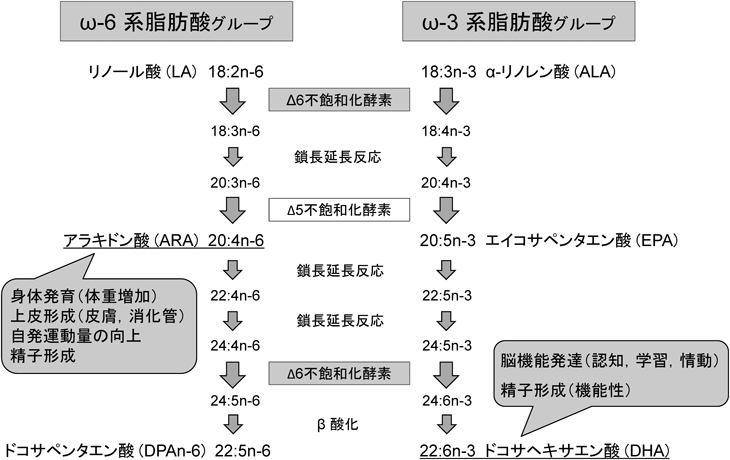
脂質は,エネルギー源としての利用の他,身体の細胞膜の重要な構成成分であり,摂取する脂質(脂肪酸)によって,その細胞,組織・器官の機能に影響し,生活習慣病の予防・罹患にも関わってくる.その中でも多価不飽和脂肪酸のω6とω3系脂肪酸は,膜の流動性の維持に寄与し,膜リン脂質の生理活性成分であり,主要な細胞シグナル伝達エイコサノイドとドコサノイドの前駆体として機能する.しかし,この多価不飽和脂肪酸は飽和脂肪酸や一価不飽和脂肪酸とは異なり,生体内で合成できないため,食品から直接摂取する必要がある必須脂肪酸である.また,炭素数20および22の多価不飽和脂肪酸,特にアラキドン酸(arachidonic acid:ARA; 20:4n-6),エイコサペンタエン酸(eicosapentaenoic acid:EPA; 20:5n-3),およびドコサヘキサエン酸(docosahexaenoic acid:DHA; 22:6n-3)は,動物性脂肪から直接摂取するか,炭素数18のリノール酸(linoleic acid:LA; 18:2n-6)とα-リノレン酸(α-linolenic acid:ALA; 18:3n-3)からΔ6不飽和化酵素(Δ-6-desaturase:D6D)と鎖長延長反応,Δ5不飽和化酵素(Δ-5-desaturase:D5D)の一連の反応によって生体内で合成される(図1)1).このように,生体内では,ω6系脂肪酸とω3系脂肪酸は同じ不飽和化酵素や反応によって代謝され,互いに競合することから,食事中の両脂肪酸のバランスを考慮する必要がある2).また,ω6系脂肪酸は必須脂肪酸であっても,多くの食材に含まれているため通常の食生活であれば欠乏する心配はないが,ω3系脂肪酸は限られた植物油や海洋性動植物にしか含まれておらず,意識して摂取しないと欠乏状態に陥りやすい.実際,循環器系疾患や炎症・アレルギー,脳機能への影響などω3系脂肪酸の生体への有用性は数多く報告されている.近年,メタボローム解析が進歩し,ARAやEPAから合成されるプロスタグランジンやロイコトリエンなどのエイコサノイド,DHAから産出されるレゾルビンやプロテクチンといったドコサノイドの研究も盛んになり,ω3やω6系脂肪酸の個々の脂肪酸の詳細な作用や役割について検討することが重要となってきた3–5).
ω3系脂肪酸の有用性を検討するには,ω3系脂肪酸欠乏(ω3欠乏)動物を作製する必要がある.一般に,実験動物用飼料は,適切量のDHAを含むω3系脂肪酸を含有している.また,通常飼料からω3欠乏飼料に切り換え飼育すると,血漿中DHA量はわずか2週間で半減するが,脳内DHA量の低下は10週間摂取しても正常マウスの約70%程度とほとんど低下しないため,神経系組織での評価は困難である6).また,通常飼育の動物は出生前から離乳までの間に胎盤や母獣乳から十分なDHAを摂取しているので,第1世代で目的とするω3欠乏動物を作製することは容易ではない.これらのことを考慮して,我々は,AIN-93精製飼料を基礎飼料としたα-リノレン酸をほとんど含まないリノール酸豊富なω3欠乏飼料で継代飼育して得た2世代目以降のω3欠乏動物を用いている.ただ,この場合の食事性ω3系脂肪酸欠乏モデルは,α-リノレン酸だけでなくEPAやDHAなど,一連のω3系脂肪酸がすべて低下してしまうため,個々の脂肪酸の役割は評価できない.
