食事性スフィンゴ脂質の機能Functions of dietary sphingolipids
京都大学大学院農学研究科Graduate School of Agriculture, Kyoto University ◇ 〒606–8502 京都府京都市左京区北白川追分町 ◇ Kitashirakawaoiwakecho, Sakyo-ku, Kyoto, Kyoto 606–8502, Japan
発行日:2020年10月25日Published: October 25, 2020
スフィンゴ脂質の化学構造は多様であり,生物種によってその構造や組成は特有である.我々はスフィンゴ脂質を日常的に摂取しているが,他の脂質成分と比べて,食品成分としての情報が少ない.しかしながら,食事性スフィンゴ脂質のもつさまざまな機能への注目が高まってきており,食品成分としての消化吸収機構や体内動態なども明らかになりつつある.特に皮膚バリア機能への効果については,特定保健用食品や機能性表示食品への利用も始まっている.しかしながら,そのメカニズムについては不明な点も多い.本稿では,筆者らの研究成果を含めて,これまでわかってきた食品成分としてのスフィンゴ脂質について解説する.
© 2020 公益社団法人日本生化学会© 2020 The Japanese Biochemical Society
スフィンゴ脂質に初めてふれたのは30年近く前である.当時は分析法もわからず,帯広畜産大学・大西正男先生にTLCを用いた分析法をご指導いただいた1).スフィンゴ脂質という言葉に神秘的な響きを感じ,生涯の研究テーマになるような気がしてから,紆余曲折はあるものの,スフィンゴ脂質を食品成分としてとらえた研究を今日まで進めてきた.スムーズとはいえないが,さまざまなことが少しずつ明らかとなってきている.著名なスフィンゴ脂質の専門家が多くいらっしゃる中で,その食品機能にこだわった研究者として,多少なりともその進展に貢献してきたと自負している.本稿では,これまでの筆者らの研究成果を含めて,食品成分としてのスフィンゴ脂質とその機能性について解説する.
スフィンゴ脂質とは,長鎖アミノアルコールであるスフィンゴイド塩基を骨格成分とする脂質の総称である(図1).ヒトを含む哺乳動物に主要なスフィンゴイド塩基は,炭素数18のスフィンゴシン(2-アミノ4-オクタデセン-1.3-ジオール,d18:1)であり,飽和型スフィンガニン(ジヒドロスフィンゴシン,d18:0)とトリヒドロキシ型フィトスフィンゴシン(t18:0)も存在する.しかしながら,脂肪酸の分子種のように,炭素数や二重結合の数が異なるスフィンゴイド塩基の分子種も自然界には存在し,生物種によってその組成や構造に特徴がある.高等植物では,8位に不飽和結合をもつものが主要であり,ジヒドロキシ型の8-モノエン(8-スフィンゲニン)や4,8-ジエン(4,8-スフィンガジエニン),トリヒドロキシ型の8-モノエン(4-ヒドロキシ-8-スフィンゲニン)といった特有のスフィンゴイド塩基が存在する2).4位の不飽和結合はtrans異性体のみであるが,8位の不飽和結合にはcisとtransのどちらの異性体も存在する.キノコや酵母などの真菌類の場合,9-メチル型(9-メチル-4,8-スフィンガジエニン)を有することや3),海産無脊椎動物では17や19といった奇数の炭素鎖をもつものも多く,さらにはトリエン型も確認されている4, 5).
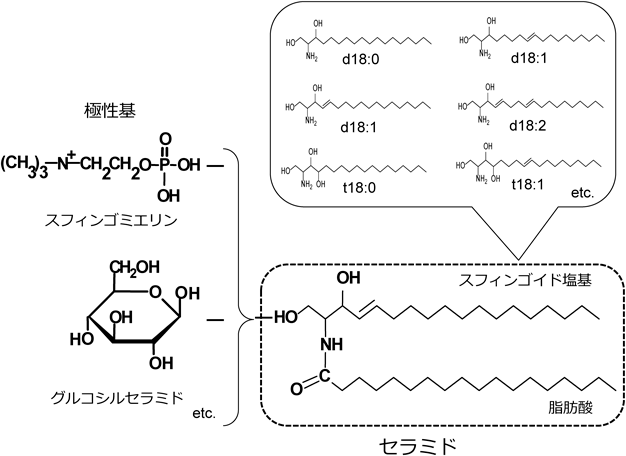
セラミドは,スフィンゴイド塩基と脂肪酸がアミド結合したものであるが,自然界での存在量はわずかであり,一般的には,その1位の水酸基に極性基が結合した複合スフィンゴ脂質として存在する.リン酸コリンが結合したスフィンゴミエリンなどのスフィンゴリン脂質と,極性基が糖からなるスフィンゴ糖脂質(グルコシルセラミド,ガラクトシルセラミド,ガングリオシドなど)に分類される.ガングリオシドはシアル酸(N-アセチルノイラミン酸)を含む構成糖をもつスフィンゴ糖脂質の総称であり,複雑な糖鎖を有する.また,厳密な意味ではリン脂質ではないが,海産軟体動物などには極性基の構造に炭素とリン原子が共有結合しているCP化合物をもつスフィンゴホスホノ脂質も相当量含まれており,セラミドアミノエチルホスホン酸(ceramide aminoethyl phosphonate:CAEP)が主要なものとして知られている6).
スフィンゴ脂質は細胞膜の構成成分であり,さまざまな食品素材から日常的に摂取されている.動物性食品では,牛乳,卵,畜肉などの畜産物に含まれるスフィンゴミエリンが主要であるが,グルコシルセラミドやガラクトシルセラミドなどのスフィンゴ糖脂質も含まれる.一方,植物性食品では,穀類や豆類などのグルコシルセラミドが主要なものとなる.たとえば,牛乳100 mL中には4~12 mg程度のスフィンゴミエリンと0.6~1.1 mgのグリコシルセラミド(グルコシルセラミドとガラクトシルセラミドを含む),0.7~1.5 mgのラクトシルセラミド,さらには数百µg~1 mgのガングリオシドが含まれている.畜肉では25~40 mg/100 g,卵では80~170 mg/100 g程度のスフィンゴミエリンがそれぞれ含まれている7).植物のスフィンゴ脂質含有量は,気候や栽培地域,品種による影響も受けるが,一般的には穀類や豆類のグルコシルセラミド含有量が他の植物に比べて多く,10~40 mg/100 gである7, 8).小麦は消費量も多いことから,植物からのスフィンゴ脂質供給源として主要なものとなる.しかしながら,植物は全般的に脂質含量が低いため,そのスフィンゴ脂質含有量は1~10 mg/100 g程度である8).
アメリカ人の1日あたりの全スフィンゴ脂質摂取量は300~400 mgと算出されており,食品全体の摂取量に対して0.01~0.02%程度を占める7).一方,日本人の1日あたりの摂取量は,食事を直接分析した値からスフィンゴミエリン50~110 mg,グルコシルセラミド60~80 mg程度と示されている9).筆者らが以前調べたさまざまな植物における糖脂質含有量を用いて,それぞれの摂取量から換算すると,植物由来グルコシルセラミドの摂取量は1日あたり約50 mgと見積もられる8).日本人の場合には水産物からの寄与も予想されるため,筆者らの分析値を用いて,年間の水産物消費量から換算したところ,1日あたりグルコシルセラミド1 mg,スフィンゴミエリン4 mg, CAEP 4 mg程度となった(未発表).ガングリオシドについては,カナダ人の場合で,シアル酸量として1日あたり平均100 µg以下(概算でガングリオシド340 µg程度)と報告されている10).
経口摂取されたグルコシルセラミドやスフィンゴミエリンなどの複合スフィンゴ脂質は,極性基(リン酸コリンや糖),脂肪酸,スフィンゴイド塩基といった構成単位に分解された後,それぞれが小腸上皮細胞から体内へと吸収されることが古くから示されている(図2).しかしながら,一般的に食品に含まれているトリアシルグリセロールやホスファチジルコリンのようなグリセロ脂質と比較すると,スフィンゴ脂質の消化率はきわめて低い.
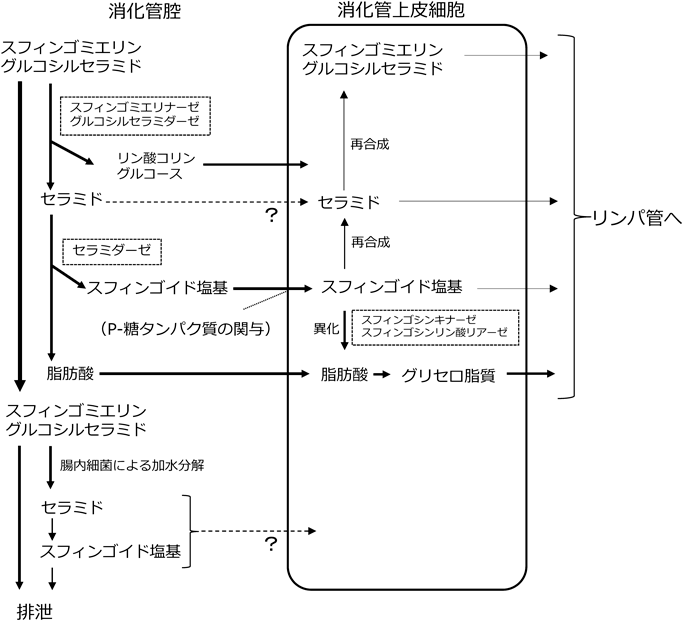
最も一般的なスフィンゴリン脂質であるスフィンゴミエリンは,小腸上皮に発現しているスフィンゴミエリナーゼによって,セラミドとリン酸コリンに加水分解される11, 12).スフィンゴミエリナーゼは,至適pHの違いから酸性,中性,アルカリ性の三つに分類されるが,小腸上皮に存在する酵素は,アルカリ性スフィンゴミエリナーゼであり,至適pH 8.5~9.5である.空腸部における上皮細胞の微絨毛部において強く発現しているが,一次胆汁酸(タウロコール酸やタウロケノデオキシコール酸など)によって活性化することや,C末端側がトリプシンによって加水分解され,活性型として消化管腔内に遊離することが示されており,スフィンゴミエリンは消化管腔内で消化されるものと考えられる13).筆者らは海産無脊椎動物に特有のスフィンゴホスホノ脂質であるCAEPも,スフィンゴミエリンと同様に極性基部分が分解され,消化管内でセラミドが生じることを動物実験で確認している14).スフィンゴミエリンの分解と比較して,中性ではより速やかに分解されることから,アルカリ性スフィンゴミエリナーゼ以外の消化酵素の関与にも興味がもたれるが,詳細は不明である.
スフィンゴ糖脂質であるグルコシルセラミドも,スフィンゴミエリンと同様に消化管内でセラミドとグルコースに分解される.小腸粘膜のグルコシルセラミダーゼ活性には,小腸上皮細胞の微絨毛膜上に存在する二糖類加水分解酵素複合体の一つであるラクターゼ・フロリジン加水分解酵素のフロリジン加水分解活性部位が関わる15, 16).哺乳動物とは異なった特有のスフィンゴイド塩基構造を有する植物由来や海産無脊椎動物由来のグルコシルセラミドについても,動物由来のものとほぼ同様に消化されることを筆者らは動物実験で確認している17, 18).
複合スフィンゴ脂質の消化で生じた,あるいは経口的に摂取されたセラミドについては,中性セラミダーゼによってスフィンゴイド塩基と遊離脂肪酸に分解される.中性セラミダーゼの至適pHは7付近であり,十二指腸,空腸,回腸といった小腸の広範囲で発現している.この中性セラミダーゼは,Ash2遺伝子であることが遺伝子欠損モデルマウスの解析から同定されている19).胆汁酸の共存による活性の増強や,小腸粘膜上皮細胞で発現していることや生理的な胆汁酸濃度で粘膜から抽出できること,トリプシンやキモトリプシンなどのタンパク質分解酵素に対して耐性を示すことなどから,消化管腔内でのセラミドの消化にも中性セラミダーゼが関与することが示されている20, 21).
スフィンゴ脂質は,消化管内でリン酸コリンや糖,脂肪酸,スフィンゴイド塩基へと分解されて,それぞれが小腸上皮細胞から吸収される.小腸上皮細胞に取り込まれた後,動物細胞の主要なスフィンゴイド塩基であるスフィンゴシンの一部は,スフィンゴ脂質(スフィンゴミエリン,グリコシルセラミド,セラミドなど)に再利用されるが,大部分は脂肪酸に代謝され,グリセロ脂質として体内に吸収されることが古くから知られている22, 23).スフィンゴシンの細胞内における異化経路の最初の反応は,スフィンゴシンキナーゼによるリン酸化であり,スフィンゴシン1-リン酸が生じる.さらに,スフィンゴシンリン酸リアーゼの作用によって,リン酸エタノールアミンとアルデヒド(ヘキサデセナール)に分解される24)(図2).このとき生じたヘキサデセナールは,パルミチン酸へと酸化された後,トリアシルグリセロールやグリセロリン脂質へと組み込まれ,カイロミクロンに取り込まれて体内へと運ばれる.実際に他の臓器と比べて,小腸ではスフィンゴシン異化に関わる酵素の発現と活性が高いことが確認されている25, 26).ヘキサデセナールからパルミチン酸への詳細な経路については,ヘキサデセナールがヘキサデセン酸へと酸化され,次いでヘキサデセノイルCoAとなった後に,飽和化されてパルミトイルCoAを生じることが明らかにされている27).また,水酸基を三つ有するフィトスフィンゴシンの場合は,同様の反応を受けるものの,2-ヒドロキシパルミチン酸が生じ,α酸化によってペンタデカン酸(C15:0)に変換される28).同位体ラベル化スフィンゴイド塩基を用いたラットの検討によって,スフィンゴシンの50~70%,ジヒドロスフィンゴシン(スフィンガニン)の約90%は,吸収された後,リンパ液中のトリアシルグリセロール画分に取り込まれることが示されている22).
スフィンゴ脂質の消化管吸収率は,グリセロ脂質などの他の脂質と比べて低い.ラットのリンパ管カニュレーション試験では,スフィンゴミエリンの場合,経口投与24時間のリンパへの回収率は,脂肪酸部分で最大60%程度,スフィンゴシン部分では10%に満たないことが示されている22).グルコシルセラミドはさらに吸収されにくく,脂肪酸部分で20~40%程度,スフィンゴシン部分では3~4%である23).消化管内のスフィンゴ脂質分解活性が低くく,消化されにくいことが理由の一つと推測される.マウスの場合,ラベル化したスフィンゴミエリンの経口投与によって,90分後には投与量の12%相当が大腸にまで到達する29).ラットの場合でも,経口投与したスフィンゴミエリンの33~45%が糞中に排泄され,そのうちの50%以上が未分解のスフィンゴミエリンであり,加えてセラミドやスフィンゴシンも検出されている30).ヒト腸内細菌やラット盲腸内容物にもグルコシルセラミダーゼ活性が認められることから,下部消化管に達した未分解のスフィンゴ脂質の一部は,腸内細菌によって分解されるものと推測される17, 31).
植物に特有のスフィンゴイド塩基は,スフィンゴシンと比べると小腸上皮細胞から吸収されにくいようである.筆者らの行ったラットリンパカニュレーション法による直接的な吸収の評価では,植物に特有の4,8-スフィンガジエニンのリンパへの回収率は,投与量に対して約0.2%程度であり,スフィンゴシンと比べて半分以下である32).また,海産物由来のd17:1やd19:2といった特有のスフィンゴイド塩基についても,ほぼ同程度吸収されることが確認されている18).小腸上皮細胞モデルとしてよく用いられるCaco-2細胞を用いた検討から,スフィンゴシン以外のスフィンゴイド塩基は,薬物排出トランスポーターであるP-糖タンパク質によって,細胞外へ排出されることが示されており33),P-糖タンパク質阻害剤の共存により,植物に特有の4,8-スフィンガジエニンの吸収が高まることがin vivo試験でも確認されている34).P-糖タンパク質の基質特異性は低く,不飽和結合の位置や数などのスフィンゴイド塩基の微細な構造の違いを認識できるとは考えにくいことから,他に重要な選択的吸収機構が存在する可能性も推測され,その詳細な機構について検討を進めている.
一方で,スフィンゴミエリン投与後のラットリンパ管から検出されるセラミド分子組成が,投与したスフィンゴミエリンのセラミド部分の組成に類似しており35),小腸からセラミド構造を維持したまま吸収される可能性が考えられる.筆者らの最近の検討から,醤油粕から調製したセラミドを経口投与したマウスの血中から,投与した分子組成に酷似したセラミド分子種が検出されており,一部のセラミドは,分解を受けずに直接吸収されるものと推測される(未発表).現在,13C-グルコースを炭素源とした菌体から調製した安定同位体ラベル化スフィンゴ脂質(グルコシルセラミドとセラミド)を用いて解析を進めているが,これらの一部は分解されずに吸収される可能性が高い.
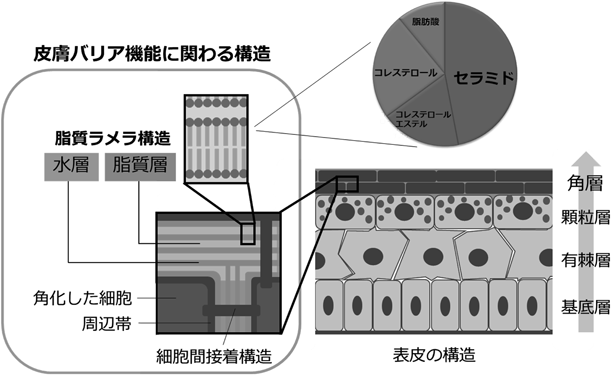
皮膚は外側から,表皮,真皮,皮下組織に大別される.皮膚バリア機能を担う表皮層は,主に表皮角化細胞(ケラチノサイト)からなっており,内側から基底層,有棘層,顆粒層へと分化しながら外側に移動し,最終的には脱核して死んだ細胞から成る角層(角質層)を形成する(終末角化)(図3).ケラチノサイトは,分化の後期(顆粒層)に大量かつ多様な分子種のセラミドを産生するが,終末角化の際にこれらのセラミドを含む脂質を細胞外に放出する(角質細胞間脂質).その約50%を占めるセラミドがラメラ構造の形成に寄与し,皮膚バリア機能を担う36).ヒトの表皮には16種類のセラミドが含まれており,4種類のスフィンゴイド塩基[スフィンゴシン(S),ジヒドロスフィンゴシン(DS),フィトスフィンゴシン(P),6-ヒドロキシスフィンゴシン(H)]と4種類の脂肪酸[ノルマル脂肪酸(N),α-ヒドロキシ脂肪酸(A),ω-ヒドロキシ脂肪酸(O),ω-ヒドロキシ脂肪酸にリノール酸がエステル結合したもの(EO)]の組合わせから分類される36, 37).加齢やアトピー性皮膚炎などによって表皮セラミドが減少することがバリア機能の低下に関係することが示されている38, 39).なかでもω-ヒドロキシ脂肪酸を含むセラミドは,哺乳動物の表皮に特有の成分であり,バリア機能に重要といわれている40, 41).
角層に多く含まれているセラミドが皮膚バリア機能に深く関わっていることから,スフィンゴ脂質の経口摂取による皮膚機能への影響が注目されてきた.実際に動物実験やヒト試験から,スフィンゴ脂質の経口摂取が皮膚バリア機能の向上に寄与することが数多く報告されている.筆者らの検討を含めて,ドライスキン様モデルマウス(HR-ADマウス)を用いた検討によって,植物由来グルコシルセラミド(こんにゃく,トウモロコシ,パイナップル,ビートなど)42–45)や牛乳由来スフィンゴミエリン46, 47),海産物由来スフィンゴ脂質18)の摂取による皮膚バリア改善作用が示されている.この場合のほとんどが,0.1%程度の飼料への添加で評価されている.一方,ヒト試験でも植物由来グルコシルセラミドの継続摂取により,経皮水分蒸散量の抑制と角層水分量の増加が確認されているが48, 49),前述したような日常的なスフィンゴ脂質の摂取量に対して,極端に少ない投与量(0.6~1.8 mg/日)で効果が認められており,そのメカニズムの解明に強く興味がもたれる.
経口摂取されたスフィンゴ脂質の消化管吸収率はきわめて低いため,皮膚に到達して再利用される可能性は低いことが容易に予想される.また,摂取されたスフィンゴ脂質と表皮に含まれるセラミドの化学構造は厳密には異なるにも関わらず,皮膚に対して効果を発揮する.これらの矛盾点を明確に説明できるデータはいまのところ示されていない.筆者らの動物実験から,スフィンゴ脂質摂取による表皮のセラミド合成酵素の発現上昇が確認されており,内因性スフィンゴ脂質の合成促進が皮膚バリア機能向上作用の機構の一つと推測される43).さらに牛乳由来スフィンゴミエリン摂取によって,表皮ω-ヒドロキシセラミド量が増加することも見いだされており47),植物由来グルコシルセラミドやイカ由来CAEPの摂取でも,表皮ω-ヒドロキシセラミド量の増加とその合成に関わる酵素の発現上昇が確認されている(投稿中).スフィンゴ脂質の皮膚への効果については,摂取させたスフィンