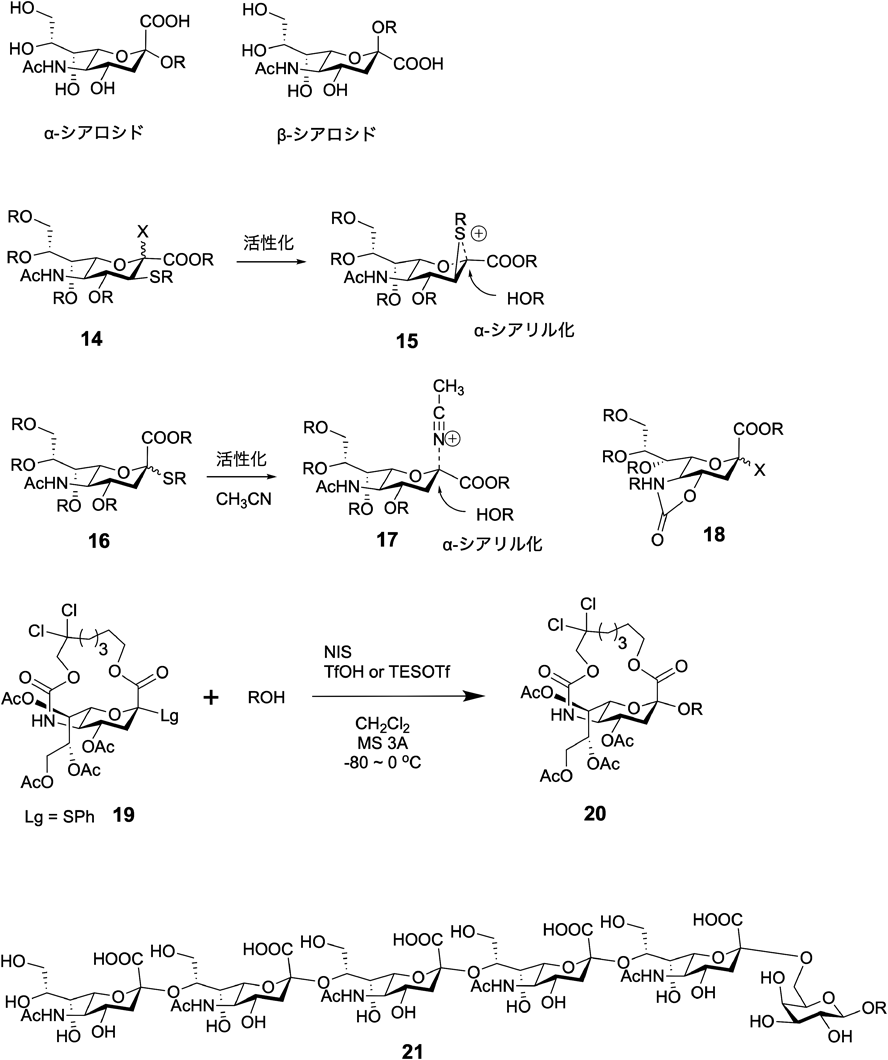
また,河村,安藤らは,図5に示したような環状のシアル酸供与体19を合成し,これがさまざまなシアル酸受容体と反応しα選択的なシアリル糖鎖20やシアル酸ポリマー21を与えることを示した15).この方法の特徴は,望まない立体であるβ体が生成する方向に環状の炭素鎖骨格を付加させることで,シアリル化反応の際,β体の生成は起こらずα体のみ与えることができる.また,その環状構造を形成する炭素鎖の長さは,2位と糖受容体水酸基が反応する際の分子全体の立体構造の安定性,ならびに,反応後に環状構造の炭素鎖の除去法などさまざまな条件検討が行われ確立された.
