近年先進国ではアレルギー疾患の有病率が急増しており,日本では人口の半数近くが喘息,花粉症,アトピー性皮膚炎,食物アレルギーなど,何らかのアレルギー疾患を持っている.現在のところ完治が困難な慢性疾患で,治療は対症療法が中心であり,根治的治療法や予防法は確立していない.アレルギー疾患の発症には遺伝的素因,環境因子,抗生物質の使用,食生活,運動,その他の要因が絡み,そのメカニズムは複雑である.アレルギー反応において,マスト細胞が中心的な役割を持ち,IgEとアレルゲンによってその活性化が起こることはよく知られている.しかし,アレルギー発症の機序を理解するためにはアレルゲン,IgE,マスト細胞だけでなく,広く免疫現象を知る必要がある.本稿では我々の最近の研究を含め,アレルギーの発症機序におけるマスト細胞の活性化と,アレルギー疾患の病態や新たな治療法について述べたい.
きょうだいの多い子供ではアレルギー性鼻炎とアトピー性皮膚炎が少ないという観察から始まり,年上の同胞が持ち込む多くの微生物にさらされているからという考え,衛生仮説(hygiene hypothesis)が1989年に提唱された1).アレルギー患者の急増は,乳幼児期に微生物に曝露されることが減り,免疫系が抗原に対して正しく反応することを学習できないためだという考えである.また,帝王切開で生まれた子供は喘息やアレルギー疾患にかかりやすいこと,逆にペットを飼っていることや農場で育つことは喘息発症率と負の相関があることが報告されている.一方で,喘息重症化には,ライノウイルス,RSウイルス等のウイルス感染が引き金になること2),また食物アレルギーの発症には,アトピー性皮膚炎のように皮膚のバリアー機能が壊れて,皮膚からアレルゲンに感作されることがわかってきた3).なお,非衛生的環境で起こる寄生虫感染症に対する生体防御反応は,IgEとマスト細胞が重要な役割を果たす2型の炎症である.
新生児期からいろいろな微生物にさらされることにより,免疫細胞は学習し,抗原に対して適切に反応することを学ぶ.常在細菌叢を持たない無菌マウスや抗生物質で常在細菌叢が減少ないし変化したマウスでは,アレルギー発症に中心的な役割を持つ血中免疫グロブリンE(immunoglobulin E:IgE)が普通のマウスに比べてきわめて高くなる4, 5).IgE産生の抑制には制御性T細胞(regulatory T cell:Treg)が関与している.Tregは胸腺由来のtTreg(またはnatural Treg)と正常細菌叢にさらされて新生児期にできる末梢性pTreg(またはinduced Treg)がある.pTregはtTregと異なり,Neuropilin−1とよばれるマーカーが陰性であり,腸管においてはRORγtという転写因子を発現している.pTregは無菌マウスにおいて,食事抗原によって小腸に誘導され6),IgE産生を抑制する7).腸管のRORγt+ pTreg細胞は正常細菌叢,レチノイン酸,短鎖脂肪酸によって誘導されて,2型の免疫反応を抑制する8).離乳期のマウスにおいて,母親のミルク成分の変化,特に上皮成長因子(epidermal growth factor:EGF)の減少により,腸管の正常細菌叢の変化が起こることで,RORγt+ pTreg細胞が誘導される.この時期の免疫系の変化は離乳反応(weaning reaction)と呼ばれていて9),無菌マウスや抗生物質投与マウスはこの反応が起こらない.その結果,2型炎症(オキサゾロン誘発性大腸炎)だけでなく,他のタイプの炎症反応も過剰になる.新生児期の短期間のうちに,正常細菌叢の変化によってpTregが誘導されることは,肺でも同じである.マウス肺では,生後2週間でPD-L1依存的に,pTregが誘導される10).
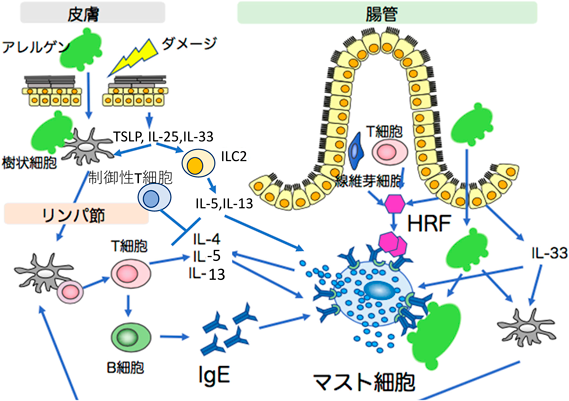
アレルギーとは,本来生体にとって無害な抗原に対して,異常な免疫反応を起こすことである.IgE依存性のアレルギー疾患の発症は,さまざまな免疫細胞が関与し,発症機構は複雑であるが,そのメカニズムを概観する(図1).アレルギー疾患にかかるには,個体はまず,抗原(アレルゲン)にさらされ,これに対するIgE抗体を作る.これを感作という.食物アレルギーでは,感作は主に皮膚を介して起こると考えられている.皮膚のバリアーが壊れ,そこからアレルゲンが進入して,これが樹状細胞(dendritic cell:DC)に捉えられる.あるいは,機械的刺激でダメージを受けると表皮細胞からthymic stromal lymphopoietin(TSLP)やIL-25, IL-33などのサイトカインが放出される.これらの上皮細胞由来サイトカインはいろいろな細胞に働きかけるが,特に2型自然リンパ球(innate lymphoid cell:ILC2)11)を刺激して,IL-5, IL-13などのサイトカインを産生・放出する12).TSLPはDCを活性化し,アレルゲンを取り込んだDCは,局所のリンパ節に移動する.ここでDCは主要組織適合遺伝子複合体クラスII分子(MHC II)によって抗原を提示し,CD40を介した刺激とで,CD4 T細胞を2型ヘルパーT細胞(Th2 cell)に分化させ活性化する.活性化されたTh2細胞はIL-4, IL-5, IL-13などのサイトカインを産生・放出する.CD4 T細胞の活性は,制御性T細胞(Treg)によってコントロールされている.2型の免疫は,pTregによって制御されている13).また,活性化されたCD4 T細胞はリンパ節の濾胞においてT follicular helper cell(Tfh)に分化し,IL-4を分泌してB細胞を刺激し,胚中心(germinal center)において,B細胞の増殖,クラススイッチ組換え(class switch recombination:CSR),体細胞超変異(somatic hypermutation:SHM)を引き起こして,高親和性の抗原特異的なIgEの産生を起こす.従来,Th2由来のIL-4がIgE産生に重要と考えられてきたが14),IgE産生に必要なIL-4はTfh由来であることは寄生虫感染モデルで証明されている15, 16).TfhによるIL-4の発現には,Th2細胞で起こる2型サイトカインの発現とは異なる機序が働き,IL-5やIL-13の発現は抑えられる17, 18).Kitaらの研究により,喘息や食物アレルギーモデルにおいてもTfhがIgE反応を引き起こすIL-4産生細胞であることが明らかになっている19, 20).リンパ節や脾臓でIL-4を産生する細胞の大半はTfhで,Th2は末梢の組織に分布している21).TfhはIL-21も産生するが,IgEへのCSRはTfhのIL-21産生レベルが低いとき,ないしB細胞のIL-21受容体の感度が低いときに起こると思われる.また,アナフィラキシーを起こすようなIgE産生には,IL-13を作るTfh細胞(Tfh13)が必要であるが22),Tfh13の活性はfollicular regulatory T cell(Tfr)によって制御されている23).分泌されたIgEは血中を運ばれて,種々の組織中のマスト細胞や血中の好塩基球に高親和性IgE受容体(FcεRI)を介して結合する.これが感作の過程である.食物アレルギーでは,食物由来のアレルゲンがIgEに結合し,FcεRIを凝集させることによって,小腸のマスト細胞を活性化する.また,腸管の上皮細胞や,その他の細胞から,後述のHRFというサイトカイン様のタンパク質が放出され,これもマスト細胞上のIgEに結合し,マスト細胞を活性化する24).アレルギー症状の発現にはアレルゲン,IgE, FcεRI,マスト細胞または好塩基球が重要であり,急性期(分のオーダー)には活性化されたマスト細胞からヒスタミン,セロトニンなどの細胞内顆粒にあるアレルギー誘引因子が放出され,血管の拡張,血管透過性の上昇を起こす.それに続く時期(時間のオーダー)ではマスト細胞由来のサイトカイン(TNFなど)によって顆粒球などの免疫細胞が反応局所に動員されることによる症状が出る.さらに,好塩基球により遅い(日単位)皮膚反応も起こることがある.しかし,ピーナッツアレルギーにみられるように,ごく微量のアレルゲンで発症するアレルギー発症のメカニズムなど,本当にはよくわかっていない現象もある.以下の節でマスト細胞とIgEのアレルギーにおける役割について述べたい.
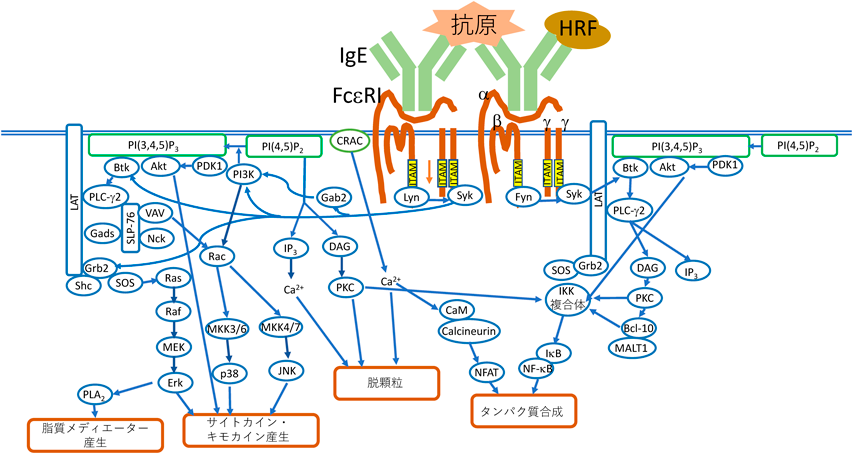
マスト細胞は,皮膚,気道粘膜,腸管粘膜など,主に生体が外界と接する組織に多くみられる骨髄由来の細胞で,寄生虫や細菌感染症に対する免疫応答という生体防御機能を担っている.マスト細胞は,多くの細胞内顆粒を持ち,顆粒の中にはヒスタミン,セロトニン,TNFαなどのすでに合成されたサイトカインやタンパク質分解酵素などが含まれている.マスト細胞(または好塩基球)の表面には高親和性IgE受容体FcεRIが発現している.これはIgEのFc部分に結合するα鎖,シグナルを増幅するβ鎖,それにシグナルを下流に伝える役割を持ちジスルフィド結合によって結合された2本のγ鎖からなる四量体である.FcεRIに抗原特異的IgEが結合し,さらにこのIgEに多価の特異抗原(アレルゲン)が結合すると複数のFcεRIが近接し[架橋(cross-linking)と呼ばれる],マスト細胞の活性化が起こる25)(図2).以下にシグナル伝達の概観を説明するが,詳細はいくつかの総説26)にあたっていただきたい.まず,β鎖に結合しているSrcファミリーのチロシンキナーゼLyn(Fyn, Hck, FgrなどのSrcファミリーのチロシンキナーゼも)が活性化されて,β鎖とγ鎖が持つimmunoreceptor tyrosine-based activation motif(ITAM)のチロシン残基をリン酸化する.チロシンリン酸化されたγ鎖には,Sykが結合し,LynやSykによってリン酸化されて活性化される.Sykはさらに,アダプター分子であるLATやSLP-76,チロシンキナーゼBtk,それにVAVなどをチロシンリン酸化する.リン酸化されたLATはGrb2を引き寄せ,そこにSOSが結合し,Rasを介してErkが活性化されて,サイトカインの産生が起こる.またErkからPLA2の活性化を介して脂質メディエーターが産生される.リン酸化されたSLP-76やVAVはRacを活性化して,さらにJNKとp38を活性化し,この経路もサイトカイン産生に関与する.LATにはBtkによってリン酸化されたPLC-γ1やPLC-γ2も結合し,PLC-γ1/2がphosphatidylinositol 4,5-bisphosphate(PIP2)を加水分解して,diacylglycerol(DAG)とinositol-3-phosphate(IP3)ができる.これらはさらにそれぞれPKC(特にPKCβII)を活性化し,あるいはCa2+を動員して脱顆粒に至らしめる.またPIP2はPI3 kinase(PI3K)によってリン酸化されてphosphatidylinositol 3-phosphate[PI(3,4,5)P3]となり,ここにBtk, PDK1, Aktなどが結合する.PDK1はAktをリン酸化して活性化し,活性化されたAktはIKKを介してNF-κBを活性化し,新たなタンパク質合成が誘導される.脱顆粒は,FcεRIが架橋されてから数秒から数分で起こり,顆粒に含まれているヒスタミン,セロトニンなどのメディエーターや,TNFαなどのサイトカインが放出される.放出された顆粒は,壊れずにそのまま局所のリンパ節に移動し,そこで免疫反応を引き起こすことも知られている27).さらに数時間後には,上記の経路の活性化により,サイトカインや脂質メディエーターの新たな産生が起こり,これらが放出される.こうして全身あるいは局所でアレルギー炎症が起こる.以上のシグナル伝達の随所を阻害する物質が,アレルギー疾患の治療薬として,研究開発されている.現在のところ実用化され広く使われているのは,IgEとFcεRIの結合を阻害するオマリズマブ(omalizumab)である.これは難治性気管支喘息や慢性特発性蕁麻疹の治療に使われている.
従来,IgEが結合しているだけのマスト細胞は,休止していると考えられていたが,我々の研究により,IgEはFcεRIに結合するだけで,抗原の存在なしでもマスト細胞の生存能その他の生物活性に影響を及ぼしていることが明らかになった.すなわち抗原がなくても,単量体IgEのみがFcεRIに結合するだけで,マスト細胞の生存が延長する28).時を同じくして,Kalesnikoffらも単量体IgEの結合によるマスト細胞の生存延長を認めて発表した29).我々の研究室では,単量体IgEの結合だけでは,マスト細胞からの脱顆粒もサイトカイン産生も認められなかったが,Kalesnikoffらは単量体IgEの結合により,マスト細胞がサイトカインを産生放出することも認めた.この違いが何によるのかを調べるために,多数のモノクローナルIgEのマスト細胞活性化能を調べたところ,我々はこの違いが実験に使われたIgEの違いによることを見いだした.そして,単量体IgEのサイトカイン産生能に応じて,高サイトカイン産生性IgE(highly cytokinergic IgE:HC IgE)および低サイトカイン産生性IgE(poorly cytokinergic IgE:PC IgE)と名づけて報告した30).HC IgEはマスト細胞上のFcεRIに結合すると,細胞の生存延長のみならず,サイトカイン産生,ヒスタミン放出,細胞移動,細胞接着など,IgEで感作されたFcεRIが抗原によって架橋されることで起こる,ほとんどすべてのマスト細胞の生物的反応を起こすことができる.しかしPC IgEはマスト細胞の生存を延長させることはできるが,それ以外のマスト細胞機能を活性化することはできない.以上により我々はIgEには多様性があることを証明した31).
1979年に,血中の単核球に,好塩基球のヒスタミン放出を促す因子があることが報告された32).この物質はヒスタミン放出因子(histamine-releasing factor:HRF)と呼ばれ,その後長く,研究が続けられてきた.HRFは1995年にMacDonaldらによってアミノ酸172個からなるタンパク質としてクローン化された33).HRFはアレルギー疾患患者の鼻汁,水疱内容液,喘息患者の気管支肺胞洗浄液などに含まれ,ある種のアトピー患者由来のIgEが結合した好塩基球を刺激して,ヒスタミン放出のみならず,IL-4やIL-13など2型のサイトカインを産生させる生理活性がある.HRFと結合して好塩基球を活性化するIgEはIgE+,活性化できないものはIgE-と分類されていた34, 35).我々はIgE+がHC IgE, IgE-はPC IgEに相当するのではないかと考え,その機序の解明を試みた.この過程で,タンパク質の同定以来15年あまりにわたって不明であったHRFの受容体を発見し,これにより,HRFによるマスト細胞活性化の機序を解明し,さらにHRFの阻害物質を発見し,これがマスト細胞の活性化を抑制し,また喘息および食物アレルギーのマウスモデルで,この抑制剤が発症の予防効果と治療効果があることを認めて報告した.以下にその詳細を述べる.
HRFはすべての細胞に発現しており,進化の過程でよく保存されたタンパク質である.細胞内機能と細胞外機能がある.細胞内分子としてのHRFはtranslationally-controlled tumor protein(TCTP)やfortilinという名称で知られ,細胞の増殖,細胞周期,悪性化,DNA修復など,細胞の生存に不可欠の細胞内機能を担っている36, 37).このため,HRF遺伝子欠損マウスは胎生期に死亡する38–40).アレルギーに関与する細胞外分子としてはHRFの名称が使われ,上記のとおり,ある種のIgEに感作されたマスト細胞や好塩基球を刺激して,ヒスタミンやIL-4やIL-13などのサイトカインの分泌をうながすことが知られている.
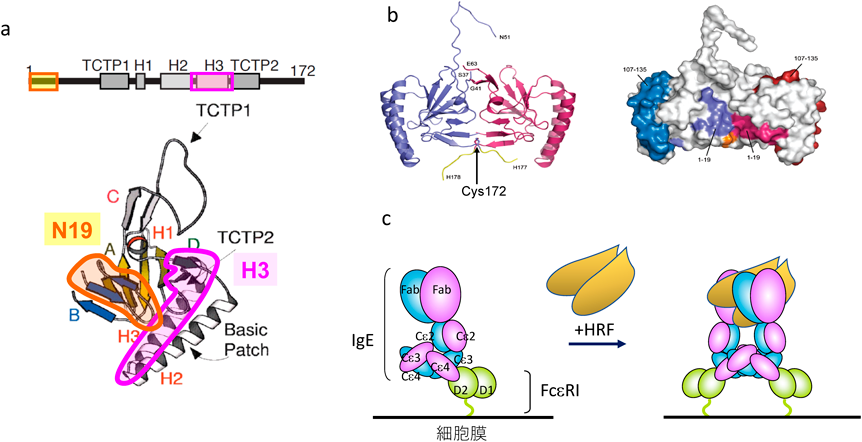
我々は以前に報告されたIgE+またはIgE−の本態を解明するために,20種類のモノクローナルIgEについてHRFと結合するかどうか調べてみた41).そのうちおよそ25%のIgEが,Fabドメインを介してHRFのN末端の19アミノ酸および,三つあるαヘリックス構造の一つ,H3ドメインと直接結合することがわかった(図3a).さらにモノクローナルIgGも,調べたクローンのうち25~30%が,IgEと同様にFabを介してHRFのN末端の19アミノ酸およびH3ドメインと結合することが明らかになった.以上より,HRFと結合するIgEおよびIgGを,それぞれHRF反応性IgEまたはIgG(HRF-reactive IgE/IgG)とHRF非反応性IgEまたはIgG(HRF-nonreactive IgE/IgG)と名づけた.ただし,当初の我々の仮説と異なり,HRF反応性IgE=HC IgE, HRF非反応性IgE=PC IgEではなかった.HRFはIgEとは結合しないという報告がなされていたが42),以前の報告はHRF非反応性IgEを用いて結合を調べたためと考えられる.
その後,HRFはC末端のシステイン(C172)を介するジスルフィド結合により,二量体を形成することがわかった.結晶化されたHRFの三次元構造解析により,二量体を形成したHRFでは,IgEとの結合部位は,結晶の表面に位置していることも明らかになった43)(図3b).1分子のHRFはIgE 2分子と結合できることから,二量体のHRFは4分子のIgEと結合することができる.理論的には,これによりHRFはFcεRIを架橋しマスト細胞または好塩基球を活性化することができる.我々は,実際に単量体ではなく二量体HRFが,IgEで感作されたマスト細胞を活性化することを証明した(図3c).以上より,HRF反応性IgEが,HRFの受容体として機能することがわかった.
以上の情報をもとに我々は数種類のHRF阻害剤を作った.一つは,HRFの結合部位であるN末端19アミノ酸やH3を大腸菌で作製したGSTとの複合タンパク質(GST-N19とGST-H3)と,これらの部位の合成ペプチドを修飾したもの(N19 peptide)である.これらはIgEとの結合を阻害することによりマスト細胞の活性化を抑制する.もう一つはHRFの二量体形成を阻害するものである.HRFには二つのシステイン残基があるが(C28, C172),この二つをアラニンに変えた変異体(HRF-2CA)はシステインを介するジスルフィド結合による二量体を形成することができない.単量体HRFはマスト細胞を活性化できないため,HRF-2CAはHRFの阻害剤として機能する.以下に述べるように,我々はこれらの阻害剤を用いて,アレルギー疾患の病態形成においてHRFが関与しているかどうか,さらにこれらの阻害剤がアレルギー疾患の予防や治療に有効かどうかを調べた.
喘息は,慢性のアレルギー性呼吸器疾患で,気道の炎症,抗原に対する気道の過剰反応(airway hyperresponsiveness:AHR),可逆性の気道閉塞などの特徴がある.アメリカでは人口のおよそ8%が喘息に罹患しており,年間4000~5000人が喘息発作で亡くなる.小児では罹患率は9%にのぼり,小児慢性疾患で最も患者数が多い.近年,ステロイドの吸入療法により,症状軽減や発作の予防が進んできたが,根治療法はない.
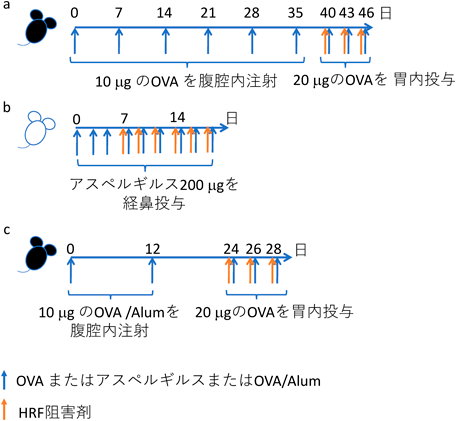
マウス喘息モデルは,その病態形成のメカニズムによりいくつもの方法が確立されている.すなわちマスト細胞依存型,部分的マスト細胞依存型,T細胞依存型などである.我々は,この三つのモデルを用いて,アレルギー性喘息におけるHRFの関与を調べた41)(図4).マスト細胞依存性モデルでは,抗原[卵白アルブミン(ovalbumin:OVA)]で感作したマウスに,抗原で刺激する直前にHRF阻害剤(GST-HRF)を投与すると,気道炎症の発症がほぼ完全に抑制された.アスペルギルスを抗原とする部分的マスト細胞依存性のモデルでも,GST-HRF投与による気道炎症の抑制が認められたが,効果はマスト細胞依存型ほどではなかった.T細胞依存型のモデルではGST-HRFの効果はまったくみられなかった.以上より,マスト細胞が関与する喘息の発症にはHRFが関与していることがわかった.さらに,ヒトの喘息患者および好酸球性肺炎患者の気管支洗浄液を調べたところ,健常者に比べてHRFの発現量が増加していることが認められた.また小児喘息の入院患者では,別の疾患で入院中の小児患者(主に感染症患者)に比べて血中のHRF反応性IgEが増加していた(未発表).ただし血中HRFおよびHRF反応性IgGの発現量には差がなかった.
以上より,喘息モデルにおいて,HRFがその病態形成に関与していることが明確に示された.またHRFは複数の喘息モデルにおいて,IgEを介して2型炎症反応を増進することが証明された.
近年,先進国では食物アレルギー患者が急増している.アメリカでは1500万人に食物アレルギーがあり,3歳以下の子供の6~8%,成人の4%が罹患している.よく問題になるアレルゲンは,ピーナッツ,アーモンドなどの木の実(tree nuts),魚,エビなどの甲殻類,鶏卵,ミルクなどである.症状はさまざまで,軽度では皮膚のかゆみ,蕁麻疹程度であるが,腹痛,嘔吐や下痢を発症し,重篤な場合はアナフィラキシーを起こし死に至る.アレルゲンの経口投与または舌下投与による免疫療法(減感作療法)が有効な患者もいるが,今のところ根本的な治療法はない(ただし本年初めにピーナッツアレルギーに対する経口投与による免疫療法剤が米国FDAによって承認された).発症メカニズムは多様で,IgE依存性,IgE非依存性・細胞免疫依存性,および複合性に分けられる.我々が用いたマウスのIgE依存性食物アレルギーモデルでは,下痢と2型炎症が起こる.これらの症状を起こすのにマスト細胞,特にIL-9産生性粘膜型マスト細胞(IL-9-producing mucosal mast cell:MMC9)44)が重要である.マスト細胞の活性化にはIgEとFcεRIが必要で,活性化マスト細胞からセロトニン,血小板活性化因子(platelet activating factor:PAF)が放出されて症状を引き起こす45).
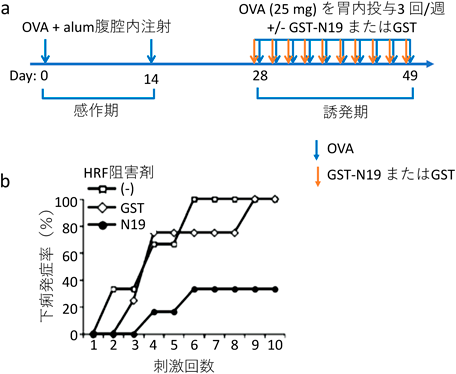
我々は卵白アルブミン(OVA)に対する食物アレルギーのモデルにおいて,HRFが発症に関与すること,またHRFの阻害剤が発症を予防することを証明した24).BALB/cマウスをアルミニウム塩(Alum)のアジュバントとともにOVAで2回感作し,その後週に3回OVAを経口投与して刺激すると4回目から6回目の刺激から下痢を発症し始める(図5a).このとき,OVAの投与30分前にHRF阻害剤(GST-OVAまたはHRF-2CA)を経口投与すると,60%以上のマウスで下痢の発症が抑制された(図5b).また小腸でのマスト細胞集積や2型サイトカインの産生も抑制された.
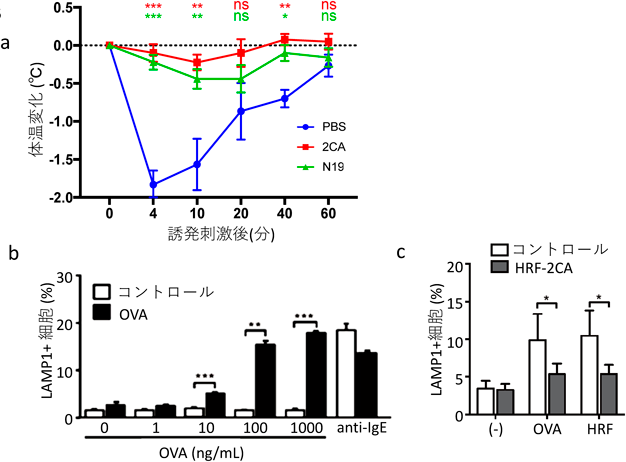
従来,マウスの食物アレルギーモデルは下痢を指標として評価されてきたが,下痢を数値化して客観的に判定することは難しい.我々は最近,ヒト用の赤外線体温計を用いてアナフィラキシーにおけるマウスの体温を測定し評価する方法を確立した46)(図6a).ビデオでみるマウスの行動と体温の間には強い相関があり,体温低下はマウスの苦痛を含めたアレルギー反応を,客観的に測定するよい指標となる.この方法を利用してマウスの食物アレルギー反応におけるHRF阻害剤の効果を調べた.抗原(OVA)で感作したマウスに抗原を経口投与すると,投与後4分から20分にかけて体温の低下が起こるが,抗原投与30分前にHRF阻害剤を経口投与すると,N19ペプチドでもHRF-2CAでも,抗原刺激による体温低下がほとんどみられなかった.
この食物アレルギーモデルにおけるマスト細胞の役割を調べるため,マウスの腸管からマスト細胞を洗い出し,ex vivoで抗原刺激した.OVAで感作し,OVAで刺激してアレルギー症状を引き起こしたマウスの腸管由来のマスト細胞は,刺激したOVAの量依存性に脱顆粒を起こしたが,感作せずにOVAで刺激したマウス由来のマスト細胞では,脱顆粒は認められなかった(図6b).また,このOVAでアレルギー症状を起こしたマウスの腸管由来のマスト細胞をOVAまたは二量体HRFで刺激すると,活性化されて脱顆粒が起こるが,この活性化は,HRF阻害剤によって抑制された(図6c).
我々は二量体および多量体のHRFを検知するELISA法を確立した.このELISAを用いて小腸のHRFを調べたところ,アレルギーを発症したマウスの小腸では,二量体および多量体のHRFが増加していることがわかった.マウスにHRF阻害剤を投与すると,二量体および多量体HRFの増加は抑制された.