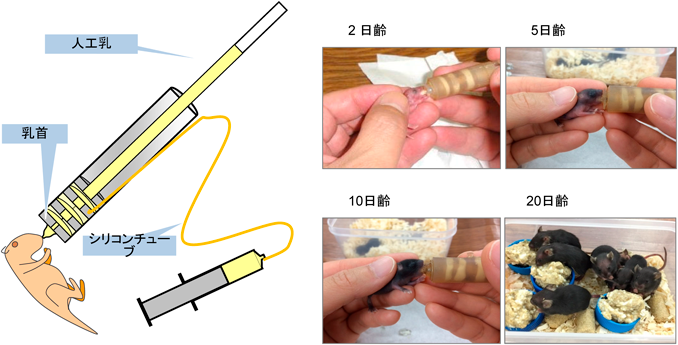
もう一方のω6系脂肪酸の評価においては,あらゆる原料にリノール酸が含まれるため,食事性ω6系脂肪酸欠乏モデルを作製すること自体が困難であった.今回紹介するD6D欠損(D6D-KO)マウスは,脂肪酸代謝の上流と下流の不飽和化が進まないため,飼料に添加した多価不飽和脂肪酸単独の作用,または組合わせの作用の検討を可能にした.さらに,人工哺育法は,生後48時間以内に新生仔を母獣から離し各脂肪酸を調整した人工乳を与えるので,母獣乳の影響を避けた早期介入も可能となり,実験期間が大幅に短縮できるようになった(図2)7).
D6D-KOマウスは,リノール酸やα-リノレン酸を摂取してもARA, EPA, DHAなどの多価不飽和脂肪酸を産生することができない.中村らは,離乳後,リノール酸,α-リノレン酸のみの飼料で飼育し,2か月齢ころから体重の減少,4か月齢ごろには比較的短期間に細胞が代謝回転する皮膚で,刺激により炎症が亢進され引っかき行動による潰瘍性皮膚炎,消化管では主に回腸や十二指腸に局所的潰瘍を観察している.また,通常,ARAやDHAが高濃度に供給される生殖器では,発達不全による不妊も確認している8, 9).
4. 多価不飽和脂肪酸のそれぞれの働きについて10–12)
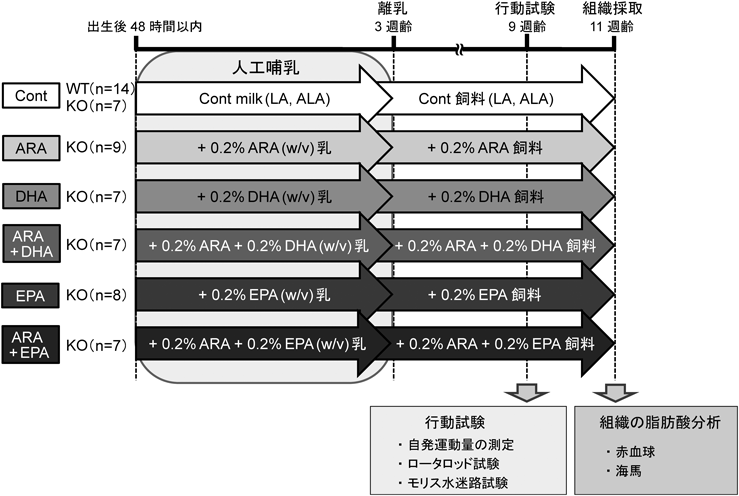
人工哺育法とD6D-KOマウスを組み合わせれば,脳機能を含む身体の著しい成長期である授乳期間から各脂肪酸の供給を調節できる.この実験系を用いて,個々の脂肪酸の役割と相互作用の評価を行った.人工乳は,タンパク質,脂質,炭水化物,ミネラル,ビタミン量は同一とし,多価不飽和脂肪酸として,リノール酸とα-リノレン酸のみを添加したControl乳(Cont乳)と,それにARA, EPA, DHA, ARA+EPA, ARA+DHAの各脂肪酸をそれぞれ0.2%(w/v)ずつ添加した6種類の人工乳を調製した.対照群としては,野生型(WT)マウスにCont乳を与えた.生後15日目からは,人工乳の授乳に加え,固形化した各人工乳を離乳食として与え,3週齢時からは,同様の脂肪酸組成の精製飼料に切り換え通常飼育して,9週齢時から脳機能を評価した後に各組織の脂肪酸組成を測定した(図3).
1)身体形成における多価不飽和脂肪酸の働き
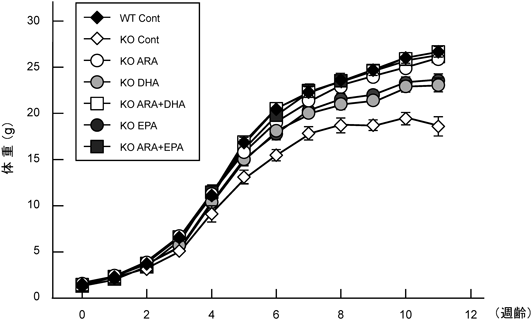
KO Cont群は,WT Cont群と比較して,離乳の3週齢以降から体重増加が顕著に抑制されている.一方,ARAを添加したKO ARA群やKO ARA+EPA群,KO ARA+DHA群はWT Cont群と類似の体重推移を示した.ARAによる身体形成の促進作用は,再現性が複数回確認されている.また,KO DHA群とKO EPA群ではKO Cont群よりも体重増加は示すものの,ARAを含んだ群には及ばず,6週齢以降,徐々に増加量は低下した(図4).これらのことから,エネルギー量はもちろんのこと,タンパク質,脂質,炭水化物量(PFCバランス)やミネラル,ビタミンを等量に調整しても,多価不飽和脂肪酸がリノール酸,α-リノレン酸のみでは,身体成長には不十分であることが確認された.特記すべきことは成長過程における外的所見である.KO Cont群およびKO DHA群では,週齢を重ねるに従って貧血に起因した四肢,耳介の白色と血便,直腸温度の低値が観察されたのに対して,KO EPA群とARAを含む群ではこのような症状はまったく確認されなかった.これはARAから産生されるプロスタグランジンなどのエイコサノイドの,骨格筋の増大や消化管粘膜の増殖・保護作用によるものと考えられた.また,KO DHA群と類似した体重推移を示したKO EPA群で血便等が観察されなかったのは,弱いながらもEPAから生成されるエイコサノイドの作用によるものと推察された.このことから,KO ContやKO DHA群で観察された血便等の症状は,ARA欠乏による消化管粘膜の発達不良に起因した粘膜剥離が示唆される.
2)脳機能における多価不飽和脂肪酸の働き
神経系の中枢である脳は,感情,思考,運動,生命の維持など神経活動の司令塔の役割を果たしている.脳機能の各種多価不飽和脂肪酸の摂取による評価としては,記憶学習に関わるモリス水迷路試験と運動協調性に関わるロータロッド試験を用いた.
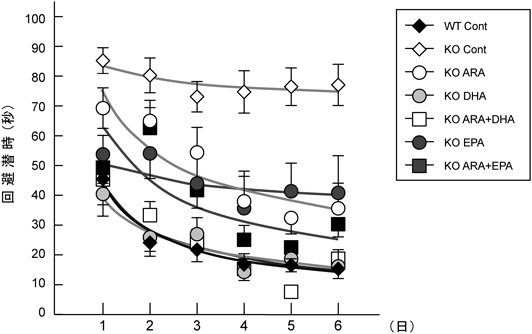
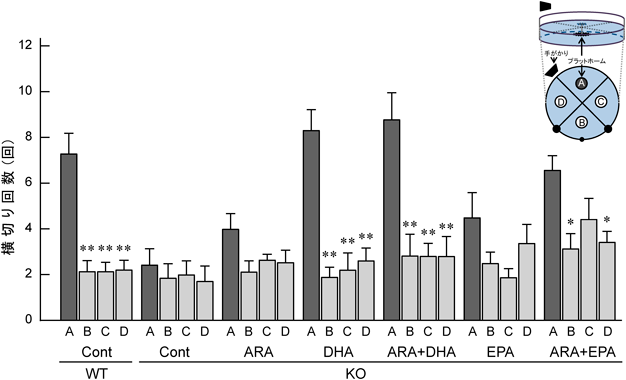
モリス水迷路試験は,プールに沈めたプラットホームを周りの景色を手がかりに探索させるものであり,回避潜時(プラットホームにたどり着く時間)が早ければ,空間認知機能が高いことになる13).WT Cont群は,経日的に回避潜時が短縮し,記憶学習の強化が認められたのに対して,KO Cont群は,試行を繰り返しても回避潜時が短縮することはほとんどなかった.ω3系脂肪酸欠乏動物の認知機能の低下は,可逆的なものであることが多く報告されており,ω3系脂肪酸欠乏動物の課題獲得の遅延は試行を繰り返すことにより,正常群との差が消失する13).しかし,KO Cont群は,学習曲線から課題獲得の遅延ではなく,病的な記憶学習の獲得障害であると考えられる.その他の群の中で,DHAを添加したKO DHAとKO ARA+DHA群はWT Cont群と同等の良好な回避潜時を示したが,ARAやEPAの単独もしくは併用添加したARA+EPA群では程度の差はあるものの,回避潜時の短縮は弱いものであった(図5).6日間の試験試行を終了した翌日に実施した再現試行による確認でも,プラットフォーム位置(ポジションA)を横切る回数は,回避潜時が短縮したKO DHAやKO ARA+DHA群,WT Cont群で他のポジションよりも有意に多く,確実に空間を認識し,プラットフォーム位置を記憶していることが示された.KO ARA+EPA群では,若干の相互作用が観察されるが,それ以外のKO ContやKO ARA, KO EPA群では,ポジションAとB~Dに違いはなく,プラットホーム位置は記憶できておらず,再現試行の90秒間を目標を見つけられずにさまよいながら泳いでいることが示唆された(図6).
運動協調性に用いたロータロッド試験は,回転加速するロッド上を歩行させ,マウスが落下するまでの時間を測定するもので,WT Cont群に対して,KO Cont群とKO ARA群は,著しく落下時間が短く,運動の中枢である小脳の機能低下が推察される.一方,DHAを添加したKO DHAとKO ARA+DHA群では,KO Cont群との間に有意差を得るまでには至っていないが,WT Contレベルまでの運動協調性の改善が観察された.これらの試験に影響を与える自発運動量を検証すると,KO Cont群とKO DHA群はWT Cont群よりも低下する傾向を示し,ARAを含んだARA+DHA, ARA+EPAとEPA群は,WT Cont群と同程度,KO ARA群では,WT Cont群よりも活動量の増加傾向を示していた.一般に,自発運動量が高いと見かけ上の記憶学習能や運動機能が向上したように,また,運動量が低いだけで記憶学習能が低下していると見誤ることがある.しかし,DHA添加群は自発運動量が低いにもかかわらず両試験ともWT Cont群と同様の良好な成績を示したのに対して,ARA群は,自発運動を高めながらも,脳機能の改善が認められなかったことから,脳機能の発達にDHAの摂取はきわめて重要であり,ARAの貢献度は低いことが考えられる.
3)脳組織の多価不飽和脂肪酸組成
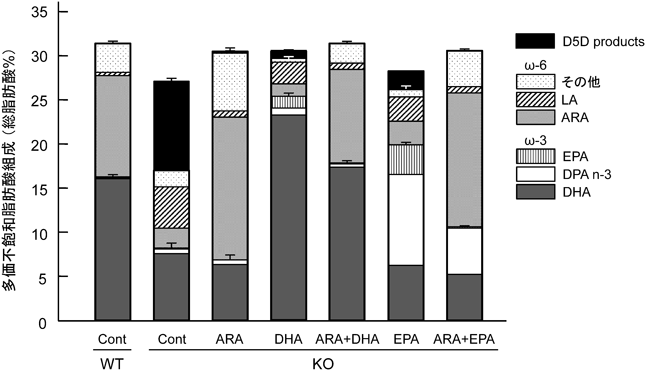
試験終了後,脳の海馬中の脂肪酸,特に多価不飽和脂肪酸の組成を比較すると,KO Cont群では,著しいω6とω3系脂肪酸の低下と,多価不飽和脂肪酸量を維持するためのΔ5不飽和化酵素により代謝された脂肪酸(D5D products)が検出された.また,KO ARA群やKO DHA群の添加した脂肪酸の顕著な上昇はいうまでもないが,KO DHA群の脳内では,ARAがほとんど存在しないため,通常,検出されないEPAがDHAのレトロ変換により蓄積していた.このことから,炭素数20の脂肪酸が脳機能の構造の維持に必要であることが推察される.さらにKO EPA, KO ARA+EPA群では,摂取させたEPAが鎖長延長したドコサペンタエン酸(docosapentaenoic acid:DPAn-3, 22:5n-3)の上昇が確認されている.通常,脳内のω3系脂肪酸はDHAが蓄積されるため,脳内DHAが減少した場合,炭素数22の脂肪酸を維持するために脳機能にはほとんど寄与しないω6系脂肪酸のDPAn-6が代償的に上昇することが知られている13–16).D6D KOマウスは脳内DHAが低下してもARAからDPAn-6を代謝することができず,さらにはEPAからDHAを代謝できないので,EPAを摂取させた際には,EPAを鎖長延長したDHAと同じ炭素数22のDPAn-3を補填していることが考えられた.しかし,KO EPAとKO ARA+EPA群が良好な脳機能の改善作用を示していない今回の行動試験の結果から,DPAn-3には,直接脳機能を改善する作用はないと考えられた16).一方,ω6系脂肪酸でEPAと同じ炭素数20のARAからは,鎖長延長反応によるドコサテトラエン酸(docosatetraenoic acid:DTA, 22:4n-6)の蓄積がほとんど確認されていないことから,脳の炭素数20の多価不飽和脂肪酸は,ω6系脂肪酸のARAが優先され,炭素数22ではω3系脂肪酸のDHAが優先的に蓄積されることが示唆された.これらのことから,脳内脂肪酸組成は,ω6/ω3比だけでなく炭素数20や22の脂肪酸も重要な要因であると考えられる(図7).すべての試験群の中で,KO ARA+DHA群の脂肪酸組成のみが脳機能評価と同様にWT Cont群と酷似している.このことから,成長発達期には,ARAとDHAの摂取が必要不可欠であると考えられる.
今回紹介したD6D-KOマウスの実験では,KO ARA+DHA群の人工乳に添加したARAとDHAは等量としたが,至適な添加量,添加比率についてまったく不明であり検討の余地が残っている.また,これらの脂肪酸を直接摂取すべき至適期間については,明確な情報は得られていない.通常飼料で飼育した母獣で養育された授乳期間中のWT新生仔の脳や肝臓,母獣乳中の脂肪酸組成の変化から,WTマウスであっても,生後2週間程度まではD5DやD6Dの代謝酵素の活性化が未熟であることが推察される(投稿準備中).
これら問題の鍵となるのはヒトの母乳の脂質組成であるが,食生活や人種の違いから国によって報告値は大きく変動する17, 18).ARAとDHAのそれぞれの平均値は総脂肪酸の0.47%と0.32%だが,どちらの脂肪酸にも最低値と最高値に大きな開きがあり,その変動は特にDHAで大きい(ARA; 0.24~1.0%,DHA; 0.06~1.4%).また,母体へのARAやEPA, DHAの直接的な補給によって,母乳中ARAやDHA濃度の増加も報告されていることから,食餌性脂質の母乳への影響は大きい19, 20).これらの報告を受け,欧州小児科学会および児童保健財団の見解が発表され,乳児およびその後の成長期間でARAとDHAの両方を与えることを推奨している21).DHAは食事による母乳中濃度の変動が大きいため,DHAの添加による有効性が見いだされることが多いが,いくつかの研究ではDHAを含む処方にARAを添加することで知能指数(IQ)の上昇などの効果またはその向上がもたらされることが示されている22).そこで,現在,DHA濃度を固定し,ARA濃度を0~2までの比率に振った1:0, 1:1, 1:2に設定し,添加量を前回の1/4[0.05%(w/v)]に減らした人工乳を作製して,D6D-KOマウスと人工哺育法を用い,授乳,発達期のDHAとARAの至適な添加比率や添加量,添加期間を検討中である.これら人工乳で哺育されたマウスの行動はいうまでもなく,脳内脂肪酸組成も興味のあるところである.