N型糖鎖については,複合型1,ハイマンノース型3の糖鎖の化学合成例が報告されている.松尾,伊藤らは,マンノース単糖を繰り返し連結することで九つのマンノースがキトビオースにつながった糖鎖を合成している16).この合成で最も困難なグリコシル化反応は,マンノースがN-アセチルグルコサミンの4位に結合したβ1,4結合である.この結合は,伊藤らにより開発された立体選択的にβ1,4結合が形成できる分子内糖転移反応によって合成されている17, 18).
伊藤らは二分枝の複合型糖鎖19),Danishefskyらは三分枝の化学合成20)を報告している.Danishefskyらの三分枝複合型糖鎖の合成では,まず,アスパラギンに結合するキトビオースにCrichらが報告したベンジリデンで構造を固定化したマンノシル供与体によるβ1,4結合形成を行っている(図6: 26→27).そして,そのβマンノースの3, 6位にマンノースをα結合で連結している(図6:27→33).最後に,シアル酸α2-3ガラクトースβ1-4グルコサミンからなる三糖骨格を反応性が最も低い,マンノースの4位にグリコシル化し,そして,マンノースの2位に二,三分枝目に相当するシアル酸α2-3ガラクトースβ1-4グルコサミン骨格を導入している(36).この合成では,ベンジル基というエーテル系の保護基を用い,糖供与体の反応性を向上させるようなルートが設定されているが,最後の脱保護は,液体アンモニア中ナトリウムによる還元反応により行われている.
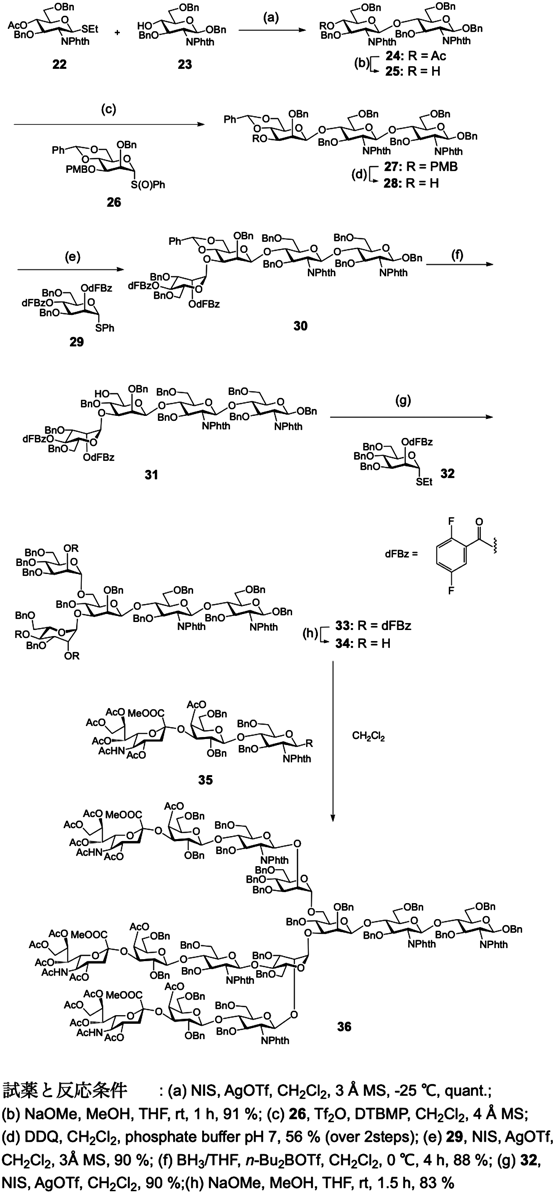
非天然型の糖を有する複合型糖鎖を合成することによって,その糖鎖機能や,その糖鎖が結合しているタンパク質の機能を制御することが可能となる.
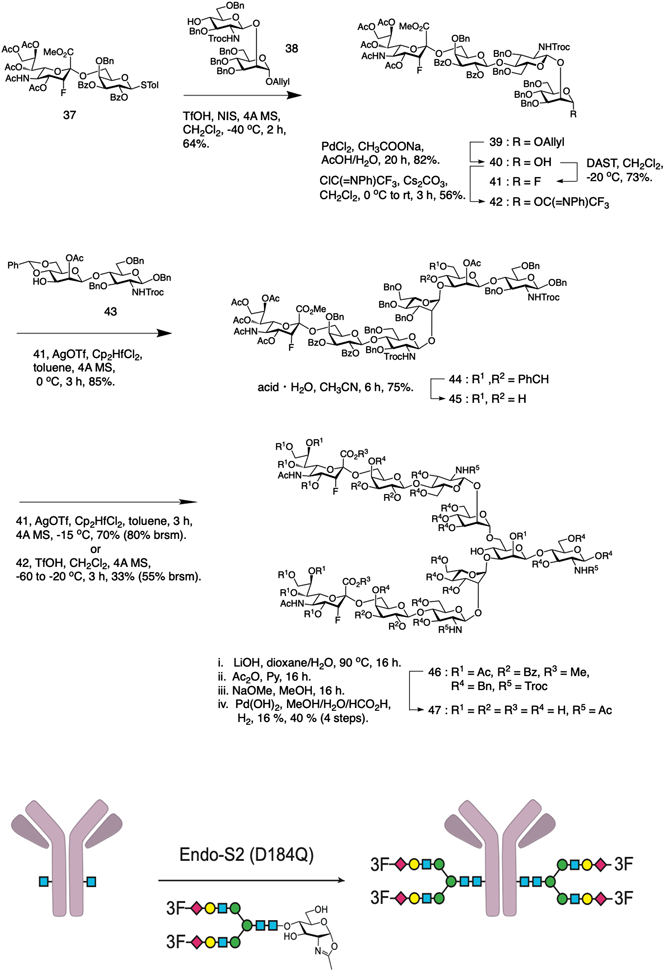
Wongらは二分枝複合型糖鎖の非還元末端のシアル酸の3位にフッ素原子を結合させた糖鎖47を化学合成後,その糖鎖を酵素を用いてヒトIgGのFc部位に導入している(図7)21).これは,シアリル糖鎖を持つ抗体がより高い機能を持つため,抗体の糖鎖をシアリル糖鎖に置換した高機能抗体の作製を目的としている.しかし,シアル酸は,生体内でシアリダーゼにより除去されるため,Wongらは,電子吸引性のフッ素原子によりグリコシド結合を安定化させ,シアル酸がシアリダーゼにより除去されないようにした.フッ素原子を持つシアリル糖鎖の合成は,前述のDanishefskyらと同様に行っているが,フッ素原子をシアル酸に導入する工程は,シアリルガラクトースを合成後に導入している(図7:37).これは,電気陰性度が高いフッ素原子の導入により,ガラクトースのシアリル化反応が極端に低下するためである.そして,シアリル三糖供与体42を合成後,コアマンノースの反応性の低い3位水酸基(44),そして6位水酸基(46)という順にシアリル糖鎖を合成した.次に還元末端のN-アセチルグルコサミンをオキサゾリン化し,Wongらが調製したエンドグリコシダーゼEndo-S2(D184Q)での糖鎖転移反応を行い,フッ素原子を持つシアリル糖鎖を持つ抗体の作製に成功した21).
シアル酸は,その分子内に酸性のカルボキシ基を持つことで特異な性質を持つと考えられるが,生体内糖鎖には,硫酸化された酸性糖鎖が存在する.硫酸化様式,構成糖の種類によりコンドロイチン硫酸,ヘパラン硫酸,デルマタン硫酸などが見いだされている.いずれも細胞表層でさまざまなシグナルを発信しているが,その生理活性は,硫酸化様式によって大きく変化する.その結果,これまで多くの硫酸化糖が化学合成され,機能解明の研究が展開されてきた.
