遺伝子発現の調節はあらゆる生命現象の源である.実際に,この調節機構の乱れはさまざまな疾患の原因になることが知られている.したがって,生命現象を理解するためには,細胞の中で遺伝子発現がどのように制御されているかを明らかにする必要がある.また,この知見は医療や創薬などへの応用研究にも貢献することが期待される.RNAは,長らくDNAにコードされた遺伝情報をタンパク質へと変換する過程を仲立ちする分子として,その役割が知られてきたが,近年の研究で,RNAは遺伝子発現を転写や翻訳の各段階で調節することでさまざまな生命現象に関わることが明らかになりつつある.
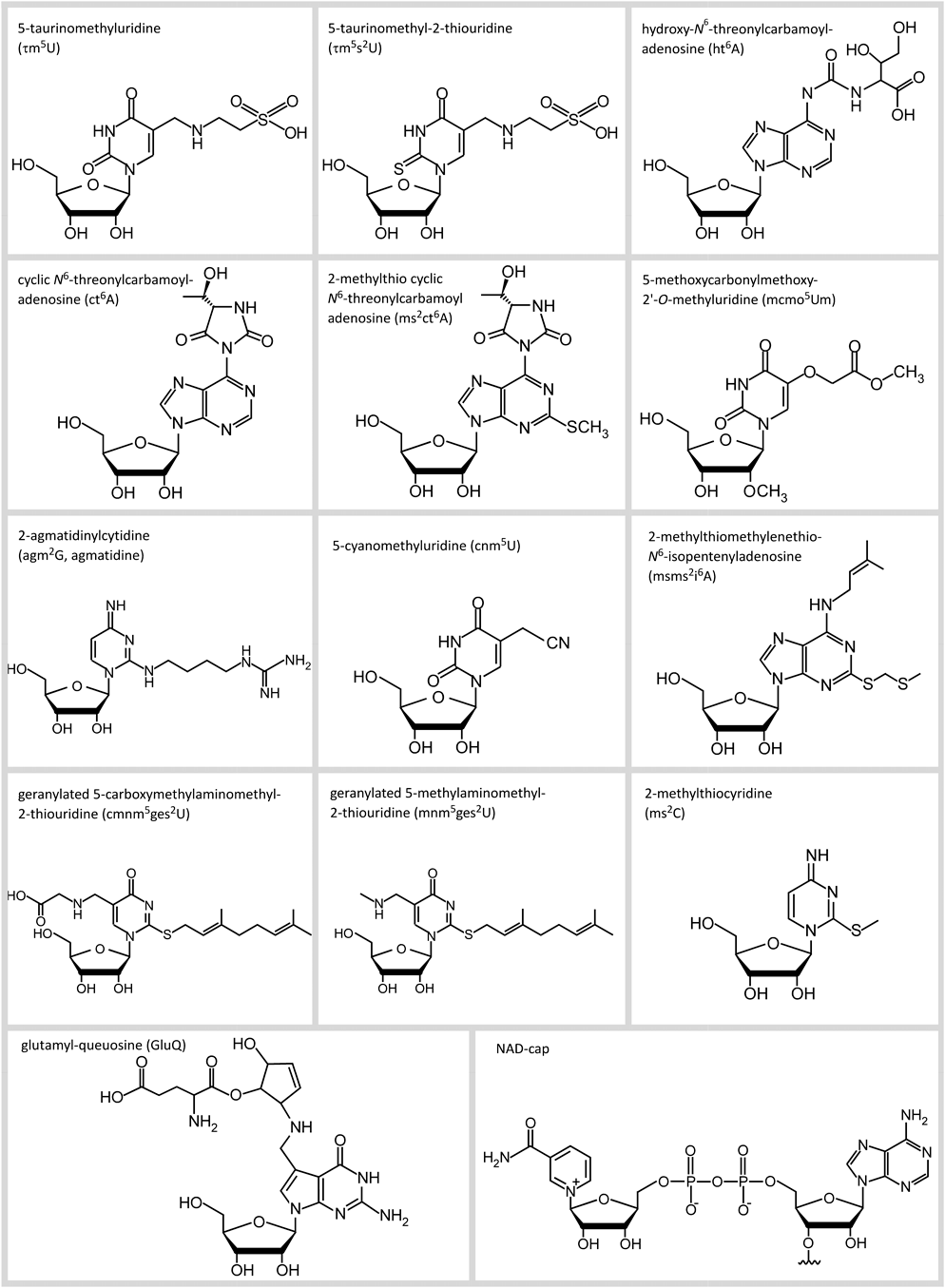
ゲノム配列が整備されたことに加え,次世代シーケンス技術(NGS)の登場により,RNAを配列情報や遺伝子の発現情報として捉えるトランスクリプトーム研究が盛んに行われている.しかし,RNAには転写後に付与される塩基の修飾やエディティング,末端構造といった質的な情報が含まれている.現在までに,約150種類のRNA修飾がさまざまな生物種から見つかっている1).これらの修飾はあらゆるRNAに普遍的に存在し,RNAが機能する上で欠くことのできない重要な質的情報である.塩基やリボースのメチル化に始まり,アセチル化,水酸化,硫化,セレノ化,還元,異性化,脱水環化,アミノ酸や糖の付加など,化学的にバリエーションに富んださまざまな修飾体が見つかっている.ここ数年でも,我々を含むいくつかのグループから新規のRNA修飾(図1)が報告されており,解析方法の発展とともにRNA修飾のケミカルスペースはさらに拡大することが期待されている.わずか数種類のDNA修飾と比較して,RNA修飾ははるかに多様であり,進化の過程において,RNAが新たな機能を担うためにRNA修飾を獲得したと捉えることもできる.
物理化学的な側面において,RNA修飾は,塩基やリボースのコンホメーションの固定や,局所的な疎水場や親水場を提供する.また,RNAの高次構造の形成や安定化に関与することも知られている.生化学的な側面においては,RNA結合タンパク質との相互作用,RNAプロセシングの制御,遺伝暗号の解読,遺伝情報の変化,翻訳調節などに関与する.また,RNA修飾は,RNAの細胞内局在を決定する目印としても働いている.自然免疫の受容体は,RNA修飾を識別することで自己と非自己を認識することが知られている.しかし,RNA修飾の機能は多様であり,未解明な部分が多く残されている.
RNA修飾の研究は歴史が古く,日本が大きな貢献を果たしてきた分野でもある.tRNAやrRNAなどの比較的存在量の多いRNAを対象に研究が行われてきたが,最近では,NGSを利用したエピトランスクリプトームシーケンス技術の発展により,真核生物mRNAにも5′末端のキャップ構造以外に,イノシン(I),N6-メチルアデノシン(m6A),5-メチルシチジン(m5C),シュードウリジン(Ψ),1-メチルアデノシン(m1A)などが大量に見いだされており,もはやゲノム配列から知りうる情報だけでRNAの機能は語れない状況にある2, 3).
RNA修飾の機能を明らかにするためには,RNA修飾を導入する修飾酵素(writer),いったん書き込んだ修飾を消去する脱メチル化酵素(eraser),RNA修飾を認識するタンパク質(reader)を同定する必要がある.m6Aに代表されるRNA修飾は,writerとeraserの働きによりダイナミックに変化し,その修飾情報をreaderが読み取ることで,RNAの分解やスプライシング制御,細胞内輸送や翻訳制御など,RNA修飾には,RNAのプロセシングやその後の働きを調節する役割があることが明らかになりつつある.実際に,m6A修飾の分布やプロファイルが組織や細胞が生育する環境によってダイナミックに変動する事例が相次いで報告されている4).しかし,その一方で,転写後に書き込まれたm6A修飾は静的で安定に存在し,ダイナミックには変動しないという報告5)もあり,RNA修飾が可逆的に変動するか?という,根本的な問題が提起され,現在,世界的に論争が巻き起こっている.
1970~80年代に数多くのRNA修飾が発見され,その化学構造が明らかにされた1).図1には今世紀に入ってから発見されたRNA修飾を示した.このうちの7種類は我々のグループによって報告されたものである.
5-taurinomethyluridine(τm5U)およびその2-チオウリジン誘導体(τm5s2U)は哺乳動物のミトコンドリアtRNAから発見されたRNA修飾である6).その後,この修飾の欠損がミトコンドリア病の原因であることが明らかになり,RNA修飾病(RNA modopathy)という新しい疾患の概念構築に貢献した7, 8).RNA精製技術の進歩9, 10)により,存在量の少ないミトコンドリアtRNAの単離が可能になったことと,RNAの高感度質量分析法(RNA-MS)の開発11)が,これらのRNA修飾の発見に貢献している.同様の理由で,hydroxy-N6-threonylcarbamoyladenosine(ht6A)は棘皮動物であるウニのミトコンドリアtRNAから発見された12).
cyclic N6-threonylcarbamoyladenosine(ct6A)13, 14)および2-methylthio cyclic N6-threonylcarbamoyladenosine(ms2ct6A)15)は大腸菌tRNAから発見された修飾体であるが,この修飾はt6Aが脱水環化したものである.1970年代にt6Aが発見され,40年以上にわたり,大腸菌をはじめとする多くの細菌tRNAにはt6Aが存在すると考えられてきた.しかし,t6Aは細胞からtRNAを抽出し,ヌクレオシドを解析する過程において,ct6Aが加水分解したアーティファクトであることが判明した14).加水分解しないような条件で解析を行った結果,細胞内にはct6Aの状態で存在することが判明している.このように,細胞内には化学的に不安定な天然物が存在し,抽出や分析のための前処理の過程で分解したアーティファクトをみている例が他にもある可能性を示唆している.実際,大腸菌においてtRNAAspのアンチコドンに存在するqueuosine(Q)は細胞内においてグルタミル化されたglutamyl-queuosine(GluQ)16)の形で存在していると考えられている.
2-agmatidinylcytidine(agm2C)17, 18)と5-cyanomethyluridine(cnm5U)19)はともにアーキアtRNAの解析から見つかったものである.アーキアのRNA修飾は十分に調べられていないため,今後も多くの新規修飾がさまざまなアーキア由来のRNAから見つかることが期待される.アーキアには,細菌や真核生物とはまったく異なる特徴的な代謝経路を持つ生物が存在することが知られており,アーキアはまさにRNA修飾の宝庫かもしれない.geranylated 5-methylaminomethyl-2-thiouridine(mnm5ges2U)やgeranylated 5-carboxymethylaminomethyl-2-thiouridine(cmnm5ges2U)20, 21)と,2-methylthiomethylenethio-N6-isopentenyladenosine(msms2i6A)22)はクロマトグラフィーを用いて,疎水性の高い修飾ヌクレオシドの解析の中から見つかったものであり,その存在量は非常に少ないが,RNA修飾反応の中間体23, 24)や過修飾体と考えられている.2-methylthiocytidine(ms2C)は最近報告された新規修飾である25).メチル化試薬の添加により増加することから,tRNAのs2C修飾のアルキル化損傷塩基の類である可能性が考えられているが,細胞内でも少量存在するようである.NAD+-capは細菌で見つかった新規のキャップ構造26, 27)であるが,その後真核生物にも見つかっている28).この修飾はRNAポリメラーゼがNAD+を転写のプライマーとして用いて転写を開始することで,RNA鎖に導入されると考えられている.存在量は少ないが,eraserとしてdecapping酵素が見つかっていることから,この修飾が担う機能に注目が集まっている29).