これまでω3系脂肪酸の脳機能に対する有用性は数多く報告されているが,今回,D6D-KOマウスとその新生仔の人工飼育法の組み合わせによって,成長発達期において身体成長にはARAが,脳機能の発達・維持にはDHAが重要な役割を果たしていることを証明した.D6Dが正常に機能する個体では前駆体脂肪酸のリノール酸やα-リノレン酸,EPAがARAやDHAに代謝され有効性を示すことが考えられるが,脂質代謝活性の低い乳幼児やある集団では,ARAやDHAの直接的な摂取が不可欠である.
近年,脂肪酸の代謝酵素活性に人種差のあることが報告されている.ヨーロッパ系,東アジア系の不飽和化酵素(FADS)遺伝子のマイナー対立遺伝子を持つ一部の集団は,酵素活性が低く,リノール酸やα-リノレン酸を摂取してもARAおよびDHAが合成されにくいことが考えられるため,周産期の母親や発達期の小児は直接食品から補給する必要がある23).Δ6不飽和化酵素ヘテロ欠損(D6D-HT)マウスは,代謝酵素活性が約半分と酵素活性が低い集団に類似していることから,リノール酸とα-リノレン酸のみを与えて長期間飼育すると,成熟期以降,体重減少や皮膚の炎症により脱落個体が目立ち始め,肝臓や脳(海馬)でのARA蓄積量の低下が老齢期まで持続的に観察されている.また,DHAも老齢期で蓄積量が低下していた24).これらのことは,ARAとDHAの各臓器の需要性の違いと考えられ,成長・成熟期からのARA摂取と,高齢期のDHA補充が重要と考えられる.
多価不飽和脂肪酸は二重結合の位置が少し違うことによって役割は大きく異なり,そのバランスで疾病の引き金になる可能性がある.三大栄養素の中でも,脂質は唯一,種類を選択して摂取できる.「脂質」ひと塊ではなく,摂取する必要がある「油脂」を理解し,食生活に活かせるようこれからも多価不飽和脂肪酸について広く広報していきたい.また,自身の状況を把握するためにも,代謝酵素の判別や血中レベル脂肪酸組成が簡便に測定できる方法が確立されることを望む.
引用文献References
1) Nakamura, M.T. & Nara, T.Y. (2004) Structure, function, and dietary regulation of delta6, delta5, and delta9 desaturases. Annu. Rev. Nutr., 24, 345–376.
2) Mathias, R.A., Sergeant, S., Ruczinski, I., Torgerson, D.G., Hugenschmidt, C.E., Kubala, M., Vaidya, D., Suktitipat, B., Ziegler, J.T., Ivester, P., et al. (2011) The impact of FADS genetic variants on ω6 polyunsaturated fatty acid metabolism in African Americans. BMC Genet., 12, 50.
3) Bannenberg, G.L., Chiang, N., Ariel, A., Arita, M., Tjonahen, E., Gotlinger, K.H., Hong, S., & Serhan, C.N. (2005) Molecular circuits of resolution: Formation and actions of resolvins and protectins. J. Immunol., 174, 4345–4355.
4) Arita, M., Bianchini, F., Aliberti, J., Sher, A., Chiang, N., Hong, S., Yang, R., Petasis, N.A., & Serhan, C.N. (2005) Stereochemical assignment, antiinflammatory properties, and receptor for the omega-3 lipid mediator resolvin E1. J. Exp. Med., 201, 713–722.
5) Arita, M. (2012) Mediator lipidomics in acute inflammation and resolution. J. Biochem., 152, 313–319.
6) Moriguchi, T., Harauma, A., & Salem, N. Jr. (2013) Plasticity of mouse brain docosahexaenoic acid: Modulation by diet and age. Lipids, 48, 343–355.
7) Yasuda, H., Harauma, A., Kato, M., Ootomo, Y., Hatanaka, E., & Moriguchi, T. (2016) Artificially reared mice exhibit anxiety-like behavior in adulthood. Exp. Anim., 65, 267–274.
8) Stroud, C.K., Nara, T.Y., Roqueta-Rivera, M., Radlowski, E.C., Lawrence, P., Zhang, Y., Cho, B.H., Segre, M., Hess, R.A., Brenna, J.T., et al. (2009) Disruption of FADS2 gene in mice impairs male reproduction and causes dermal and intestinal ulceration. J. Lipid Res., 50, 1870–1880.
9) Roqueta-Rivera, M., Abbott, T.L., Sivaguru, M., Hess, R.A., & Nakamura, M.T. (2011) Deficiency in the omega-3 fatty acid pathway results in failure of acrosome biogenesis in mice. Biol. Reprod., 58, 721–732.