しかし,糖鎖への硫酸化も酵素により制御されており,その硫酸化様式は不均一な構造を示す.したがって硫酸化糖の機能を解明するには,化学的に合成した均一な試料を利用することが必要不可欠である.硫酸化糖鎖の合成は,まず,基本となる骨格,たとえば,二糖と二糖を連結し四糖,さらには,六,八糖を合成し,そして硫酸化する位置の保護基を除去し,硫酸化する方法が一般的である.また,硫酸化は,三酸化硫黄のピリジン塩が一般に用いられるが,その反応性が低いことは長年の課題となっている.田村22, 23),隅田24),Hung25, 26),Huang27),Boons28),Hsieh-Wilson29)らによってさまざまな硫酸化様式の糖鎖が合成され,硫酸化様式と活性発現の関係が調べられた.また,田村30),Huang27)らは,保護基で保護した硫酸基を糖供与体に導入しておき,糖と糖の連結を行い,硫酸化糖鎖が合成できることも示している.
コンドロイチン硫酸は,半合成によってそのライブラリーの合成が可能であることが報告されている.この方法では,コンドロイチン硫酸から酸処理による硫酸基の除去を行い,ヒアルロニダーゼで二および四糖骨格を調製している.そして,これら糖鎖に対し保護基を導入することで硫酸基を導入する位置を制御し,さまざまな位置に硫酸基が導入されたコンドロイチン硫酸が合成されている(図8)31).
グルコースが環状につながった糖鎖はシクロデキストリンと呼ばれ,その環状内部への包摂作用によりさまざまな応用が展開されてきた.シクロデキストリンの化学合成は小川ら32)が世界で最初に達成し,そして合成が難しいと思われていたグルコース五つからなるシクロデキストリンの合成が中川ら33)によって達成された.

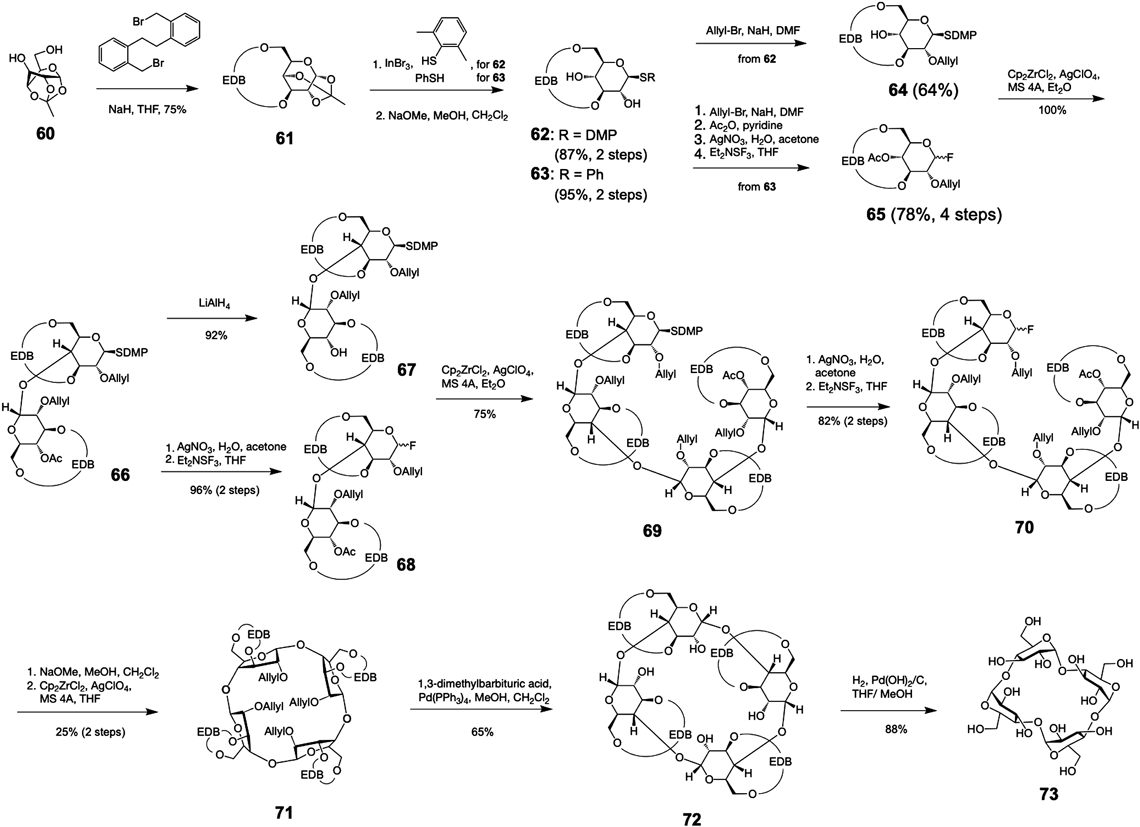
そして,2019年には,合成は不可能と思われていた,グルコースが三つ,あるいは四つからなるシクロデキストリンが山田英俊らによって合成ができることが証明された34).山田らは,グルコースのピラノース環に彼ら独自の保護基(EDB基)を導入することで,環状の保護基を持つグルコース誘導体61を合成した.この構造を構築することでグルコースのピラノース環がゆがんだ構造となり,グリコシル化反応の高い立体選択性を示すグルコース二糖供与体68を得た.そして,これら二糖を連結して四糖骨格を構築後(図9: 67, 68→69),分子内グリコシル化反応を実施し目的とする環状四糖,すなわち四糖シクロデキストリンの合成に成功した(図9:71→73).また,山田らは,この手法を用いて,グルコース三つからなるシクロデキストリンの合成にも成功した.これら成果は,Science誌で,難しい化学結合も環状構造をゆがませることで合成が可能になったと高く評価され,歴史に残る成果を糖質化学に記した34).
糖鎖合成は,多くの合成化学者により発展し,今では作ることができない糖鎖はないといっても過言ではない.以上のようなさまざまな化学的合成法が発展し,最近では,細菌表層の大型糖鎖をも合成できるようになっている.この糖鎖は,40残基以上の五員環のフラノース骨格を多く含む特異な構造をしている.
このフラノース骨格の細菌の糖鎖は,石渡,伊藤ら35)によって行われ,また最近,Yeらによって最も長い糖鎖の合成例が報告された36).これらの合成では,糖鎖連結時にフラノースが持つ1位のα選択性は,2位に結合させたエステル系保護基によって制御されている(図10).
これまで述べたような合成糖鎖は,マイクロアレイに固定化し糖鎖チップとして,生体内のさまざまな受容体タンパク質などとの相互作用を調べる実験に利用することができる.また,最近では,これら糖鎖を自動合成装置で合成する試みも展開され,装置の市販品も出てきている37).
糖鎖合成が自在にできるようになった現在,今後の糖鎖研究において検討するべき課題は,ある糖鎖分子を鋳型とする糖鎖の増幅,数百個以下の分子しかない極微量の糖鎖の構造解析法があげられる.糖鎖は,DNAやタンパク質のように鋳型を用いて生合成されるわけではないので,生体から単離した微量の糖鎖をポリメラーゼ連鎖反応のように,その分子数を増幅することができない.また,対象となる糖タンパク質は血中濃度が0.5 pg/mLと非常に微量で,得られた微量の活性糖タンパク質から糖鎖を切り出し,その構造を質量分析装置等で構造解析することはいまだ容易ではない.そのため,現状ではある程度量が確保できる糖鎖の構造解析のみ可能である.今後,有用な活性を示す極微量糖鎖の構造解析38)が可能になれば,その糖鎖を化学合成により大量に調製し,さまざまな応用研究や創薬に発展することが期待される.
1) Stanley, P., Schachter, H., & Taniguchi, N. (2009) N-glycans, in Essential Glycobiology (Varki, A. et al. eds), pp. 101–114, Cold Spring Harbor Laboratory Press.
2) Bertozzi, C.R. & Rabuka, D.(2009) Structural basis of glycan diversity, in Essential Glycobiology (Varki, A. et al. eds), pp. 23–26, Cold Spring Harbor Laboratory Press.
3) D’Alessio, C., Caramelo, J.J., & Parodi, A.J. (2010) UDP-GlC: Glycoprotein glucosyltransferase-glucosidase II, the ying-yang of the ER quality control. Semin. Cell Dev. Biol., 21, 491–499.
4) Anderson, N.L. & Anderson, N.G. (2002) The human plasma proteome: History, character, and diagnostic prospects. Mol. Cell. Proteomics, 1, 845–867.
5) E. Fischer (1893) Ueber die Gluooside der Alkohole. Ber. Dtsch. Chem. Ges., 26, 2400–2412.
6) Kajihara, Y., Suzuki, Y., Yamamoto, N., Sasaki, K., Sakakibara, T., & Juneja, L.R. (2004) Prompt chemoenzymatic synthesis of diverse complex-type oligosaccharides and its application to the solid-phase synthesis of a glycopeptide with Asn-linked sialyl-undeca- and asialo-nonasaccharides. Chemistry, 10, 971–985.
7) Fukae, K., Yamamoto, N., Hatakeyama, Y., & Kajihara, Y. (2004) Chemoenzymatic synthesis of diverse asparagine-linked alpha-(2,3)-sialyloligosaccharides. Glycoconj. J., 21, 243–250.
8) Maki, Y., Okamoto, R., Izumi, M., Murase, T., & Kajihara, Y. (2016) Semisynthesis of intact complex-type triantennary oligosaccharides from a biantennary oligosaccharide isolated from a natural source by selective chemical and enzymatic glycosylation. J. Am. Chem. Soc., 138, 3461–3468.
9) Ichikawa, Y., Lin, Y.C., Dumas, D.P., Shen, G.J., Garcia-Junceda, E., Williams, M.A., Bayer, R., Ketcham, C., Walker, L.E., Paulson, J.C., et al. (1992) Chemical-enzymic synthesis and conformational analysis of sialyl Lewis X and derivatives. J. Am. Chem. Soc., 114, 9283–9298.
10) Liu, L., Prudden, A. R., Capicciotti, C. J., Bosman, G. P., Yang, J.-Y., Chapla, D. G., Moremen, K. W., & Boons, G.-J. ((2019) Streamlining the chemoenzymatic synthesis of complex N-glycans by a stop and go strategy. Nat. Chem., 11, 161–169.
11) Wang, Z., Chinoy, Z.S., Ambre, S.G., Peng, W., McBride, R., de Vries, R.P., Glushka, J., Paulson, J.C., & Boons, G.-J. (2013) A general strategy for the chemoenzymatic synthesis of asymmetrically branched N-glycans. Science, 341, 379–383.
12) Ogawa, T. & Sugimoto, M. (1985) Synthesis of α-Neu5Acp-(2→3)-D-Gal and α-Neu5Acp-(2→3)-β-D-Galp-(1→4)-D-Glc. Carbohydr. Res., 135, C5–C9.
13) Kanie, O., Kiso, M., & Hasegawa, A. (1988) Glycosylation using methylthioglycosides of N-acetylneuraminic acid and dimethyl(methylthio)sulfonium triflate. J. Carbohydr. Chem., 7, 501–506.
14) Tanaka, H., Nishiura, Y., & Takahashi, T. (2009) Stereoselective Synthesis of alpha(2,9) di- to tetrasialic acids, using a 5,4-N,O-carbonyl protected thiosialoside. J. Org. Chem., 74, 4383–4386.
15) Komura, N., Kato, K., Udagawa, T., Asano, S., Tanaka, H.-N., Imamura, A., Ishida, H., Kiso, M., & Ando, H. (2019) Constrained sialic acid donors enable selective synthesis of α-glycosides. Science, 364, 677–680.
16) Matsuo, I., Wada, M., Manabe, S., Yamaguchi, Y., Otake, K., Kato, K., & Ito, Y. (2003) Synthesis of monoglucosylated high-mannose-type dodecasaccharide, a putative ligand for molecular chaperone, calnexin, and calreticulin. J. Am. Chem. Soc., 125, 3402–3403.
17) Ito, Y. & Ogawa, T. (1994) A novel approach to the stereoselective synthesis of β-mannosides. Angew. Chem. Int. Ed. Engl., 33, 1765–1767.
18) Ito, Y. & Ogawa, T. (1997) Intramolecular aglycon delivery on polymer support: Gatekeeper monitored glycosylation. J. Am. Chem. Soc., 119, 5562–5566.
19) Seifert, J., Lergenmuller, M., & Ito, Y. (2000) Synthesis of anα-(2,3)-sialylated, complex-type undecasaccharide. Angew. Chem. Int. Ed., 39, 531–534.
20) Walczak, M.A. & Danishefsky, S. (2012) Solving the convergence problem in the synthesis of triantennary N-glycan relevant to prostate-specific membrane antigen (PSMA). J. Am. Chem. Soc., 134, 16430–16433.
21) Lo, H.-J., Krasnova, L., Dey, S., Cheng, T., Liu, H., Tsai, T.-I., Wu, K.B., Wu, C.-Y., & Wong, C.-H. (2019) Synthesis of sialidase-resistant oligosaccharide and antibody glycoform containing α2,6-linked 3Fax-Neu5Ac. J. Am. Chem. Soc., 141, 6484–6488.
22) Matsushita, K., Nakata, T., Takeda-Okuda, N., Nadanaka, S., Kitagawa, H., & Tamura, J.-i. (2018) Synthesis of chondroitin sulfate CC and DD tetrasaccharides and interactions with 2H6 and LY111. Bioorg. Med. Chem., 26, 1016–1025.
23) Takeda-Okuda, N., Yamaguchi, Y., Uzawa, J., & Tamura, J.-i. (2017) Synthesis of a biotinylated keratan sulfate tetrasaccharide composed of dimeric Galβ1-4GlcNAc6Sβ. Carbohydr. Res., 452, 97–107.
24) Miyachi, K., Wakao, M., & Suda, Y. (2015) Syntheses of chondroitin sulfate tetrasaccharide structures containing 4,6-disulfate patterns and analysis of their interaction with glycosaminoglycan-binding protein. Bioorg. Med. Chem. Lett., 25, 1552–1555.
25) Sakamoto, K., Ozaki, T., Ko, Y.-C., Tsai, C.-F., Gong, Y., Morozumi, M., Ishikawa, Y., Uchimura, K., Nadanaka, S., Kitagawa, H., et al. (2019) Glycan sulfation patterns define autophagy flux at axon tip via PTPRσ-cortactin axis. Nat. Chem. Biol., 15, 699–709.
26) Zulueta, M.M.L., Chyan, C.-L., & Hung, S.-C. (2018) Structural analysis of synthetic heparan sulfate oligosaccharides with fibroblast growth factors and heparin-binding hemagglutinin. Curr. Opin. Struct. Biol., 50, 126–133.
27) Tiruchinapally, G., Yin, Z., El-Dakdouki, M., Wang, Z., & Huang, X. (2011) Divergent heparin oligosaccharide synthesis with preinstalled sulfate esters. Chemistry, 17, 10106–10112.
28) Lu, W., Zong, C., Chopra, P., Pepi, L.E., Xu, Y., Amster, I.J., Liu, J., & Boons, G.-J. (2018) Controlled chemoenzymatic synthesis of heparan sulfate oligosaccharides. Angew. Chem. Int. Ed., 57, 5340–5344.
29) Gama, C.I., Tully, S.E., Sotogaku, N., Clark, P.M., Rawat, M., Vaidehi, N., Goddard, W.A. 3rd, Nishi, A., & Hsieh-Wilson, L.C. (2006) Sulfation patterns of glycosaminoglycans encode molecular recognition and activity. Nat. Chem. Biol., 2, 467–473.
30) Matsushita, K., Nakata, T., & Tamura, J.-i. (2015) The application of 2,2,2-trichloroethyl sulfate to the synthesis of chondroitin sulfate C and D. Carbohydr. Res., 406, 76–85.
31) Xiao, Z., Huiying, L., Wang, Y., Xiangbao, M., & Zhongjun, L. (2019). J. Org. Chem., 84, 7418–7425.
32) Mori, M., Ito, Y., & Ogawa, T. (1989) A highly efficient and stereoselective cycloglycosylation. Synthesis of cyclo{→4)-[α-Man-(1→4)]5-α-Man-(1→} a manno isomer of α-cyclodextrin. Tetrahedron Lett., 30, 1273–1276.
33) Nakagawa, T., Ueno, K., Kashiwa, M., & Watanabe, J. (1994) The stereoselective synthesis of cyclomaltopentaose. A novel cyclodextrin homologue with D.P. five. Tetrahedron Lett., 35, 1921–1924.
34) Ikuta, D., Hirata, Y., Wakamori, S., Shimada, H., Tomabechi, Y., Kawasaki, Y., Ikeuchi, K., Hagimori, T., Matsumoto, S., & Yamada, H. (2019) Conformationally supple glucose monomers enable synthesis of the smallest cyclodextrins. Science, 364, 674–677.
35) Ishiwata, A. & Ito, Y. (2011) Synthesis of docosasaccharide arabinan motif of mycobacterial cell wall. J. Am. Chem. Soc., 133, 2275–2291.
36) Wu, Y., Xiong, D.-C., Chen, S.-C., Wang, Y.-S., & Ye, X.-S. (2017) Total synthesis of mycobacterial arabinogalactan containing 92 monosaccharide units. Nat. Commun., 8, 14851.
37) Hahm, H.S., Schlegel, M.K., Hurevich, M., Eller, S., Schuhmacher, F., Hofmann, J., Pagel, K., & Seeberger, P.H. (2017) Automated glycan assembly using the Glyconeer 2.1 synthesizer. Proc. Natl. Acad. Sci. USA, 114, E3385–E3389.
38) Gray, C.J., Migas, L.G., Barran, P.E., Pagel, K., Seeberger, P.H., Eyers, C.E., Boons, G.-J., Pohl, N.L.B., Compagnon, I., Widmalm, G., et al. (2019) L. J. Am. Chem. Soc., 141, 14463–14479.

大阪大学大学院理学研究科化学専攻教授.博士(理学).
1988年3月神奈川大学工学部応用化学科卒業.90年3月同大学院工学研究科応用化学修了(指導教官:佐藤憲一教授,天然物合成:テトロドトキシン).93年3月東京工業大学大学院総合理工学研究科博士後期課程生命化学専攻修了(指導教官:橋本弘信教授,酵素阻害剤合成,糖鎖の酵素化学合成).同年4月日本たばこ産業株式会社生命科学研究所博士研究員(研究内容:糖ヌクレオチドの化学合成と糖鎖の酵素合成).95年6月横浜市立大学理学部助手.2001年4月同大学院総合理学研究科助教授(研究内容:糖タンパク質の化学合成現在に至).05年4月同大学院国際総合科学研究科准教授.07年4月同教授.09年4月大阪大学大学院理学研究科教授.
研究テーマと抱負糖鎖機能を解明するための,糖タンパク質精密化学合成.化学合成した糖タンパク質を細胞表層に導入できるよう検討中.
ウェブサイトhttp://www.chem.sci.osaka-u.ac.jp/lab/kajihara/index.html
趣味釣り(磯),テニス.

This page was created on 2020-04-21T11:00:01.326+09:00
This page was last modified on 2020-06-04T14:01:34.000+09:00
このサイトは(株)国際文献社によって運用されています。