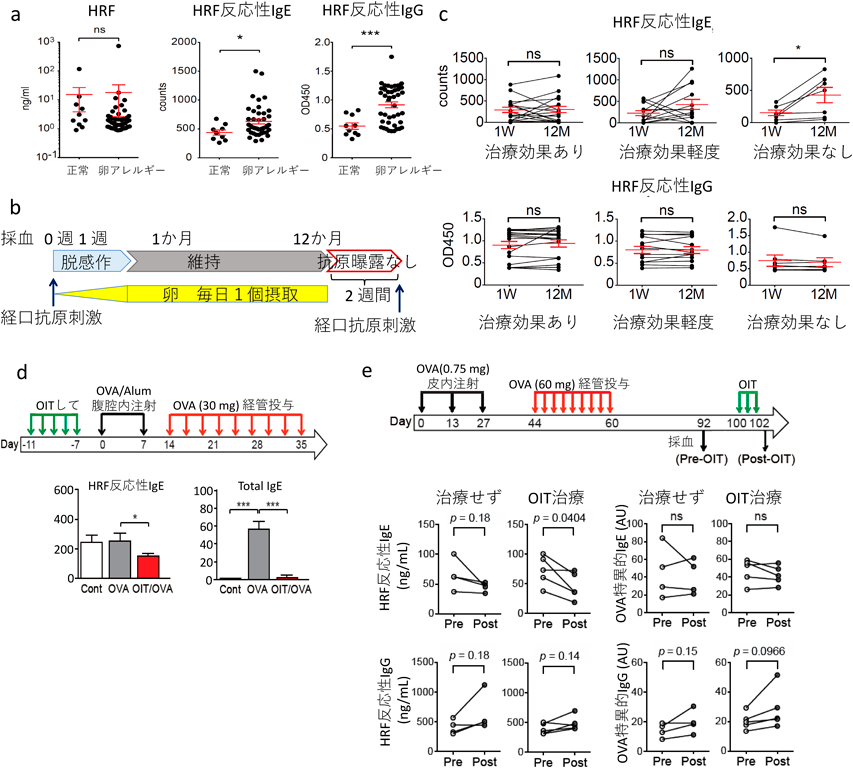
さらに,ヒトの卵アレルギー患者では,健常人に比べて血中HRF反応性IgEおよびHRF反応性IgGが増加していることもわかった24)(図7a).この患者グループは,図7bに示すプロトコールに従い,急速経口免疫療法(rapid oral immunotherapy:OIT)を受けた47).患者が重篤な症状を起こさずに卵1個が食べられるようになるまで減感作するのに,およそ2週間かかった.その後,12か月にわたって毎日卵を1個ずつ食べ続け,さらに次の2週間,卵を摂取しない休息期間を挟み,その後に卵を食べさせてOITの治療効果を判定した.その結果,80%の患者にOITの効果が認められたが,19%の患者には効果がなかった.効果があったグループと無効グループで,治療前後の血中HRFおよびHRF反応性IgE, HRF反応性IgGを測定し比較してみたところ,無効だったグループでは,有効だったグループに比べてOIT治療後のHRF反応性IgEが有意に高かった24)(図7c).次に,上記のOITモデルがマウスでも使えるかどうかを二つのプロトコールで調べてみた.一つ目は,古典的な免疫寛容誘導モデルで,比較的大量のOVAでOIT治療し,その後OVAで感作し,刺激した(図7d).OITで治療したグループでは,予想どおりHRF反応性IgEが減少したが,ヒトの治療後とは異なり,総IgEの産生も激減した.したがって,これはヒトのOITのよいモデルとはいえない.次に,まずマウスにOVAを皮内注射して感作し,さらにOVAを経口投与で刺激して食物アレルギーを起こした後,32日間の休養期間を挟んで,OITを行った(図7e).この条件下では,OITを受けたマウスは,治療後HRF反応性IgEが低下したが,OVA特異的IgEは治療前後で変化がなかった.すなわち,この方法はヒトのOITのモデルとなることがわかった.
以上およびその他のデータより,食物アレルギーにおけるHRFの役割について明らかになったことを以下にまとめ,列記する.
- HRF阻害剤の予防的投与により,下痢,低体温,活動性,2型炎症が抑制される.
- HRF阻害剤は,感作期ではなく,誘発期に効果を発揮する.
- OVA誘発性食物アレルギーにおいて,二量体または多量体のHRFが小腸で増加する.HRF阻害剤はその増加を抑える.
- 小腸のマスト細胞はOVAによって活性化されるが,HRF阻害剤で抑制される.
- HRF阻害剤は,選択的にマスト細胞に結合する.
- 経口減感作療法(OIT)後にHRF反応性IgEが増加する例では,減感作が起こらない.
さらに,好塩基球はOVA誘発性食物アレルギーにおいて,IL-4を産生し,腸管内マスト細胞数を増やし,誘発期に重要であることも示された48).
本稿ではアレルギー疾患の発生機序とマスト細胞の役割について概観した.冒頭に,アレルギー疾患の病態形成には,アレルゲン,IgE, FcεRI,マスト細胞,それにヒスタミン,セロトニンなどのアレレルギー誘引性メディエーターのみならず,その他の要素も必要ではないかと述べた.HRFは抗原によるマスト細胞の活性化を増強する.すなわちごく微量の抗原でもHRFの存在下ではアレルギー反応を引き起こしうる.また,抗原なしにHRFだけでもFcεRIを介してマスト細胞を活性化する.HRFの阻害剤は喘息と食物アレルギーの動物モデルで発症抑制に有効であること,経口投与で有効であること,個々のアレルゲンとは関係なく治療できることなどから,今後,すべてのアレルギー疾患,特に難治性喘息や食物アレルギーの根治に有効な新たな治療薬としての開発が期待される.
引用文献References
1) Strachan, D.P. (1989) Hay fever, hygiene, and household size. BMJ, 299, 1259–1260.
2) Jackson, D.J. & Johnston, S.L. (2010) The role of viruses in acute exacerbations of asthma. J. Allergy Clin. Immunol., 125, 1178–1187., quiz, 1188–1179.
3) Kelleher, M.M., Dunn-Galvin, A., Gray, C., Murray, D.M., Kiely, M., Kenny, L., McLean, W.H.I., Irvine, A.D., & Hourihane, J.O. (2016) Skin barrier impairment at birth predicts food allergy at 2 years of age. J. Allergy Clin. Immunol., 137, 1111–1116.e8.
4) Cahenzli, J., Koller, Y., Wyss, M., Geuking, M.B., & McCoy, K.D. (2013) Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe, 14, 559–570.
5) Hill, D.A., Siracusa, M.C., Abt, M.C., Kim, B.S., Kobuley, D., Kubo, M., Kambayashi, T., Larosa, D.F., Renner, E.D., Orange, J.S., et al. (2012) Commensal bacteria-derived signals regulate basophil hematopoiesis and allergic inflammation. Nat. Med., 18, 538–546.
6) Kim, K.S., Hong, S.W., Han, D., Yi, J., Jung, J., Yang, B.G., Lee, J.Y., Lee, M., & Surh, C.D. (2016) Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science, 351, 858–863.
7) Hong, S.W., O, E., Lee, J.Y., Lee, M., Han, D., Ko, H.J., Sprent, J., Surh, C.D., & Kim, K.S. (2019) Food antigens drive spontaneous IgE elevation in the absence of commensal microbiota. Sci. Adv., 5, eaaw1507.
8) Ohnmacht, C., Park, J.H., Cording, S., Wing, J.B., Atarashi, K., Obata, Y., Gaboriau-Routhiau, V., Marques, R., Dulauroy, S., Fedoseeva, M., et al. (2015) Mucosal immunology. The microbiota regulates type 2 immunity through RORgammat(+) T cells. Science, 349, 989–993.
9) Al Nabhani, Z., Dulauroy, S., Marques, R., Cousu, C., Al Bounny, S., Déjardin, F., Sparwasser, T., Bérard, M., Cerf-Bensussan, N., & Eberl, G. (2019) A Weaning Reaction to Microbiota Is Required for Resistance to Immunopathologies in the Adult. Immunity, 50, 1276–1288.e5.
10) Gollwitzer, E.S., Saglani, S., Trompette, A., Yadava, K., Sherburn, R., McCoy, K.D., Nicod, L.P., Lloyd, C.M., & Marsland, B.J. (2014) Lung microbiota promotes tolerance to allergens in neonates via PD-L1. Nat. Med., 20, 642–647.
11) Roan, F., Obata-Ninomiya, K., & Ziegler, S.F. (2019) Epithelial cell-derived cytokines: more than just signaling the alarm. J. Clin. Invest., 129, 1441–1451.
12) McKenzie, A.N. (2014) Type-2 innate lymphoid cells in asthma and allergy. Ann. Am. Thorac. Soc., 11(Suppl 5), S263–S270.
13) Josefowicz, S.Z., Niec, R.E., Kim, H.Y., Treuting, P., Chinen, T., Zheng, Y., Umetsu, D.T., & Rudensky, A.Y. (2012) Extrathymically generated regulatory T cells control mucosal TH2 inflammation. Nature, 482, 395–399.
14) Gould, H.J. & Ramadani, F. (2018) Peanut allergen-specific antibodies go public. Science, 362, 1247–1248.
15) Liang, H.E., Reinhardt, R.L., Bando, J.K., Sullivan, B.M., Ho, I.C., & Locksley, R.M. (2011) Divergent expression patterns of IL-4 and IL-13 define unique functions in allergic immunity. Nat. Immunol., 13, 58–66.
16) Meli, A.P., Fontes, G., Leung Soo, C., & King, I.L. (2017) T Follicular Helper Cell-Derived IL-4 Is Required for IgE Production during Intestinal Helminth Infection. J. Immunol., 199, 244–252.
17) Vijayanand, P., Seumois, G., Simpson, L.J., Abdul-Wajid, S., Baumjohann, D., Panduro, M., Huang, X., Interlandi, J., Djuretic, I.M., Brown, D.R., et al. (2012) Interleukin-4 production by follicular helper T cells requires the conserved Il4 enhancer hypersensitivity site V. Immunity, 36, 175–187.
18) Yusuf, I., Kageyama, R., Monticelli, L., Johnston, R.J., Ditoro, D., Hansen, K., Barnett, B., & Crotty, S. (2010) Germinal center T follicular helper cell IL-4 production is dependent on signaling lymphocytic activation molecule receptor (CD150). J. Immunol., 185, 190–202.
19) Dolence, J.J., Kobayashi, T., Iijima, K., Krempski, J., Drake, L.Y., Dent, A.L., & Kita, H. (2018) Airway exposure initiates peanut allergy by involving the IL-1 pathway and T follicular helper cells in mice. J. Allergy Clin. Immunol., 142, 1144–1158.e8.
20) Kobayashi, T., Iijima, K., Dent, A.L., & Kita, H. (2017) Follicular helper T cells mediate IgE antibody response to airborne allergens. J. Allergy Clin. Immunol., 139, 300–313.e7.
21) Reinhardt, R.L., Liang, H.E., & Locksley, R.M. (2009) Cytokine-secreting follicular T cells shape the antibody repertoire. Nat. Immunol., 10, 385–393.
22) Gowthaman, U., Chen, J.S., Zhang, B., Flynn, W.F., Lu, Y., Song, W., Joseph, J., Gertie, J.A., Xu, L., Collet, M.A., et al. (2019) Identification of a T follicular helper cell subset that drives anaphylactic IgE. Science, 356, eaaw6433.
23) Clement, R.L., Daccache, J., Mohammed, M.T., Diallo, A., Blazar, B.R., Kuchroo, V.K., Lovitch, S.B., Sharpe, A.H., & Sage, P.T. (2019) Follicular regulatory T cells control humoral and allergic immunity by restraining early B cell responses. Nat. Immunol., 20, 1360–1371.
24) Ando, T., Kashiwakura, J.I., Itoh-Nagato, N., Yamashita, H., Baba, M., Kawakami, Y., Tsai, S.H., Inagaki, N., Takeda, K., Iwata, T., et al. (2017) Histamine-releasing factor enhances food allergy. J. Clin. Invest., 127, 4541–4553.
25) Turner, H. & Kinet, J.P. (1999) Signalling through the high-affinity IgE receptor Fc epsilonRI. Nature, 402(Suppl), B24–B30.
26) Gilfillan, A.M. & Rivera, J. (2009) The tyrosine kinase network regulating mast cell activation. Immunol. Rev., 228, 149–169.
27) Kunder, C.A., St John, A.L., Li, G., Leong, K.W., Berwin, B., Staats, H.F., & Abraham, S.N. (2009) Mast cell-derived particles deliver peripheral signals to remote lymph nodes. J. Exp. Med., 206, 2455–2467.
28) Asai, K., Kitaura, J., Kawakami, Y., Yamagata, N., Tsai, M., Carbone, D.P., Liu, F.T., Galli, S.J., & Kawakami, T. (2001) Regulation of mast cell survival by IgE. Immunity, 14, 791–800.
29) Kalesnikoff, J., Huber, M., Lam, V., Damen, J.E., Zhang, J., Siraganian, R.P., & Krystal, G. (2001) Monomeric IgE stimulates signaling pathways in mast cells that lead to cytokine production and cell survival. Immunity, 14, 801–811.
30) Kitaura, J., Song, J., Tsai, M., Asai, K., Maeda-Yamamoto, M., Mocsai, A., Kawakami, Y., Liu, F.T., Lowell, C.A., Barisas, B.G., et al. (2003) Evidence that IgE molecules mediate a spectrum of effects on mast cell survival and activation via aggregation of the FcepsilonRI. Proc. Natl. Acad. Sci. USA, 100, 12911–12916.
31) Kawakami, T. & Kitaura, J. (2005) Mast Cell Survival and Activation by IgE in the Absence of Antigen: A Consideration of the Biologic Mechanisms and Relevance. J. Immunol., 175, 4167–4173.
32) Thueson, D.O., Speck, L.S., Lett-Brown, M.A., & Grant, J.A. (1979) Histamine-releasing activity (HRA). I. Production by mitogen- or antigen-stimulated human mononuclear cells. J. Immunol., 123, 626–632.
33) MacDonald, S.M., Rafnar, T., Langdon, J., & Lichtenstein, L.M. (1995) Molecular identification of an IgE-dependent histamine-releasing factor. Science, 269, 688–690.
34) MacDonald, S.M., Lichtenstein, L.M., Proud, D., Plaut, M., Naclerio, R.M., MacGlashan, D.W., & Kagey-Sobotka, A. (1987) Studies of IgE-dependent histamine releasing factors: heterogeneity of IgE. J. Immunol., 139, 506–512.
35) MacDonald, S.M.(1993) Histamine releasing factors and IgE heterogeneity, 4th Ed., pp.1–11, Mosby-Year Book Incorporated.
36) Bommer, U.A. (2012) Cellular function and regulation of the translationally controlled tumor protein TCTP. Open Allergy J., 5, 19–32.
37) Bommer, U.A. & Thiele, B.J. (2004) The translationally controlled tumour protein (TCTP). Int. J. Biochem. Cell Biol., 36, 379–385.
38) Chen, S.H., Wu, P.S., Chou, C.H., Yan, Y.T., Liu, H., Weng, S.Y., & Yang-Yen, H.F. (2007) A knockout mouse approach reveals that TCTP functions as an essential factor for cell proliferation and survival in a tissue- or cell type-specific manner. Mol. Biol. Cell, 18, 2525–2532.
39) Koide, Y., Kiyota, T., Tonganunt, M., Pinkaew, D., Liu, Z., Kato, Y., Hutadilok-Towatana, N., Phongdara, A., & Fujise, K. (2009) Embryonic lethality of fortilin-null mutant mice by BMP-pathway overactivation. Biochim. Biophys. Acta, 1790, 326–338.
40) Susini, L., Besse, S., Duflaut, D., Lespagnol, A., Beekman, C., Fiucci, G., Atkinson, A.R., Busso, D., Poussin, P., Marine, J.C., et al. (2008) TCTP protects from apoptotic cell death by antagonizing bax function. Cell Death Differ., 15, 1211–1220.
41) Kashiwakura, J., Ando, T., Matsumoto, K., Kimura, M., Kitaura, J., Matho, M.H., Zajonc, D.M., Ozeki, T., Ra, C., MacDonald, S.M., et al. (2012) Histamine-releasing factor has a proinflammatory role in mouse models of asthma and allergy. J. Clin. Invest., 122, 218–228.
42) Wantke, F., MacGlashan, D.W., Langdon, J.M., & MacDonald, S.M. (1999) The human recombinant histamine releasing factor: functional evidence that it does not bind to the IgE molecule. J. Allergy Clin. Immunol., 103, 642–648.
43) Dore, K.A., Kashiwakura, J.I., McDonnell, J.M., Gould, H.J., Kawakami, T., Sutton, B.J., & Davies, A.M. (2018) Crystal structures of murine and human Histamine-Releasing Factor (HRF/TCTP) and a model for HRF dimerisation in mast cell activation. Mol. Immunol., 93, 216–222.
44) Chen, C.Y., Lee, J.B., Liu, B., Ohta, S., Wang, P.Y., Kartashov, A.V., Mugge, L., Abonia, J.P., Barski, A., Izuhara, K., et al. (2015) Induction of Interleukin-9-Producing Mucosal Mast Cells Promotes Susceptibility to IgE-Mediated Experimental Food Allergy. Immunity, 43, 788–802.
45) Brandt, E.B., Strait, R.T., Hershko, D., Wang, Q., Muntel, E.E., Scribner, T.A., Zimmermann, N., Finkelman, F.D., & Rothenberg, M.E. (2003) Mast cells are required for experimental oral allergen-induced diarrhea. J. Clin. Invest., 112, 1666–1677.
46) Kawakami, Y., Sielski, R., & Kawakami, T. (2018) Mouse Body Temperature Measurement Using Infrared Thermometer During Passive Systemic Anaphylaxis and Food Allergy Evaluation. J. Vis. Exp., e58391.
47) Itoh-Nagato, N., Inoue, Y., Nagao, M., Fujisawa, T., Shimojo, N., Iwata, T., Adachi, Y., Arakawa, K., Arima, T., Fukushima, K., et al.; J-OIT group. (2017) Desensitization to a whole egg by rush oral immunotherapy improves the quality of life of guardians: A multicenter, randomized, parallel-group, delayed-start design study. Allergol. Int., 67, 209–216.
48) Kashiwakura, J.I., Ando, T., Karasuyama, H., Kubo, M., Matsumoto, K., Matsuda, T., & Kawakami, T. (2019) The basophil-IL-4-mast cell axis is required for food allergy. Allergy, 74, 1992–1996.