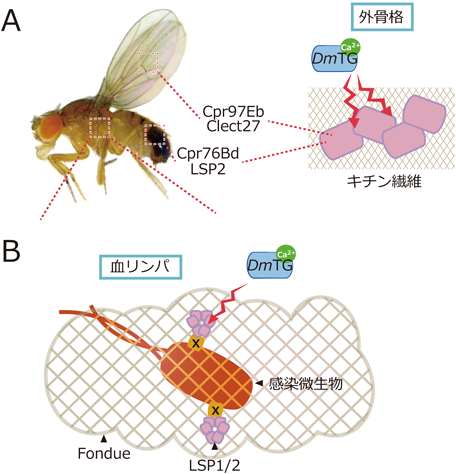
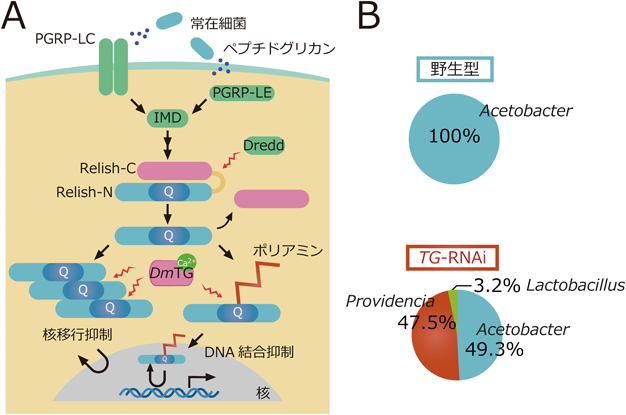
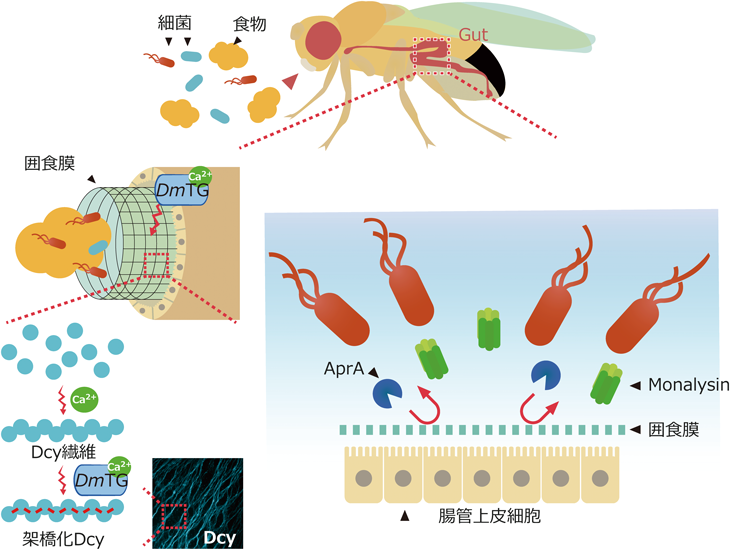
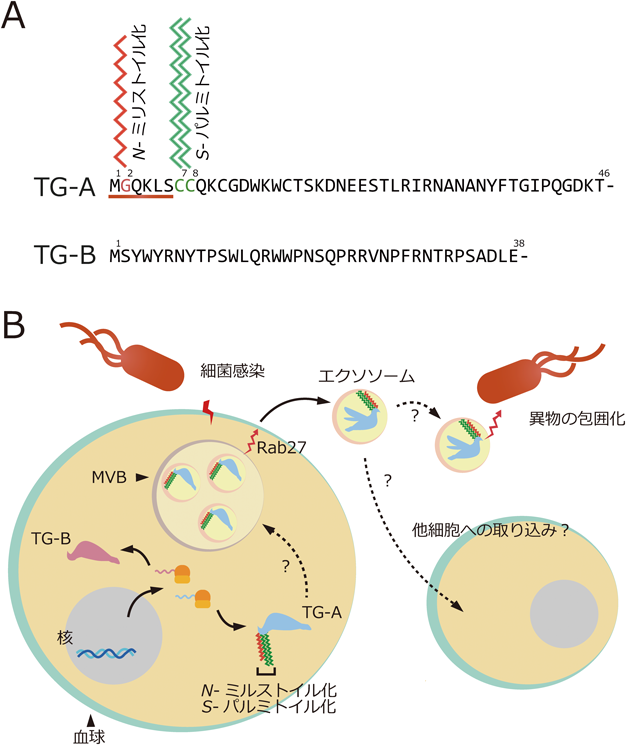
キイロショウジョウバエを用いたトランスグルタミナーゼの生理機能と分泌機構の解析Functional analysis and a secretion mechanism of Drosophila Transglutaminase
九州大学大学院理学研究院生物科学部門生体高分子学研究室Department of Biology, Faculty of Science, Kyushu University ◇ 福岡市西区元岡744 ◇ 744 Motooka, Nishi-ku, Fukuoka
発行日:2020年6月25日Published: June 25, 2020