RNA修飾の生合成と機能を探るためには,RNA修飾を導入する修飾酵素やそのパートナータンパク質,基質を供給するための酵素などの同定が不可欠である.我々は,RNA修飾酵素に限らず,RNA修飾を形成するために必要な遺伝子をまとめてRNA修飾遺伝子と呼んでいる.RNA修飾遺伝子が同定できれば,組換えタンパク質を発現させて,試験管内でRNA修飾の再構成が可能になり,修飾反応の詳細な解析が可能になる.また,RNA修飾酵素と基質RNA複合体の結晶構造解析に道が開け,RNA修飾反応の分子基盤の理解など,構造生物学的な解析が可能になる.さらに,RNA修飾遺伝子をノックアウトした際の細胞やマウスの表現型解析から,RNA修飾が担う生理学的な機能や疾患との関連について初めて探求することができるようになる.我々は,RNA修飾遺伝子を網羅的に探索するための方法論であるリボヌクレオーム解析法を開発した11).この手法は,大腸菌や酵母の遺伝子破壊株ライブラリーからRNAを抽出し,質量分析法(LC/MS)により,RNA修飾の有無を網羅的にスクリーニングする方法である.標的となるRNA修飾の欠損がある機能未知遺伝子の欠損株で見つかれば,RNA修飾遺伝子を逆遺伝学的に同定することが可能になる.この手法の利点は,RNA修飾酵素のみならず,そのパートナータンパク質や,基質を供給するためのキャリアータンパク質など,RNA修飾の形成に必要な因子を網羅的に同定できる点にある.実際にこの手法を用いて,筆者らは大腸菌の2000以上の遺伝子,およびパン酵母においては400以上の遺伝子についてスクリーニングを行った結果,30個以上の新規RNA修飾遺伝子を同定することに成功した30–42).
さらに,新規なRNA修飾遺伝子を同定するためには,比較ゲノム解析が有効である.標的となるRNA修飾を持つ生物は対応するRNA修飾遺伝子を持つはずであり,その修飾を持たない生物には対応する修飾遺伝子は存在しないはずである.この仮定に基づき,複数の生物種において,標的となるRNA修飾の有無をLC/MSによって解析し,そのRNA修飾を持つ生物間で共通の遺伝子群を選出し,その中から,そのRNA修飾を持たない生物が有する遺伝子を排除することで,候補遺伝子の絞り込みを行う.この過程を生物種の組合わせを変えながら,試行錯誤することで,候補遺伝子を数個から数十個まで絞り込むことが可能である.ここから機能が明らかに異なる遺伝子を排除したり,予測される機能ドメインの有無を判別することで,さらに絞り込むことができる.最終的には遺伝子破壊株を取得して,LC/MSで標的となるRNA修飾の有無を判別することでRNA修飾遺伝子の同定を行う.本稿では,我々のグループが最近発見したRNA修飾遺伝子について紹介する.
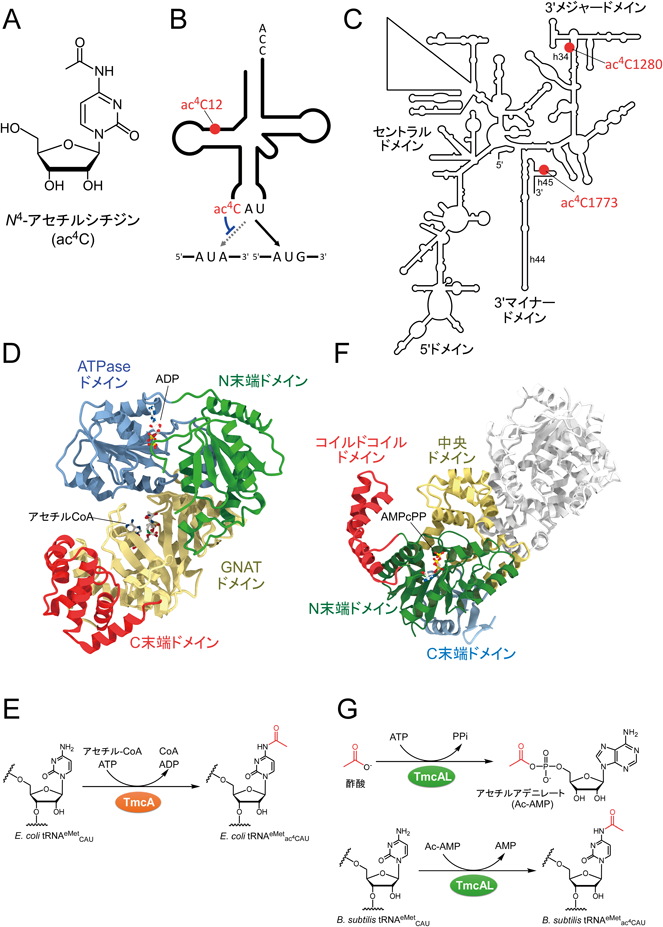
タンパク質が翻訳後にアセチル化されるように,RNAも転写後にアセチル化されることが知られている.RNAにおけるメジャーなアセチル化修飾ヌクレオシドとしてN4-アセチルシチジン(ac4C)(図2A)が知られている.ac4CはtRNAやrRNAに存在する.大腸菌など細菌ではac4C修飾がtRNAMetのアンチコドンの1字目に存在し,真核生物においてはtRNAの12位に存在する(図2B).さらに,真核生物の18S rRNAには,2か所にac4C修飾が存在する(図2C).我々は前述したリボヌクレオーム解析により,大腸菌の機能未知遺伝子群を網羅的にスクリーニングし,RNAアセチル化酵素TmcAを世界に先駆けて発見した30).TmcAにはアセチル化に必要なGNATドメインとRNAヘリカーゼドメイン(図2D)43)を持ち,アセチルCoAとATPを基質として,tRNAにac4C修飾を導入することが明らかとなった(図2E)30).一般的なアセチル化酵素によるタンパク質のアセチル化はアセチルCoAのみを基質として用いるが,TmcAはアセチルCoAに加えてATPを基質として用いていることが特徴である.ATPはRNAヘリカーゼの活性に必要であると考えられており,この活性を用いたRNAの構造変化がac4C修飾に必要であると解釈できる.
さらに,我々はTmcAの酵母ホモログであるKRE33/RRA144)やヒトホモログであるNAT1045)が,アセチルCoAとATPを基質として,18S rRNAにac4C修飾を導入することを明らかにした.酵母KRE33/RRA1は必須遺伝子であり,温度感受性変異株の解析から,KRE33/RRA1はrRNA前駆体のプロセシングおよび40Sサブユニットの成熟に必須であることを明らかにした44).さらに,KRE33/RRA1は核内アセチルCoAの濃度を感知することで18S rRNAのプロセシングを制御する可能性を示した44).また,NAT10もヒト細胞の生育に必要で,実際にNAT10をノックダウンするとアポトーシスが生じることが明らかとなった45).最近の研究で,抗ac4C抗体を用いた免疫沈降とNGS解析が行われ,ヒト細胞中のmRNA上に大量にac4C修飾が見つかったという報告がなされた46).さらに,これらのac4C修飾はNAT10によるものであると報告された.しかし,この論文では抗体の偽陽性をきちんと評価できていない点が問題である.さらに,NAT10は配列特異性が高くCCG配列の真ん中のCにac4C修飾を導入する44, 45)が,mRNA上で見つかった修飾部位にはこうした配列モチーフが見つかっておらず,我々はmRNAのac4C修飾については慎重に再検証する必要があると考えている.
TmcAとそのホモログは唯一見つかっているRNAアセチル化酵素であったが,我々は,さらに,TmcAホモログを持たない枯草菌においてもtRNAMetにac4C修飾が存在することを見いだした47).この事実は,枯草菌にはTmcA以外のアセチル化酵素が存在することを示唆している.比較ゲノムを用いた遺伝子探索の結果,枯草菌をはじめとする一群の細菌には,酢酸イオンを基質としてac4C修飾を導入するユニークなアセチル化酵素TmcALが存在することが明らかとなった(図2F)47).生化学および構造生物学的な解析の結果,TmcALはアミノアシルtRNA合成酵素と同じファミリーに属し,tRNAのアミノアシル化と同様な機構でアセチル化を行うことが判明した(図2G).TmcALは,まず酢酸イオンとATPからアセチルアデニレートを形成し,次に,アセチルアデニレートに対し,tRNAアンチコドン1字目シチジンのN4-アミノ基に求核置換攻撃させることで,ac4Cが形成される.さらなる解析により,TmcALが,AUAコドンの解読に必要なtRNAIleのアンチコドン修飾であるライシジン(L)の修飾酵素TilSとの間に,遺伝学的な相互作用があることが判明した.TmcALがない状態でTilSの働きを抑制すると,AUAコドンが,tRNAMetによって間違ってメチオニンとして解読されてしまうことを見いだし,ac4C修飾がAUAコドンの誤認識を妨げる役割を担っていることを明らかにした(図2B).これはac4C修飾の生物学的な役割を解明した最初の報告である.
ac4Cは細菌のtRNAに広くみられる修飾であるが,大腸菌およびγ-プロテオバクテリアではTmcAがac4C修飾を形成するのに対し,枯草菌,マイコプラズマを含む一群の細菌では,TmcALがac4Cを形成することが判明した.TmcAはGNATドメインを持ち(図2D),アセチルCoAを基質としてアセチル化する(図2E)のに対し,TmcALはRossmann foldを有し(図2F),酢酸イオンを基質にTmcAとはまったく異なる反応機構でac4Cを形成していた(図2G).細菌が進化の過程で,まったく異なる機構で同一のtRNA修飾を獲得したことはとても興味深い.魚とイルカが水中を速く泳ぐために類似の体形を獲得したように,遺伝暗号を正確に解読するために,異なる細菌の系統間で同じtRNA修飾を選択した,まさに分子レベルでの収斂進化といえよう.
