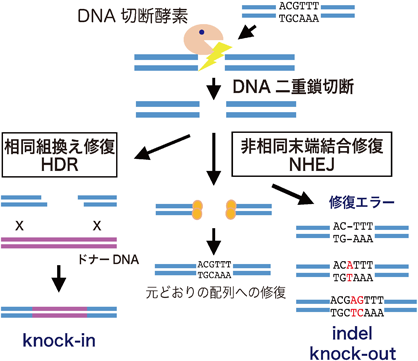
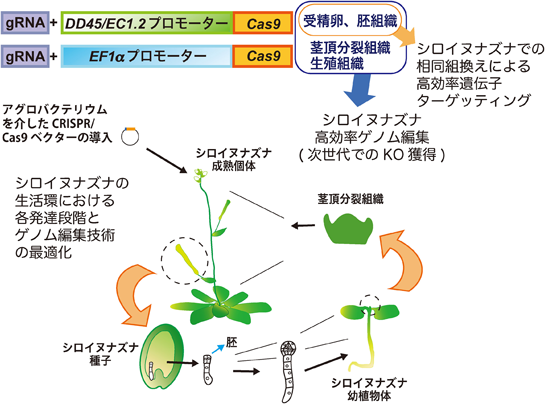
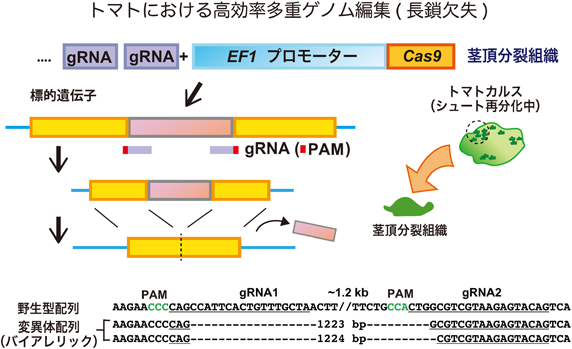
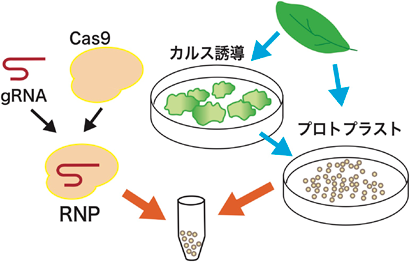
植物におけるゲノム編集技術と応用の最新展開Perspectives on the application of plant genome editing
徳島大学大学院社会産業理工学研究部准教授Faculty of Bioscience and Bioindustry, Tokushima University ◇ 〒770–8513 徳島県徳島市南常三島町2–1 ◇ 2–1 Minami-josanjima, Tokushima-city, Tokushima 770–8513, Japan
発行日:2020年6月25日Published: June 25, 2020