10) Hatanaka, E., Harauma, A., Yasuda, H., Watanabe, J., Nakamura, M.T., Salem, N. Jr., & Moriguchi, T. (2016) Essentiality of arachidonic acid intake in murine early development. Prostaglandins Leukot. Essent. Fatty Acids, 108, 51–57.
11) Harauma, A., Yasuda, H., Hatanaka, E., Nakamura, M.T., Salem, N. Jr., & Moriguchi, T. (2017) The essentiality of arachidonic acid in addition to docosahexaenoic acid for brain growth and function. Prostaglandins Leukot. Essent. Fatty Acids, 116, 9–18.
12) Harauma, A., Hatanaka, E., Nakamura, M.T., Salem, N. Jr., & Moriguchi, T. (2017) Effects of arachidonic acid, eicosapentaenoic acid and docosahexaenoic acid on brain development using artificial rearing of delta-6-desaturase knockout mice. Prostaglandins Leukot. Essent. Fatty Acids, 127, 32–39.
13) Moriguchi, T., Greiner, R.S., & Salem, N. Jr. (2000) Behavioral deficits associated with dietary induction of decreased brain docosahexaenoic acid concentration. J. Neurochem., 75, 2563–2573.
14) Moriguchi, T. & Salem, N. Jr. (2003) Recovery of brain docosahexaenoate leads to recovery of spatial task performance. J. Neurochem., 87, 297–309.
15) Greiner, R.S., Catalan, J.N., Moriguchi, T., & Salem, N. Jr. (2003) Docosapentaenoic acid does not completely replace DHA in n-3 FA-deficient rats during early development. Lipids, 38, 431–435.
16) Lim, S.Y., Hoshiba, J., Moriguchi, T., & Salem, N. Jr. (2005) N-3 fatty acid deficiency induced by a modified artificial rearing method leads to poorer performance in spatial learning tasks. Pediatr. Res., 58, 741–748.
17) Yuhas, R., Pramuk, K., & Lien, E.L. (2006) Human milk fatty acid composition from nine countries varies most in DHA. Lipids, 41, 851–858.
18) Brenna, J.T., Varamini, B., Jensen, R.G., Diersen-Schade, D.A., Boettcher, J.A., & Arterburn, L.M. (2007) Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am. J. Clin. Nutr., 85, 1457–1464.
19) Makrides, M., Neumann, M.A., & Gibson, R.A. (1996) Effect of maternal docosahexaenoic acid (DHA) supplementation on breast milk composition. Eur. J. Clin. Nutr., 50, 352–357.
20) Weseler, A.R., Dirix, C.E., Bruins, M.J., & Hornstra, G. (2008) Dietary arachidonic acid dose-dependently increases the arachidonic acid concentration in human milk. J. Nutr., 138, 2190–2197.
21) Koletzko, B., Bergmann, K., Brenna, J.T., Calder, P.C., Campoy, C., Clandinin, M.T., Colombo, J., Daly, M., Decsi, T., Demmelmair, H., et al. (2020) Should formula for infants provide arachidonic acid along with DHA? A position paper of the European Academy of Paediatrics and the Child Health Foundation. Am. J. Clin. Nutr., 111, 10–16.
22) Birch, E.E., Garfield, S., Hoffman, D.R., Uauy, R., & Birch, D.G. (2000) A randomized controlled trial of early dietary supply of long-chain polyunsaturated fatty acids and mental development in term infants. Dev. Med. Child Neurol., 42, 174–181.
23) Kothapalli, K.S., Ye, K., Gadgil, M.S., Carlson, S.E., O’Brien, K.O., Zhang, J.Y., Park, H.G., Ojukwu, K., Zou, J., Hyon, S.S., et al. (2016) Positive selection on a regulatory insertion–deletion polymorphism in FADS2 influences apparent endogenous synthesis of arachidonic acid. Mol. Biol. Evol., 33, 1726–1739.
24) Harauma, A., Sueyasu, T., Tokuda, H., Yasuda, H., Hoshi, Y., Kaneda, Y., Rogi, T., Shibata, H., Nakamura, T.M., & Moriguchi, T. (2020) Changes in behavior and fatty acid composition induced by long-term reduction in murine Δ6-desaturation activity. Prostaglandins Leukot. Essent. Fatty Acids, 155, 102079.