5. 脊椎動物mRNAのキャップ構造におけるm6A修飾酵素の同定
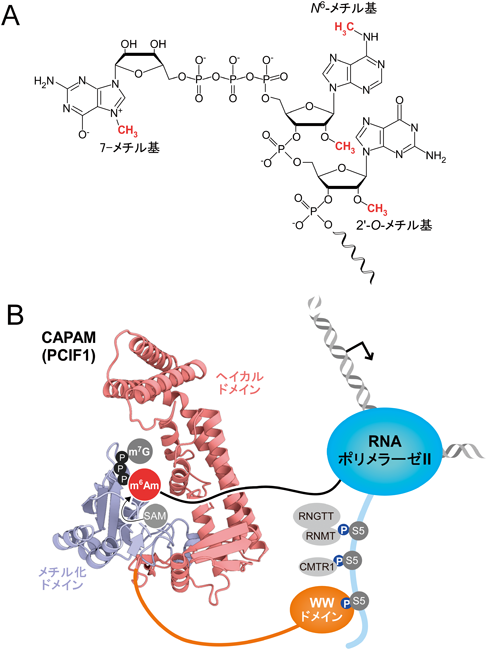
脊椎動物のmRNAや長鎖非コードRNAにN6-メチルアデノシン(m6A)が大量に見いだされ,m6AはRNAの代謝や正常な機能に重要であることが明らかになってきた4, 48).一般にm6AはmRNAの内部に存在しているが,脊椎動物では,mRNAの5′末端構造である7-メチルグアノシン(m7G)キャップ構造に続く1塩基目にもN6,2′-O-ジメチルアデノシン(m6Am)として存在する(図3A)49).このm6Am修飾の生合成や機能はほとんどわかっておらず,その解明のためにはm6Am修飾のN6-メチル基を導入する酵素の発見が必要であった.我々は,脊椎動物に保存された機能未知なメチル化酵素遺伝子の中から,酵母に存在するものを除いた結果,最終的に15種類の候補遺伝子を絞り込んだ.この中で,PCIF1遺伝子に着目した.PCIF1はRNAポリメラーゼII(RNAPII)のリン酸化されたC末端ドメイン(CTD)に結合する因子として同定された50).また,DNA m6Aメチル化酵素に類似したドメインを有している51)ことから,PCIF1は転写装置に結合して新生RNA鎖にm6A修飾を導入する可能性が浮上した.そこで,我々はHEK293T細胞でCRISPR/Cas9システムを用いてPCIF1のノックアウト(KO)株を作製した.この細胞からRNAを抽出し,抗キャップ抗体を用いてmRNAの5′末端断片を濃縮し,キャピラリー液体クロマトグラフィーナノエレクトロスプレーイオン化質量分析法を用いたRNA断片の直接的解析(RNA-MS)を行った.野生株由来のRNAからは,5′末端にm7Gpppm6Amを有する複数の断片が観測され,m7GpppAm断片との比率から,m6Amの修飾率は92%であると見積もられた.一方でPCIF1 KO細胞由来のRNAを解析したところ,m7Gpppm6Am断片は完全に消失し,m7GpppAm断片のみが観測された.この結果から,PCIF1はm6Am修飾のN6-メチル基を導入する修飾酵素であることが判明し,我々はPCIF1をCap-specific adenosine N6-methyltransferase(CAPAM)と命名した(図3B)52).CAPAMを欠損した細胞は酸化ストレスに対する感受性が増しており,m6Am修飾が生理学的に重要な意義を持つことが示唆された.生化学的な解析から,CAPAMはS-アデノシルメチオニン(SAM)をメチル基供与体として用い,m7Gキャップ構造およびm6Am修飾の2′-O-メチル基を特異的に認識することが明らかとなった.CAPAMのN末端に存在するWWドメインは,セリン5番がリン酸化されたRNAPIIのCTDに特異的に結合したことから,CAPAMは転写伸長の初期段階にRNAPIIへとリクルートされ,転写と共役しながらm6Am修飾を導入することが示唆された(図3B).東京大学理学部の濡木理研究室との共同研究により,結晶構造解析を行ったところ,CAPAMのコア部分は,メチル化ドメインとαヘリックスに富むヘリカルドメインの二つから構成されていることが判明した(図3B).m7Gキャップ構造はこれら二つのドメインの間のポケットで認識され,SAMはメチル化ドメインに特徴的なNPPFモチーフ配列からなる活性中心で認識されていた.これらの結晶構造はCAPAMによるキャップ構造特異的なN6-メチル基転移反応を理解するための分子基盤となる.他のグループによる先行研究では脱メチル化酵素であるFTO(fat mass and obesity associated gene)の過剰発現によってm6Am修飾率が低下しmRNAが不安定化されることが報告されている53).しかしm6Am修飾を完全に失ったCAPAM欠損細胞のmRNA量を網羅的に解析した結果,mRNA量に大きな変動はみられなかったことから,m6Am修飾はmRNAの安定性には寄与していないことが示された.一方でmRNAの翻訳効率を網羅的に解析したところ,m6Am修飾はmRNAの翻訳効率を向上する機能を持つことが示された.今後はm6Am修飾が変動することで調節される遺伝子発現機構の探求と,酸化ストレス応答との関係を明らかにすることを目指す.また,網羅的なスクリーニングからCAPAMは膀胱がんの抑制遺伝子としての働きが報告54)されているが,生理機能に関しては不明な点が多く残されている.CAPAMのモデルマウスの作出と表現型解析が,今後の研究に大きな展開をもたらすであろう.
6. 炭酸ガスに敏感なtRNA修飾とワールブルク効果
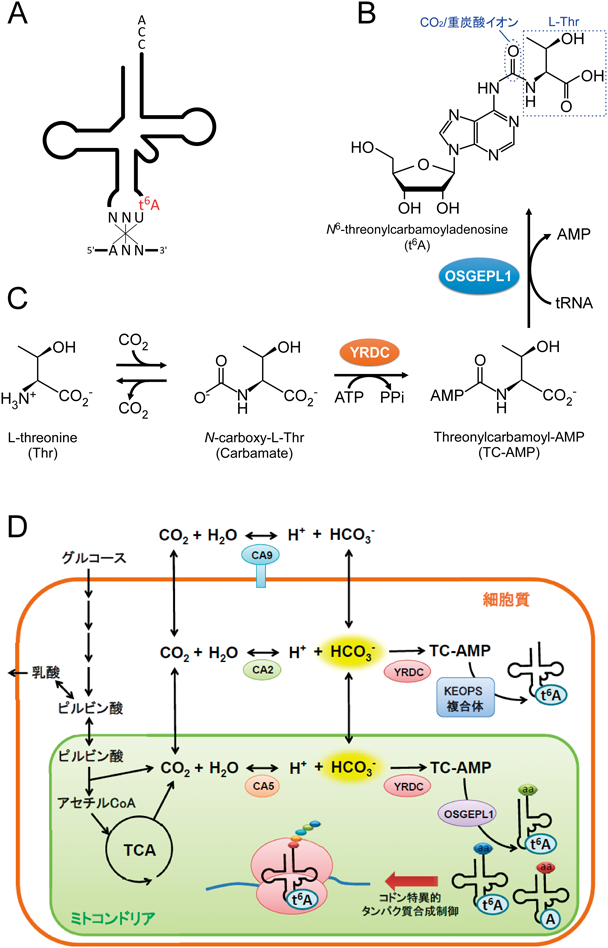
N6-threonylcarbamoyladenosine(t6A)(図4A, B)はすべての生物界で共通に用いられている修飾塩基であり,ANNコドンを解読するtRNAの37位(アンチコドン3′隣接塩基)に存在する(図4A).tRNAとmRNAが結合したリボソームの構造解析から,t6Aはコドン1字目のアデニンとスタッキング相互作用することでコドン-アンチコドン対合を安定化することが示されている55).実際にt6Aは,コドンの正確な認識や読み枠の維持,効率的なアミノアシル化や転座反応など,タンパク質合成のさまざまな過程に関わることが知られている.細菌においてt6A修飾に関わる酵素は必須遺伝子であり,この修飾の生理学的な重要性が示されている56).我々は,ヒトや哺乳動物のミトコンドリアにおいて,t6A37の修飾酵素としてYRDCとOSGEPL1を同定した57).ヒトミトコンドリアにおいて,t6Aは4種類のtRNAに存在する.t6AはアデニンのN6位にカルボニル基を介してThr残基が結合した修飾であるが,生合成においてカルボニル基は二酸化炭素(CO2)あるいは重炭酸イオン(または炭酸水素イオン,HCO3−)に由来し,Thr残基はL-Thrに由来することが知られている(図4B).はじめにL-ThrがCO2と非酵素的に反応し,カルバメート中間体を形成する(図4C).YRDCがATPを用いてこの中間体を活性化し,Threonylcarbamoyl-AMP(TC-AMP)を形成する(図4C).続いて,OSGEPL1がTC-AMPのThreonylcarbamoyl基をtRNAのA37位へと転移し,t6Aが形成される(図4B, C).YRDCは主に細胞質tRNAのt6A形成に関わる一方,弱いミトコンドリア移行シグナルを有し,一部はミトコンドリアtRNAのt6A形成にも関わることが明らかとなった.OSGEPL1はミトコンドリアのみに局在し,ミトコンドリアtRNAのt6A37形成を担っていることが判明した.実際に,OSGEPL1をノックアウトするとミトコンドリアtRNAのt6Aが完全に欠失し,タンパク質合成能は低下し,呼吸不全を引き起こすことを見いだした.
次に,YRDCとOSGEPL1の組換えタンパク質を取得し,L-Thr, ATP, HCO3−を基質に,試験管内で修飾の再構成を試みたところ,tRNA上に効率よくt6A修飾を導入することに成功した.各基質に対して,酵素反応速度論的な解析を行った.四つの基質のうち,ミトコンドリアtRNA,L-Thr, ATPに対するKm値は十分に低い値を示したが,驚いたことに,HCO3−に対するKm値は異常に高く,31 mMであった.したがって,t6A修飾反応の律速段階はHCO3−濃度であるという結果を得た.前述したようにt6A修飾の第一段階はL-ThrがCO2と非酵素的に反応しカルバメート中間体を形成する(図4C).この過程は平衡状態であると考えられることから,HCO3−に対する高いKm値は非酵素的なカルバメート形成によって説明できる.ミトコンドリア内のHCO3−濃度は10~40 mMである58)ことから,生理的な条件でt6A修飾は変動しうると考えた.実際に,ヒト細胞株を低いHCO3−濃度の培地で培養したところ,複数のtRNAでt6A修飾率が顕著に減少する結果を得た.一般的に,tRNA修飾は静的で安定であるとであると考えられてきたが,この結果から,t6A37修飾はHCO3−濃度を感知してダイナミックに変化することが判明した.t6A37修飾の変動は直接的にtRNAの暗号解読能に影響するため,呼吸の状態や細胞が置かれた環境によって,ミトコンドリアの翻訳が制御されると考えられる.ミトコンドリア内のCO2は主に,好気的条件下において,ピルビン酸がアセチルCoAに変換される過程とTCA回路から生じることが知られている.CO2はカルボニックアンヒドラーゼ(CA)の作用により,水と反応して重炭酸イオン(HCO3−)に変換される.低酸素下では,CA9が細胞膜上に高発現し,細胞外で大量のHCO3−を生成する.HCO3−は嫌気的解糖系で生じた乳酸を中和し,アシドーシスを防ぐ役割がある.今回の研究で新たにHCO3−はt6A修飾の基質となり,tRNAの機能に必須の役割が明らかとなった.低酸素環境下では,ミトコンドリア内部からのCO2の供給量が低下し,t6A修飾率が減少すると考えられる(図4D).t6A修飾の低下はミトコンドリアタンパク質合成能を直接的に低下させることから,この現象は,低酸素下で細胞がミトコンドリアの機能を制限するワールブルク効果を説明するメカニズムの一つであると考えられる.
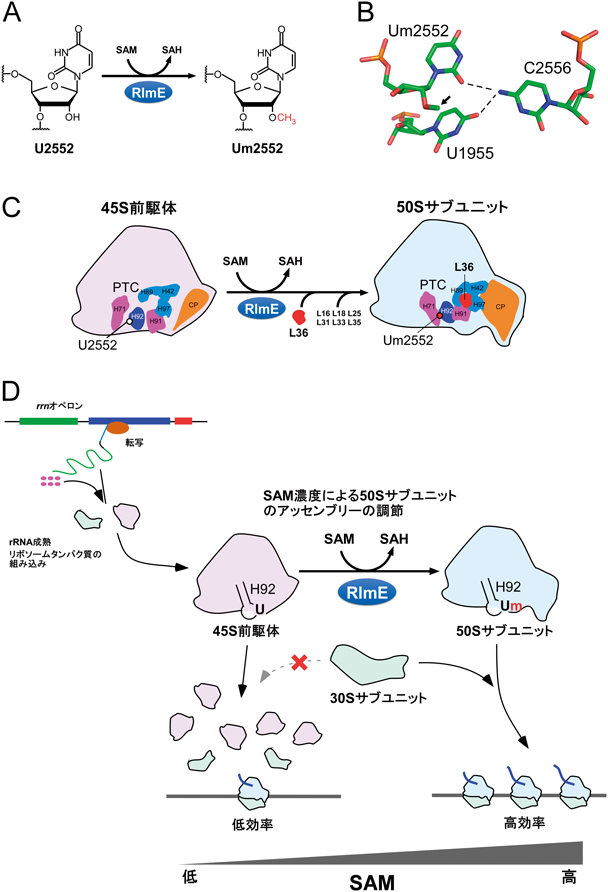
7. リボソーム50Sサブユニットの後期アッセンブリーを促進するSAM濃度依存的なrRNAメチル化修飾
リボソームの生合成過程において,RNAヘリカーゼやrRNA修飾酵素などのアッセンブリー因子が重要な役割を担っているが,個々のアッセンブリー因子がどのような分子機構でリボソームの生合成に関与するかについては,不明な点が多く残されている.大腸菌において,RlmEはSAMをメチル基供与体として,23S rRNAのHelix 92(H92)に存在する2552位のウリジンを2′-O-メチル化(Um2552)する修飾酵素である(図5A).我々は,50Sサブユニットの後期アッセンブリー過程において,RlmEによるUm2552のたった1か所のメチル化が,45S前駆体から50Sサブユニットへの成熟を促進する役割があることを示した59).Um2552のメチル化は,リボースのパッカリングをC3′endo型に固定することでC2556およびU1955からなるbase triple(図5B)を安定化させ,ドメインIVとVの会合を促すと考えられる.その結果,L36や他のリボソームタンパク質の組込みを促進することで,45S前駆体から50Sへの生合成が進行することが判明した(図5C).この成果は,rRNA修飾の作用メカニズムを示したことのみならず,アッセンブリー因子の酵素活性によるリボソーム生合成の一部を再現した初めての知見である.
SAMは細胞内の主要なメチル基供与体であり,DNA, RNA,タンパク質などのメチル化や低分子代謝物の生合成を通じてさまざまな細胞内プロセスに関与する.SAMの欠乏はこれらメチル化の低下を招き,細胞の増殖や分化,脳の機能などに影響を与え,さまざまな疾患の原因になっていることが知られている.細胞内のSAM濃度が約1/3程度まで減少する大腸菌株(Δmtn)は増殖速度が低下することが知られているが,我々はこのΔmtn株でリボソームを解析したところ,45S前駆体が顕著に蓄積することを見いだした.また,Δmtn株にRlmEを過剰発現すると蓄積した45S前駆体が減少するとともに,生育速度が優位に回復することを見いだした.SAMは,生体高分子のメチル化修飾のみならず,生体内のさまざまな代謝に関わる.しかし,この観測結果から,Um2552はSAM濃度に敏感なメチル化修飾であり,細胞がSAM濃度の低下を感知し,リボソームの生合成を止めることで生育速度を調節する機構が明らかになった(図5D)60).
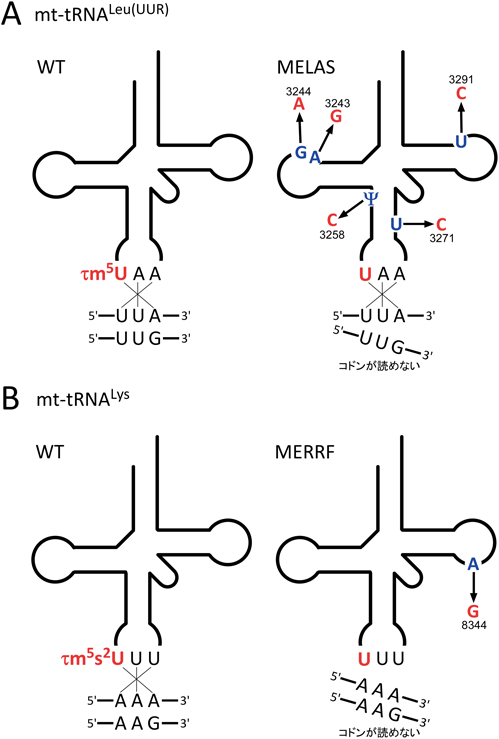
前述したように,我々はヒトミトコンドリアtRNAからタウリンを含む修飾塩基5-taurinomethyluridine(τm5U)とその2-チオウリジン誘導体(τm5s2U)を発見した(図1)6).これらの修飾はtRNALeu(UUR)やtRNALysを含む5種類のtRNAのアンチコドン1字目に存在し,コドン解読に重要な役割を担っている8).
ミトコンドリア脳筋症は,ミトコンドリアの機能異常が原因で生じる重篤な疾患であり,エネルギー消費の高い脳や骨格筋など臓器を中心に障害が生じる.ミトコンドリア脳筋症の代表病型のうち,脳卒中を特徴とするMELAS(mitochondrial myopathy, encephalopathy, lactic acidosis and stroke-like episodes)や,てんかんを特徴とするMERRF(myoclonus epilepsy associated with ragged-red fibers)は,それぞれミトコンドリアDNAにコードされたtRNALeu(UUR)遺伝子およびtRNALys遺伝子上の点変異が原因で発症することが知られていた.我々は,MELASおよびMERRF患者由来の細胞から変異tRNAを単離し,質量分析法を用いた詳細な解析を行ったところ,タウリン修飾(τm5Uやτm5s2U)がこれらの変異tRNAでは著しく減少し,未修飾Uのままになっていることが判明した(図6A, B)7).MELASやMERRFの原因点変異は,τm5U修飾酵素の認識を妨げることで,タウリン修飾の欠損をもたらすと考えられた.これらの変異tRNAはコドンの認識能が低下し,タンパク質合成能とミトコンドリアの活性が低下することが明らかとなっている.タウリン修飾を欠損したMELAS由来の変異tRNALeu(UUR)は,対応する二つのコドンUUAとUUGのうち,UUGコドンのみを特異的に翻訳できないことが明らかとなった(図6A)61).また,同じくタウリン修飾を欠損したMERRF由来の変異tRNALysは,対応するコドンであるAAAとAAGをともに認識できなくなる(図6B)ことで,結果としてミトコンドリアタンパク質合成能が全体的に低下することが判明した62).リボソームのA部位におけるtRNAとコドンの対合実験によって,タウリン修飾を欠損したtRNALeu(UUR)は,正常tRNALeu(UUR)と同様にUUAコドンとは対合できるものの,UUGコドンとは対合できなかったことから,タウリン修飾はU-Gのwobble対合を安定化させる役割があることが示された61, 63).また実際に,MELAS細胞でリボソームプロファイリングが行われた結果,UUGコドンにリボソームが蓄積しやすい64)という結果が示され,タウリン修飾の欠損によりUUGコドンの翻訳能が低下する,という我々が提唱したモデルが裏づけられた結果となった.先行研究により,MELASの変異率が上がると,ミトコンドリアにおける呼吸鎖酵素複合体IのサブユニットであるND6のタンパク質合成量が顕著に低下するという現象が報告されている65).ND6はmtDNAにコードされる13種類の遺伝子の中でもUUGコドンの使用頻度が顕著に高く,タウリン修飾の欠損がUUGコドンリッチなタンパク質の発現を特異的に低下させていると考えられる.この仕組みは,長年分子レベルでの原因が不明であった,呼吸鎖酵素複合体Iの活性低下というMELAS患者の生化学的症状をうまく説明することができる7).
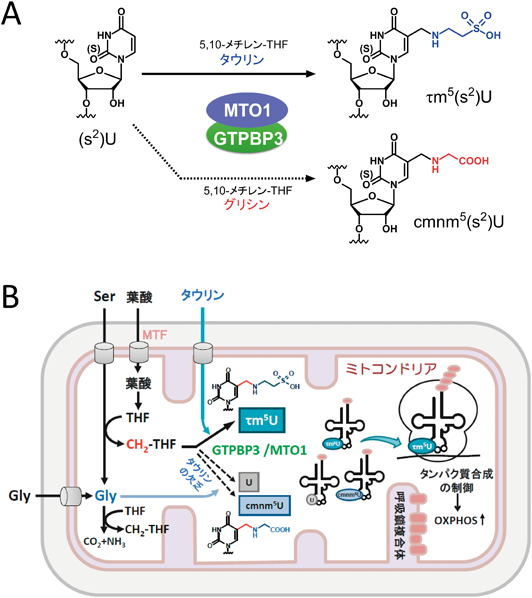
我々は酵母を用いた研究でMTO1とGTPBP3(MSS1)がミトコンドリアtRNAのタウリン修飾酵素であることを報告している(図7A)66).さらに,タウリン修飾の基質としてタウリンとメチレンテトラヒドロ葉酸を同定した67).実際に,MTO1とGTPBP3の組換えタンパク質を取得し,タウリン,メチレンテトラヒドロ葉酸,GTP,各種補酵素の存在下で,試験管内で修飾の再構成を試みたところ,低効率ながらtRNA上にτm5U修飾を導入することに成功した(図7A)67).
主に食事から摂取するタウリンがRNA修飾の基質でもあるという知見は医療関係者や食品業界からも注目されている.ネコやキツネなどの肉食系の動物はタウリンを生合成できないため,タウリンは必須の栄養素である.実際に,タウリンの欠乏により失明や心筋症を発症することが知られている.また,ヒラメの養殖においてタウリンは正常な生育に必須であることも知られている.一般にタウリンは,システインから生合成されるが,これらの動物ではこの生合成活性が低下していることから,タウリンを食餌から摂取する必要がある.ヒトを含む霊長類は,タウリンを生合成することができるが,新生児はタウリン合成能が低いため,母乳からタウリンを摂取することが正常な発育に必須である68).我々は,タウリンの欠乏がτm5U修飾に影響を与える関係を調べるために,タウリンの欠乏により心筋症を発症したネコの肝臓からミトコンドリアtRNAを単離し,修飾の状態を調べたところ,予想どおり,τm5Uの修飾率が有意に低下していた.同様に,タウリン欠乏食を与え飼育したヒラメからミトコンドリアtRNAを単離し,修飾を調べたところ,同様にτm5U修飾の低下を観測した.さらに,HeLa細胞をタウリンを欠乏した培地で培養したところ,ミトコンドリアtRNAのτm5Uの修飾率が顕著に低下した(図7B)67).ヒト細胞ではタウリンのde novo合成経路が働いていると考えられるが,外界からのタウリンの供給が高い修飾率の維持に必須であることを示した.さらに興味深いことに,タウリン修飾の低下とともに,tRNAの同じ位置に5-carboxymethylaminomethyluridine(cmnm5U)修飾が生じることを見いだした(図7AB)67).cmnm5Uは細菌,酵母や線虫のミトコンドリアtRNAにみられる修飾であり,τm5Uのタウリンの代わりにグリシンが取り込まれた構造をしている(図7A).タウリンとグリシンは化学構造的に類似していることから,タウリンが欠乏した状況でグリシンが取り込まれたものと考えられる.培地にタウリンを添加するとcmnm5Uは完全に消失し,τm5Uの修飾率が上昇することを確認している.この知見は,生理的な条件において,メタボライトの濃度変化により,RNA修飾の化学構造が変化することを示した初めての例である.cmnm5Uが取り込まれたtRNAがどのように振る舞い,タンパク質合成に影響を与えるかについては今後の研究が必要であるが,τm5Uとcmnm5Uは化学的な性質が大きく異なることから,修飾構造の違いによるコドン解読能へ与える影響が考えられる.今後は,タウリン欠乏症や高グリシン血症においてtRNA修飾が変化することで,翻訳に影響を与え,最終的な表現型や症状につながる可能性について探求していきたい.
また,τm5U修飾のウラシル5位に結合しているメチル基は,メチレンテトラヒドロキシ葉酸(メチレンTHF)を由来とすることが判明した.葉酸誘導体はC1炭素を運搬し,さまざまな代謝物に転移する役割を担う.ミトコンドリアにおいて,メチレンTHFは,SHMT2の作用により,Serのβ炭素がTHFに転移されることで形成される.実際に,ミトコンドリア葉酸トランスポーター(MTF)やSHMT2に変異の入ったCHO細胞では,τm5U修飾率が顕著に低下することを見いだしている67).また,興味深いことに,SHMT2をノックアウトした細胞ではミトコンドリアの活性が顕著に低下すること,またこの原因として,tRNAのタウリン修飾の低下が主な原因であることが判明している64).SHMT2はさまざまながんで高発現していることが知られており,肝臓がんでは,SHMT2の抑制で腫瘍形成能が低下することが知られている69, 70).また,ヒトの老化とともに,ミトコンドリアの活性が低下することが知られているが,原因として,SHMT2発現量の低下が示唆されている71).老化に伴い,τm5Uの修飾率が低下する可能性が考えられる.また,受胎前後における葉酸の欠乏は,胎児の神経管閉鎖障害(neural tube defects:NTDs)を引き起こし,二分脊椎症の原因となることが知られている72).τm5U修飾の減少によるミトコンドリアの活性低下がこの疾患の原因である可能性が考えられる.
さらに,我々の研究成果を基に,川崎医科大学神経内科学講座においてMELAS患者にタウリンを経口投与する医師主導臨床治験が行われ,MELASの脳卒中様発作の再発抑制効果が証明された73, 74).実際に,2019年2月にタウリンを主成分とする薬が,保険適応薬として正式に認可された.
これらの研究は,RNA修飾の異常が疾患の一義的な要因であることを示した世界で最初の例であり,我々は疾患の新しいカテゴリーとしてRNA修飾病(RNA modopathy)を提唱している67).実際に,ヒトRNA修飾酵素の異常が原因の疾患が続々と報告されている(表1)75, 76).前述したタウリン修飾酵素であるMTO1の変異は,肥大性心筋症や乳酸アシドーシスを引き起こすことが報告されている77, 78).患者細胞は,酸素消費量の減少と呼吸鎖複合体の活性低下がみられ,ミトコンドリアの機能異常を示している.Mto1のノックアウトマウスはタウリン修飾の欠損を引き起こし,ミトコンドリアタンパク質合成の異常をもたらした.また,細胞質にタンパク質の凝集体が蓄積するという興味深い表現型を示した79).一方で,GTPBP3の変異も呼吸鎖不全症を引き起こすが,MTO1変異と異なり,Leigh脳症様の脳疾患を伴うという特徴がある80).また我々はMTU1がτm5s2Uのチオ化修飾酵素であることを報告した66).MTU1が欠損すると,ミトコンドリアタンパク質合成能が顕著に低下するとともに,ミトコンドリアの活性が著しく低下することが明らかとなっている.また,小児性急性肝不全患者でMTU1(TRMU)遺伝子の変異が見つかっており,患者の肝臓においてミトコンドリアtRNAのチオ化修飾の著しい減少と,呼吸鎖複合体の活性低下が報告されている81).Mtu1のノックアウトマウスは胚性致死(E7.5~8)を示し,ミトコンドリアtRNAのチオ化修飾が発生と分化に必須の役割を担っていることが判明した82).さらに,Mtu1の肝臓特異的なコンディショナルノックアウトマウスでは,血清乳酸値,GOT, GPTの増加とアルブミン濃度の低下,すなわち肝臓の炎症や傷害が確認された82).この表現型から,MTU1の欠損が小児性急性肝不全の直接的な原因であることが証明された.これらタウリン修飾酵素の変異がミトコンドリアの機能異常を示すという知見は,MELASやMERRFの主な原因がタウリン修飾欠損で生じることを裏づけている.
表1 RNA修飾の異常が原因と考えられる疾患| 疾患 | 遺伝子 | RNA修飾 | RNA種 |
|---|
| ミトコンドリア脳筋症(MELAS) | mt-tRNALeu(UUR) | τm5U | mt-tRNA |
| ミトコンドリア脳筋症(MERRF) | mt-tRNALys | τm5s2U | mt-tRNA |
| 拡張型心筋症 | MTO1 | τm5U | mt-tRNA |
| 拡張型心筋症 | GTPBP3 | τm5U | mt-tRNA |
| 小児性急性肝不全 | MTU1 | τm5s2U | mt-tRNA |
| 複合型呼吸鎖異常症 | NSUN3 | f5C | mt-tRNA |
| HSD10ミトコンドリア病 | TRMT10A (MRPP1) | m1A, m1G | mt-tRNA |
| HSD10ミトコンドリア病 | SDR5C1 (MRPP2) | m1A, m1G | mt-tRNA |
| 小児肥満(predicted) | METTL15 | m4C | mt-12S rRNA |
| MELAS様症候群 | MRM2/FTSJ2 | 2′-O-methylation | mt-16S rRNA |
| 鉄芽球性貧血(MLASA) | PUS1 | Ψ | cyto-tRNA, mt-tRNA |
| 複合型酸化的リン酸化異常 | TRIT1 | i6A | cyto-tRNA, mt-tRNA |
| 複合型酸化的リン酸化異常 | TRMT5 | m1G | cyto-tRNA, mt-tRNA |
| 知的障害,Dubowitz様症候群 | NSUN2 | m5C | cyto-tRNA, mt-tRNA |
| 知的障害 | TRMT1 | m2,2G | cyto-tRNA, mt-tRNA |
| 重度の神経発達障害 | PUS3 | Ψ | cyto-tRNA, mt-tRNA |
| 2型糖尿病 | Cdkal1 | ms2t6A | cyto-tRNA |
| 家族性自律神経失調症 | IKBKAP/ELP1 | mcm5U | cyto-tRNA |
| 知的障害 | ELP2 | mcm5U | cyto-tRNA |
| 筋萎縮性側索硬化症(ALS) | ELP3 | mcm5U | cyto-tRNA |
| 知的障害(DREAM-PL) | CTU2 | mcm5s2U | cyto-tRNA |
| 知的障害 | FTSJ1 | 2′-O-methylation | cyto-tRNA |
| 小頭症,小人症 | METTL1 | m7G | cyto-tRNA |
| 膀胱がん | ALKBH8 | mchm5U | cyto-tRNA |
| 知的障害,斜視,小頭症,成長遅延 | ADAT3 | I | cyto-tRNA |
| 乳がん | TRMT12/TYW2 | yW | cyto-tRNA |
| 乳がん | TRMT2A | m5U | cyto-tRNA |
| Galloway-Mowat症候群(奇形,脳の発達障害) | KEOPS | t6A | cyto-tRNA |
| 知的障害,小頭症 | METTL5 | m6A | 18S rRNA |
| 結腸直腸がん | NAT10 | ac4C | 18S rRNA, cyto-tRNA |
| 乳がん | FTSJ3 | 2′-O-methylation | 28S rRNA |
| 先天性角化不全症 | DKC1 (dyskerin) | Ψ | 28S and 18S rRNAs |
| Bowen-Conradi症候群 | EMG1 (Nep1) | m1acp3Ψ | 18S rRNA |
| 遺伝性対側性色素異常症(DSH),Aicardi-Goutières症候群(AGS) | ADAR1 | I | mRNA |
| 肥満,アルツハイマー病 | FTO | demethylation of m6A | mRNA, UsnRNA |
| 神経膠芽腫,乳がん | ALKBH5 | demethylation of m6A | mRNA |
| 膀胱がん | CAPAM | m6A (cap-specific) | mRNA, lncRNA |
| 神経膠芽腫,肝臓がん,乳がん,急性骨髄性白血病 | METTL3 | m6A | mRNA, lncRNA |
| 神経膠芽腫,肝臓がん,急性骨髄性白血病 | METTL14 | m6A | mRNA, lncRNA |
| 知的障害,頭蓋顔面形態異常(マウス) | METTL4 | m6A | U2 snRNA |
ヒトミトコンドリアtRNAMetのアンチコドン1字目には5-formylcytidine(f5C)修飾が存在する83).この修飾が,AUGコドンに加えてAUAコドンをMetに解読するために必須であることは知られていた84)が,f5C修飾の生合成や,細胞および個体レベルにおける生理機能については未解明であった.我々はf5C修飾が,NSUN3によるシトシン5位をメチル化することで5-メチルシチジン(m5C)を形成し85),それに続いてALKBH1がm5Cをヒドロキシル化と酸化を行う86)ことで,f5C修飾を生合成する機構を明らかにした.NSUN3やALKBH1をノックアウトした細胞においては,ミトコンドリアtRNAMetにおいてf5C修飾が形成されず,ミトコンドリアタンパク質の合成能およびミトコンドリアの活性が顕著に低下した.実際,複合型呼吸鎖異常症の患者からNSUN3の機能欠損型変異が見つかっており,f5C修飾の欠損がミトコンドリア病の原因となることも示されている87).
ミトコンドリア筋症を併発する鉄芽球性貧血(mitochondrial myopathy and sideroblastic anemia:MLASA)ではPUS1(pseudouridine synthase 1)のミスセンス変異が報告されている88).PUS1はtRNA, U2 snRNA, mRNAなどさまざまなRNAにΨを導入する酵素である.実際,患者細胞から抽出したtRNAにはΨの減少がみられたことから,PUS1の基質であるtRNAや他の機能性RNAの機能不全が疾患の原因であると考えられる.PUS1はミトコンドリアtRNAのみならず,細胞質のtRNAやmRNAにもΨを導入することが知られており,MLASAは複合的な要因で発症する可能性がある.
先天性角化不全症(dyskeratosis congenita:DC)は早期老化,間質性肺線維症を伴いがんを発症しやすい疾患である.原因遺伝子DKC1はH/ACA snoRNPのシュードウリジン(Ψ)合成酵素ジスケリン(dyskerin)をコードしている.DKC1変異マウスは骨髄形成不全を引き起こし肺と乳腺に発がんが観測され,ヒトDCの表現型を示した89).実際に,rRNA中に含まれるΨの減少が観測されており,RNA修飾欠損に起因するリボソームの機能低下が疾患の原因と考えられている.実際,Ψを欠いたリボソームはtRNAやIRESとの結合能が低下していることから,DCの患者の細胞では翻訳開始に異常が生じている可能性が提唱されている90).
Cdkal1は人種を超えて2型糖尿病との関連が報告されていた91).Cdkal1は細胞質tRNALysの37位に存在する2-methylthio-N6-threonylcarbamoyladenosine(ms2t6A)の2-メチルチオ基を導入する修飾酵素の遺伝子である.Cdkal1のβ細胞特異的なノックアウトマウスは,膵臓においてms2t6A修飾が大きく減少するとともに,インスリンの分泌が低下し,血糖値がコントロールできなくなった92).2-メチルチオ修飾の欠損は,tRNALysの翻訳能を減弱させ,結果的にプロインスリンの発現低下につながるのではないかと考えられている.
家族性自律神経失調症(familial dysautonomia:FD)は,神経の発達異常により,感覚神経と自律神経に障害をもたらす先天性の遺伝病である.ELP1(IKBKAP)遺伝子の異常スプライシングが原因であることが知られている93).ELP1は細胞質tRNAのアンチコドン1字目に存在するウリジン修飾である5-methoxycarbonylmethyluridine(mcm5U)とその誘導体の生合成に不可欠な因子の遺伝子である.実際に,FD患者細胞ではtRNAのmcm5U修飾が顕著に低下していることが判明している94, 95).この修飾はコドンの精確な解読に重要な役割を担っていることから,修飾欠損はタンパク質合成の異常を引き起こし,最終的に神経の発達異常や変性につながっていると考えられるが,その詳細なメカニズムは未解明である.興味深いことに,mcm5U修飾の生合成に関わるELP2の変異が知的障害96),ELP3の変異が筋萎縮性側索硬化症(ALS)の発症と関連があることが報告されている97).
他にも知的障害に関係するRNA修飾遺伝子として,NSUN298),TRMT199),FTSJ1100),CTU2101),ADAT3102),METTL4103),METTL5104)などが見つかっている.いずれもtRNA, rRNA, U snRNAの修飾酵素の遺伝子であり,なぜユビキタスに発現している遺伝子の異常が脳特異的な症状を引き起こすのか,非常に興味深い.
RNA修飾の研究は今後も拡大し続けるであろう.しかし,RNA修飾を解析するための手法には限界があり,この分野のさらなる発展には,革新的な解析技術の開発が不可欠である.新規のRNA修飾の探索には,天然物化学や分析化学のパワフルな解析技術を積極的に導入することが必要であろう.NGSを用いたエピトランスクリプトームシーケンス技術は,RNA修飾の大規模かつハイスループットな探索を可能にしているが,一方で偽陽性の問題があり,マッピング精度の低さが大きな問題になっている.より精度の高いシーケンス技術の開発が不可欠である.RNA修飾の変動により,遺伝子発現が制御されるという概念を確立するためには,個々のRNA修飾部位の修飾率を簡便かつ精度よく解析する手法が必要である.また,本稿ではふれなかったが,RNA修飾酵素をエンジニアリングすることで,さまざまなRNA修飾を狙った位置に導入することができれば,遺伝子発現や生命機能の制御を可能にするRNA修飾編集技術が誕生するであろう.また人為的なRNA修飾の導入技術は,RNAセンサーによる認識を妨げることにより,自然免疫から回避させることが可能になるなど,求める機能を自由自在にデザインすることが可能になるであろう105).実際,デアミナーゼをエンジニアリングすることで,DNA編集やRNA編集が可能になっている106).企業も含めた異分野からの参入や,若い研究者の活躍に期待したい.
この原稿を書いている最中に,新型コロナウイルスの感染拡大で人類は,これまでに経験したことのないような危機に直面しています.コロナウイルスはプラス鎖RNAウイルスなので,私も含め多くのRNA研究者たちが,今できることを探しています.SARS-CoV-2は,2つのRNAメチル化酵素をコードしています.いずれも自身のRNAゲノムのキャップ構造を修飾することで,宿主の翻訳マシナリーに認識されるようになります.また,宿主の自然免疫系から逃れる機能を獲得しています.これらのRNAメチル化酵素に対する阻害剤が開発できれば,将来的にユニークな作用機序を持つ抗ウイルス薬が作れるかもしれません.私たちは自分たちの専門性を生かして少しでもここ危機に立ち向かう努力を続けていきたいと考えています.ぜひ,日本RNA学会からの情報発信にご注目いただければと思います.https://www.rnaj.org/
謝辞Acknowledgments
本稿で紹介した研究は,研究室のスタッフと学生によって行われた.また,多くの共同研究者に支えられた.この場を借りて感謝申しあげたい.
引用文献References
1) Boccaletto, P., Machnicka, M.A., Purta, E., Piątkowski, P., Bagiński, B., Wirecki, T.K., de Crécy-Lagard, V., Ross, R., Limbach, P.A., Kotter, A., et al. (2018) MODOMICS: a database of RNA modification pathways. 2017 update. Nucleic Acids Res., 46(D1), D303–D307.
2) Frye, M., Jaffrey, S.R., Pan, T., Rechavi, G., & Suzuki, T. (2016) RNA modifications: what have we learned and where are we headed? Nat. Rev. Genet., 17, 365–372.
3) Harcourt, E.M., Kietrys, A.M., & Kool, E.T. (2017) Chemical and structural effects of base modifications in messenger RNA. Nature, 541, 339–346.
4) Roundtree, I.A., Evans, M.E., Pan, T., & He, C. (2017) Dynamic RNA modifications in gene expression regulation. Cell, 169, 1187–1200.
5) Ke, S., Pandya-Jones, A., Saito, Y., Fak, J.J., Vagbo, C.B., Geula, S., Hanna, J.H., Black, D.L., Darnell, J.E. Jr., & Darnell, R.B. (2017) m(6)A mRNA modifications are deposited in nascent pre-mRNA and are not required for splicing but do specify cytoplasmic turnover. Genes Dev., 31, 990–1006.
6) Suzuki, T., Suzuki, T., Wada, T., Saigo, K., & Watanabe, K. (2002) Taurine as a constituent of mitochondrial tRNAs: new insights into the functions of taurine and human mitochondrial diseases. EMBO J., 21, 6581–6589.
7) Suzuki, T., Nagano, T., & Suzuki, T. (2011a) Human mitochondrial diseases caused by lack of taurine modification in mitochondrial tRNAs. WIREs RNA, 2, 376–386.
8) Suzuki, T., Nagao, A., & Suzuki, T. (2011b) Human Mitochondrial tRNAs: Biogenesis, Function, Structural Aspects, and Diseases. Annu. Rev. Genet., 45, 299–329.
9) Miyauchi, K., Ohara, T., & Suzuki, T. (2007) Automated parallel isolation of multiple species of non-coding RNAs by the reciprocal circulating chromatography method. Nucleic Acids Res., 35, e24.
10) Suzuki, T. & Suzuki, T. (2007) Chaplet column chromatography: isolation of a large set of individual RNAs in a single step. Methods Enzymol., 425, 231–239.
11) Suzuki, T., Ikeuchi, Y., Noma, A., Suzuki, T., & Sakaguchi, Y. (2007) Mass spectrometric identification and characterization of RNA-modifying enzymes. Methods Enzymol., 425, 211–229.
12) Nagao, A., Ohara, M., Miyauchi, K., Yokobori, S.I., Yamagishi, A., Watanabe, K., & Suzuki, T. (2017) Hydroxylation of a conserved tRNA modification establishes non-universal genetic code in echinoderm mitochondria. Nat. Struct. Mol. Biol., 24, 778–782.
13) Matuszewski, M., Wojciechowski, J., Miyauchi, K., Gdaniec, Z., Wolf, W.M., Suzuki, T., & Sochacka, E. (2017) A hydantoin isoform of cyclic N6-threonylcarbamoyladenosine (ct6A) is present in tRNAs. Nucleic Acids Res., 45, 2137–2149.
14) Miyauchi, K., Kimura, S., & Suzuki, T. (2013) A cyclic form of N6-threonylcarbamoyladenosine as a widely distributed tRNA hypermodification. Nat. Chem. Biol., 9, 105–111.
15) Kang, B.I., Miyauchi, K., Matuszewski, M., D’Almeida, G.S., Rubio, M.A.T., Alfonzo, J.D., Inoue, K., Sakaguchi, Y., Suzuki, T., Sochacka, E., et al. (2017) Identification of 2-methylthio cyclic N6-threonylcarbamoyladenosine (ms2ct6A) as a novel RNA modification at position 37 of tRNAs. Nucleic Acids Res., 45, 2124–2136.
16) Salazar, J.C., Ambrogelly, A., Crain, P.F., McCloskey, J.A., & Soll, D. (2004) A truncated aminoacyl-tRNA synthetase modifies RNA. Proc. Natl. Acad. Sci. USA, 101, 7536–7541.
17) Ikeuchi, Y., Kimura, S., Numata, T., Nakamura, D., Yokogawa, T., Ogata, T., Wada, T., Suzuki, T., & Suzuki, T. (2010) Agmatine-conjugated cytidine in a tRNA anticodon is essential for AUA decoding in archaea. Nat. Chem. Biol., 6, 277–282.
18) Mandal, D., Kohrer, C., Su, D., Russell, S.P., Krivos, K., Castleberry, C.M., Blum, P., Limbach, P.A., Soll, D., & RajBhandary, U.L. (2010) Agmatidine, a modified cytidine in the anticodon of archaeal tRNA(Ile), base pairs with adenosine but not with guanosine. Proc. Natl. Acad. Sci. USA, 107, 2872–2877.
19) Mandal, D., Kohrer, C., Su, D., Babu, I.R., Chan, C.T., Liu, Y., Soll, D., Blum, P., Kuwahara, M., Dedon, P.C., et al. (2014) Identification and codon reading properties of 5-cyanomethyl uridine, a new modified nucleoside found in the anticodon wobble position of mutant haloarchaeal isoleucine tRNAs. RNA, 20, 177–188.
20) Chen, P., Crain, P.F., Nasvall, S.J., Pomerantz, S.C., & Bjork, G.R. (2005) A “gain of function” mutation in a protein mediates production of novel modified nucleosides. EMBO J., 24, 1842–1851.
21) Dumelin, C.E., Chen, Y., Leconte, A.M., Chen, Y.G., & Liu, D.R. (2012) Discovery and biological characterization of geranylated RNA in bacteria. Nat. Chem. Biol., 8, 913–919.
22) Dal Magro, C., Keller, P., Kotter, A., Werner, S., Duarte, V., Marchand, V., Ignarski, M., Freiwald, A., Muller, R.U., Dieterich, C., et al. (2018) A Vastly Increased Chemical Variety of RNA Modifications Containing a Thioacetal Structure. Angew. Chem., 57, 7893–7897.
23) Jager, G., Chen, P., & Bjork, G.R. (2016) Transfer RNA Bound to MnmH Protein Is Enriched with Geranylated tRNA–A Possible Intermediate in Its Selenation? PLoS One, 11, e0153488.
24) Sierant, M., Leszczynska, G., Sadowska, K., Komar, P., Radzikowska-Cieciura, E., Sochacka, E., & Nawrot, B. (2018) Escherichia coli tRNA 2-selenouridine synthase (SelU) converts S2U-RNA to Se2U-RNA via S-geranylated-intermediate. FEBS Lett., 592, 2248–2258.
25) Reichle, V.F., Petrov, D.P., Weber, V., Jung, K., & Kellner, S. (2019) NAIL-MS reveals the repair of 2-methylthiocytidine by AlkB in E. coli. Nat. Commun., 10, 5600.
26) Cahova, H., Winz, M.L., Hofer, K., Nubel, G., & Jaschke, A. (2015) NAD captureSeq indicates NAD as a bacterial cap for a subset of regulatory RNAs. Nature, 519, 374–377.
27) Chen, Y.G., Kowtoniuk, W.E., Agarwal, I., Shen, Y., & Liu, D.R. (2009) LC/MS analysis of cellular RNA reveals NAD-linked RNA. Nat. Chem. Biol., 5, 879–881.
28) Walters, R.W., Matheny, T., Mizoue, L.S., Rao, B.S., Muhlrad, D., & Parker, R. (2017) Identification of NAD+capped mRNAs in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA, 114, 480–485.
29) Jiao, X., Doamekpor, S.K., Bird, J.G., Nickels, B.E., Tong, L., Hart, R.P., & Kiledjian, M. (2017) 5′ End Nicotinamide Adenine Dinucleotide Cap in Human Cells Promotes RNA Decay through DXO-Mediated deNADding. Cell, 168, 1015–1027.e10.
30) Ikeuchi, Y., Kitahara, K., & Suzuki, T. (2008) The RNA acetyltransferase driven by ATP hydrolysis synthesizes N4-acetylcytidine of tRNA anticodon. EMBO J., 27, 2194–2203.
31) Ikeuchi, Y., Shigi, N., Kato, J., Nishimura, A., & Suzuki, T. (2006) Mechanistic Insights into Sulfur Relay by Multiple Sulfur Mediators Involved in Thiouridine Biosynthesis at tRNA Wobble Positions. Mol. Cell, 21, 97–108.
32) Kimura, S., Ikeuchi, Y., Kitahara, K., Sakaguchi, Y., Suzuki, T., & Suzuki, T. (2012) Base methylations in the double-stranded RNA by a fused methyltransferase bearing unwinding activity. Nucleic Acids Res., 40, 4071–4085.
33) Kimura, S., Miyauchi, K., Ikeuchi, Y., Thiaville, P.C., Crecy-Lagard, V., & Suzuki, T. (2014) Discovery of the beta-barrel-type RNA methyltransferase responsible for N6-methylation of N6-threonylcarbamoyladenosine in tRNAs. Nucleic Acids Res., 42, 9350–9365.
34) Kimura, S., Sakai, Y., Ishiguro, K., & Suzuki, T. (2017) Biogenesis and iron-dependency of ribosomal RNA hydroxylation. Nucleic Acids Res., 45, 12974–12986.
35) Kimura, S. & Suzuki, T. (2010) Fine-tuning of the ribosomal decoding center by conserved methyl-modifications in the Escherichia coli 16S rRNA. Nucleic Acids Res., 38, 1341–1352.
36) Noma, A., Kirino, Y., Ikeuchi, Y., & Suzuki, T. (2006) Biosynthesis of wybutosine, a hyper-modified nucleoside in eukaryotic phenylalanine tRNA. EMBO J., 25, 2142–2154.
37) Noma, A., Sakaguchi, Y., & Suzuki, T. (2009) Mechanistic characterization of the sulfur-relay system for eukaryotic 2-thiouridine biogenesis at tRNA wobble positions. Nucleic Acids Res., 37, 1335–1352.
38) Noma, A., Yi, S., Katoh, T., Takai, Y., Suzuki, T., & Suzuki, T. (2011) Actin-binding protein ABP140 is a methyltransferase for 3-methylcytidine at position 32 of tRNAs in Saccharomyces cerevisiae. RNA, 17, 1111–1119.
39) Sakai, Y., Kimura, S., & Suzuki, T. (2019) Dual pathways of tRNA hydroxylation ensure efficient translation by expanding decoding capability. Nat. Commun., 10, 2858.
40) Sakai, Y., Miyauchi, K., Kimura, S., & Suzuki, T. (2016) Biogenesis and growth phase-dependent alteration of 5-methoxycarbonylmethoxyuridine in tRNA anticodons. Nucleic Acids Res., 44, 509–523.
41) Soma, A., Ikeuchi, Y., Kanemasa, S., Kobayashi, K., Ogasawara, N., Ote, T., Kato, J., Watanabe, K., Sekine, Y., & Suzuki, T. (2003) An RNA-modifying enzyme that governs both the codon and amino acid specificities of isoleucine tRNA. Mol. Cell, 12, 689–698.
42) Takakura, M., Ishiguro, K., Akichika, S., Miyauchi, K., & Suzuki, T. (2019) Biogenesis and functions of aminocarboxypropyluridine in tRNA. Nat. Commun., 10, 5542.
43) Chimnaronk, S., Suzuki, T., Manita, T., Ikeuchi, Y., Yao, M., Suzuki, T., & Tanaka, I. (2009) RNA helicase module in an acetyltransferase that modifies a specific tRNA anticodon. EMBO J., 28, 1362–1373.
44) Ito, S., Akamatsu, Y., Noma, A., Kimura, S., Miyauchi, K., Ikeuchi, Y., Suzuki, T., & Suzuki, T. (2014a) A Single Acetylation of 18 S rRNA Is Essential for Biogenesis of the Small Ribosomal Subunit in Saccharomyces cerevisiae. J. Biol. Chem., 289, 26201–26212.
45) Ito, S., Horikawa, S., Suzuki, T., Kawauchi, H., Tanaka, Y., Suzuki, T., & Suzuki, T. (2014b) Human NAT10 is an ATP-dependent RNA acetyltransferase responsible for N4-acetylcytidine formation in 18 S ribosomal RNA (rRNA). J. Biol. Chem., 289, 35724–35730.
46) Arango, D., Sturgill, D., Alhusaini, N., Dillman, A.A., Sweet, T.J., Hanson, G., Hosogane, M., Sinclair, W.R., Nanan, K.K., Mandler, M.D., et al. (2018) Acetylation of Cytidine in mRNA Promotes Translation Efficiency. Cell, 175, 1872–1886.e24.
47) Taniguchi, T., Miyauchi, K., Sakaguchi, Y., Yamashita, S., Soma, A., Tomita, K., & Suzuki, T. (2018) Acetate-dependent tRNA acetylation required for decoding fidelity in protein synthesis. Nat. Chem. Biol., 14, 1010–1020.
48) Frye, M., Harada, B.T., Behm, M., & He, C. (2018) RNA modifications modulate gene expression during development. Science, 361, 1346–1349.
49) Banerjee, A.K. (1980) 5′-terminal cap structure in eucaryotic messenger ribonucleic acids. Microbiol. Rev., 44, 175–205.
50) Fan, H., Sakuraba, K., Komuro, A., Kato, S., Harada, F., & Hirose, Y. (2003) PCIF1, a novel human WW domain-containing protein, interacts with the phosphorylated RNA polymerase II. Biochem. Biophys. Res. Commun., 301, 378–385.
51) Iyer, L.M., Zhang, D., & Aravind, L. (2016) Adenine methylation in eukaryotes: Apprehending the complex evolutionary history and functional potential of an epigenetic modification. BioEssays, 38, 27–40.
52) Akichika, S., Hirano, S., Shichino, Y., Suzuki, T., Nishimasu, H., Ishitani, R., Sugita, A., Hirose, Y., Iwasaki, S., Nureki, O., et al. (2019) Cap-specific terminal N (6)-methylation of RNA by an RNA polymerase II-associated methyltransferase. Science, 363, 363.
53) Mauer, J., Luo, X., Blanjoie, A., Jiao, X., Grozhik, A.V., Patil, D.P., Linder, B., Pickering, B.F., Vasseur, J.J., Chen, Q., et al. (2017) Reversible methylation of m(6)Am in the 5′ cap controls mRNA stability. Nature, 541, 371–375.
54) Hensel, J., Duex, J.E., Owens, C., Dancik, G.M., Edwards, M.G., Frierson, H.F., & Theodorescu, D. (2015) Patient Mutation Directed shRNA Screen Uncovers Novel Bladder Tumor Growth Suppressors. Molecular cancer research. MCR, 13, 1306–1315.
55) Rozov, A., Demeshkina, N., Westhof, E., Yusupov, M., & Yusupova, G. (2015) Structural insights into the translational infidelity mechanism. Nat. Commun., 6, 7251.
56) Thiaville, P.C., Iwata-Reuyl, D., & de Crecy-Lagard, V. (2014) Diversity of the biosynthesis pathway for threonylcarbamoyladenosine (t(6)A), a universal modification of tRNA. RNA Biol., 11, 1529–1539.
57) Lin, H., Miyauchi, K., Harada, T., Okita, R., Takeshita, E., Komaki, H., Fujioka, K., Yagasaki, H., Goto, Y.I., Yanaka, K., et al. (2018) CO2-sensitive tRNA modification associated with human mitochondrial disease. Nat. Commun., 9, 1875.
58) Smith, D.G., Pal, R., & Parker, D. (2012) Measuring equilibrium bicarbonate concentrations directly in cellular mitochondria and in human serum using europium/terbium emission intensity ratios. Chemistry, 18, 11604–11613.
59) Arai, T., Ishiguro, K., Kimura, S., Sakaguchi, Y., Suzuki, T., & Suzuki, T. (2015) Single methylation of 23S rRNA triggers late steps of 50S ribosomal subunit assembly. Proc. Natl. Acad. Sci. USA, 112, E4707–E4716.
60) Ishiguro, K., Arai, T., & Suzuki, T. (2019) Depletion of S-adenosylmethionine impacts on ribosome biogenesis through hypomodification of a single rRNA methylation. Nucleic Acids Res., 47, 4226–4239.
61) Kirino, Y., Yasukawa, T., Ohta, S., Akira, S., Ishihara, K., Watanabe, K., & Suzuki, T. (2004) Codon-specific translational defect caused by a wobble modification deficiency in mutant tRNA from a human mitochondrial disease. Proc. Natl. Acad. Sci. USA, 101, 15070–15075.
62) Yasukawa, T., Suzuki, T., Ishii, N., Ohta, S., & Watanabe, K. (2001) Wobble modification defect in tRNA disturbs codon-anticodon interaction in a mitochondrial disease. EMBO J., 20, 4794–4802.
63) Kurata, S., Weixlbaumer, A., Ohtsuki, T., Shimazaki, T., Wada, T., Kirino, Y., Takai, K., Watanabe, K., Ramakrishnan, V., & Suzuki, T. (2008) Modified uridines with C5-methylene substituents at the first position of the tRNA anticodon stabilize U.G wobble pairing during decoding. J. Biol. Chem., 283, 18801–18811.
64) Morscher, R.J., Ducker, G.S., Li, S.H., Mayer, J.A., Gitai, Z., Sperl, W., & Rabinowitz, J.D. (2018) Mitochondrial translation requires folate-dependent tRNA methylation. Nature, 554, 128–132.
65) Dunbar, D.R., Moonie, P.A., Zeviani, M., & Holt, I.J. (1996) Complex I deficiency is associated with 3243G:C mitochondrial DNA in osteosarcoma cell cybrids. Hum. Mol. Genet., 5, 123–129.
66) Umeda, N., Suzuki, T., Yukawa, M., Ohya, Y., Shindo, H., Watanabe, K., & Suzuki, T. (2005) Mitochondria-specific RNA-modifying enzymes responsible for the biosynthesis of the wobble base in mitochondrial tRNAs. Implications for the molecular pathogenesis of human mitochondrial diseases. J. Biol. Chem., 280, 1613–1624.
67) Asano, K., Suzuki, T., Saito, A., Wei, F.Y., Ikeuchi, Y., Numata, T., Tanaka, R., Yamane, Y., Yamamoto, T., Goto, T., et al. (2018) Metabolic and chemical regulation of tRNA modification associated with taurine deficiency and human disease. Nucleic Acids Res., 46, 1565–1583.
68) Sturman, J.A. (1993) Taurine in development. Physiol. Rev., 73, 119–147.
69) Woo, C.C., Chen, W.C., Teo, X.Q., Radda, G.K., & Lee, P.T. (2016) Downregulating serine hydroxymethyltransferase 2 (SHMT2) suppresses tumorigenesis in human hepatocellular carcinoma. Oncotarget, 7, 53005–53017.
70) Ye, J., Fan, J., Venneti, S., Wan, Y.W., Pawel, B.R., Zhang, J., Finley, L.W., Lu, C., Lindsten, T., Cross, J.R., et al. (2014) Serine catabolism regulates mitochondrial redox control during hypoxia. Cancer Discov., 4, 1406–1417.
71) Hashizume, O., Ohnishi, S., Mito, T., Shimizu, A., Ishikawa, K., Nakada, K., Soda, M., Mano, H., Togayachi, S., Miyoshi, H., et al. (2015) Epigenetic regulation of the nuclear-coded GCAT and SHMT2 genes confers human age-associated mitochondrial respiration defects. Sci. Rep., 5, 10434.
72) Molloy, A.M. (2002) Folate bioavailability and health. Int. J. Vitam. Nutr. Res., 72, 46–52.
73) Ohsawa, Y., Hagiwara, H., Nishimatsu, S.I., Hirakawa, A., Kamimura, N., Ohtsubo, H., Fukai, Y., Murakami, T., Koga, Y., Goto, Y.I., et al.; KN01 Study Group. (2019) Taurine supplementation for prevention of stroke-like episodes in MELAS: a multicentre, open-label, 52-week phase III trial. J. Neurol. Neurosurg. Psychiatry, 90, 529–536.
74) Rikimaru, M., Ohsawa, Y., Wolf, A.M., Nishimaki, K., Ichimiya, H., Kamimura, N., Nishimatsu, S., Ohta, S., & Sunada, Y. (2012) Taurine ameliorates impaired the mitochondrial function and prevents stroke-like episodes in patients with MELAS. Intern. Med., 51, 3351–3357.
75) Ramos, J. & Fu, D. (2019) The emerging impact of tRNA modifications in the brain and nervous system. Biochim. Biophys. Acta. Gene Regul. Mech., 1862, 412–428.
76) Torres, A.G., Batlle, E., & Ribas de Pouplana, L. (2014) Role of tRNA modifications in human diseases. Trends Mol. Med., 20, 306–314.
77) Baruffini, E., Dallabona, C., Invernizzi, F., Yarham, J.W., Melchionda, L., Blakely, E.L., Lamantea, E., Donnini, C., Santra, S., Vijayaraghavan, S., et al. (2013) MTO1 mutations are associated with hypertrophic cardiomyopathy and lactic acidosis and cause respiratory chain deficiency in humans and yeast. Hum. Mutat., 34, 1501–1509.
78) Ghezzi, D., Baruffini, E., Haack, T.B., Invernizzi, F., Melchionda, L., Dallabona, C., Strom, T.M., Parini, R., Burlina, A.B., Meitinger, T., et al. (2012) Mutations of the Mitochondrial-tRNA Modifier MTO1 Cause Hypertrophic Cardiomyopathy and Lactic Acidosis. Am. J. Hum. Genet., 90, 1079–1087.
79) Fakruddin, M., Wei, F.Y., Suzuki, T., Asano, K., Kaieda, T., Omori, A., Izumi, R., Fujimura, A., Kaitsuka, T., Miyata, K., et al. (2018) Defective Mitochondrial tRNA Taurine Modification Activates Global Proteostress and Leads to Mitochondrial Disease. Cell Rep., 22, 482–496.
80) Kopajtich, R., Nicholls, T.J., Rorbach, J., Metodiev, M.D., Freisinger, P., Mandel, H., Vanlander, A., Ghezzi, D., Carrozzo, R., Taylor, R.W., et al. (2014) Mutations in GTPBP3 cause a mitochondrial translation defect associated with hypertrophic cardiomyopathy, lactic acidosis, and encephalopathy. Am. J. Hum. Genet., 95, 708–720.
81) Zeharia, A., Shaag, A., Pappo, O., Mager-Heckel, A.M., Saada, A., Beinat, M., Karicheva, O., Mandel, H., Ofek, N., Segel, R., et al. (2009) Acute infantile liver failure due to mutations in the TRMU gene. Am. J. Hum. Genet., 85, 401–407.
82) Wu, Y., Wei, F.Y., Kawarada, L., Suzuki, T., Araki, K., Komohara, Y., Fujimura, A., Kaitsuka, T., Takeya, M., Oike, Y., et al. (2016) Mtu1-Mediated Thiouridine Formation of Mitochondrial tRNAs Is Required for Mitochondrial Translation and Is Involved in Reversible Infantile Liver Injury. PLoS Genet., 12, e1006355.
83) Moriya, J., Yokogawa, T., Wakita, K., Ueda, T., Nishikawa, K., Crain, P.F., Hashizume, T., Pomerantz, S.C., McCloskey, J.A., Kawai, G., et al. (1994) A novel modified nucleoside found at the first position of the anticodon of methionine tRNA from bovine liver mitochondria. Biochemistry, 33, 2234–2239.
84) Takemoto, C., Spremulli, L.L., Benkowski, L.A., Ueda, T., Yokogawa, T., & Watanabe, K. (2009) Unconventional decoding of the AUA codon as methionine by mitochondrial tRNAMet with the anticodon f5CAU as revealed with a mitochondrial in vitro translation system. Nucleic Acids Res., 37, 1616–1627.
85) Nakano, S., Suzuki, T., Kawarada, L., Iwata, H., Asano, K., & Suzuki, T. (2016) NSUN3 methylase initiates 5-formylcytidine biogenesis in human mitochondrial tRNA(Met). Nat. Chem. Biol., 12, 546–551.
86) Kawarada, L., Suzuki, T., Ohira, T., Hirata, S., Miyauchi, K., & Suzuki, T. (2017) ALKBH1 is an RNA dioxygenase responsible for cytoplasmic and mitochondrial tRNA modifications. Nucleic Acids Res., 45, 7401–7415.
87) Van Haute, L., Dietmann, S., Kremer, L., Hussain, S., Pearce, S.F., Powell, C.A., Rorbach, J., Lantaff, R., Blanco, S., Sauer, S., et al. (2016) Deficient methylation and formylation of mt-tRNA(Met) wobble cytosine in a patient carrying mutations in NSUN3. Nat. Commun., 7, 12039.
88) Patton, J.R., Bykhovskaya, Y., Mengesha, E., Bertolotto, C., & Fischel-Ghodsian, N. (2005) Mitochondrial myopathy and sideroblastic anemia (MLASA): missense mutation in the pseudouridine synthase 1 (PUS1) gene is associated with the loss of tRNA pseudouridylation. J. Biol. Chem., 280, 19823–19828.
89) Ruggero, D., Grisendi, S., Piazza, F., Rego, E., Mari, F., Rao, P.H., Cordon-Cardo, C., & Pandolfi, P.P. (2003) Dyskeratosis Congenita and Cancer in Mice Deficient in Ribosomal RNA Modification. Science, 299, 259–262.
90) Jack, K., Bellodi, C., Landry, D.M., Niederer, R.O., Meskauskas, A., Musalgaonkar, S., Kopmar, N., Krasnykh, O., Dean, A.M., Thompson, S.R., et al. (2011) rRNA Pseudouridylation Defects Affect Ribosomal Ligand Binding and Translational Fidelity From Yeast to Human Cells. Mol. Cell, 44, 660–666.
91) Steinthorsdottir, V., Thorleifsson, G., Reynisdottir, I., Benediktsson, R., Jonsdottir, T., Walters, G.B., Styrkarsdottir, U., Gretarsdottir, S., Emilsson, V., Ghosh, S., et al. (2007) A variant in CDKAL1 influences insulin response and risk of type 2 diabetes. Nat. Genet., 39, 770–775.
92) Wei, F.Y., Suzuki, T., Watanabe, S., Kimura, S., Kaitsuka, T., Fujimura, A., Matsui, H., Atta, M., Michiue, H., Fontecave, M., et al. (2011) Deficit of tRNALys modification by Cdkal1 causes the development of type 2 diabetes in mice. J. Clin. Invest., 121, 3598–3608.
93) Anderson, S.L., Coli, R., Daly, I.W., Kichula, E.A., Rork, M.J., Volpi, S.A., Ekstein, J., & Rubin, B.Y. (2001) Familial dysautonomia is caused by mutations of the IKAP gene. Am. J. Hum. Genet., 68, 753–758.
94) Karlsborn, T., Tukenmez, H., Chen, C., & Bystrom, A.S. (2014) Familial dysautonomia (FD) patients have reduced levels of the modified wobble nucleoside mcm(5)s(2)U in tRNA. Biochem. Biophys. Res. Commun., 454, 441–445.
95) Yoshida, M., Kataoka, N., Miyauchi, K., Ohe, K., Iida, K., Yoshida, S., Nojima, T., Okuno, Y., Onogi, H., Usui, T., et al. (2015) Rectifier of aberrant mRNA splicing recovers tRNA modification in familial dysautonomia. Proc. Natl. Acad. Sci. USA, 112, 2764–2769.
96) Cohen, J.S., Srivastava, S., Farwell, K.D., Lu, H.M., Zeng, W., Lu, H., Chao, E.C., & Fatemi, A. (2015) ELP2 is a novel gene implicated in neurodevelopmental disabilities. Am. J. Med. Genet. A., 167, 1391–1395.
97) Simpson, C.L., Lemmens, R., Miskiewicz, K., Broom, W.J., Hansen, V.K., van Vught, P.W., Landers, J.E., Sapp, P., Van Den Bosch, L., Knight, J., et al. (2009) Variants of the elongator protein 3 (ELP3) gene are associated with motor neuron degeneration. Hum. Mol. Genet., 18, 472–481.
98) Abbasi-Moheb, L., Mertel, S., Gonsior, M., Nouri-Vahid, L., Kahrizi, K., Cirak, S., Wieczorek, D., Motazacker, M.M., Esmaeeli-Nieh, S., Cremer, K., et al. (2012) Mutations in NSUN2 cause autosomal-recessive intellectual disability. Am. J. Hum. Genet., 90, 847–855.
99) Davarniya, B., Hu, H., Kahrizi, K., Musante, L., Fattahi, Z., Hosseini, M., Maqsoud, F., Farajollahi, R., Wienker, T.F., Ropers, H.H., et al. (2015) The Role of a Novel TRMT1 Gene Mutation and Rare GRM1 Gene Defect in Intellectual Disability in Two Azeri Families. PLoS One, 10, e0129631.
100) Freude, K., Hoffmann, K., Jensen, L.R., Delatycki, M.B., des Portes, V., Moser, B., Hamel, B., van Bokhoven, H., Moraine, C., Fryns, J.P., et al. (2004) Mutations in the FTSJ1 gene coding for a novel S-adenosylmethionine-binding protein cause nonsyndromic X-linked mental retardation. Am. J. Hum. Genet., 75, 305–309.
101) Shaheen, R., Al-Salam, Z., El-Hattab, A.W., & Alkuraya, F.S. (2016) The syndrome dysmorphic facies, renal agenesis, ambiguous genitalia, microcephaly, polydactyly and lissencephaly (DREAM-PL): Report of two additional patients. Am. J. Med. Genet. A., 170, 3222–3226.
102) Alazami, A.M., Hijazi, H., Al-Dosari, M.S., Shaheen, R., Hashem, A., Aldahmesh, M.A., Mohamed, J.Y., Kentab, A., Salih, M.A., Awaji, A., et al. (2013) Mutation in ADAT3, encoding adenosine deaminase acting on transfer RNA, causes intellectual disability and strabismus. J. Med. Genet., 50, 425–430.
103) Riazuddin, S., Hussain, M., Razzaq, A., Iqbal, Z., Shahzad, M., Polla, D.L., Song, Y., van Beusekom, E., Khan, A.A., Tomas-Roca, L., et al.; UK10K. (2017) Exome sequencing of Pakistani consanguineous families identifies 30 novel candidate genes for recessive intellectual disability. Mol. Psychiatry, 22, 1604–1614.
104) Richard, E.M., Polla, D.L., Assir, M.Z., Contreras, M., Shahzad, M., Khan, A.A., Razzaq, A., Akram, J., Tarar, M.N., Blanpied, T.A., et al. (2019) Bi-allelic Variants in METTL5 Cause Autosomal-Recessive Intellectual Disability and Microcephaly. Am. J. Hum. Genet., 105, 869–878.
105) Freund, I., Eigenbrod, T., Helm, M., & Dalpke, A.H. (2019) RNA Modifications Modulate Activation of Innate Toll-Like Receptors. Genes (Basel), 10, 92.
106) Vogel, P. & Stafforst, T. (2019) Critical Review on Engineering Deaminases for Site-Directed RNA Editing. Curr. Opin. Biotechnol., 55, 74–80.
著者紹介Author Profile
 鈴木 勉(すずき つとむ)
鈴木 勉(すずき つとむ)東京大学大学院工学系研究科教授.博士(理学).
略歴1996年東京工業大学にて博士取得.2008年より現職.
研究テーマと抱負タンパク質合成のメカニズムとRNA修飾の機能に興味があります.RNAが持つ分子的な側面をあぶりだすことで生命現象の本質に迫るような研究を意識しています.
ウェブサイトhttp://rna.chem.t.u-tokyo.ac.jp/index.html
趣味プールで泳ぐこと.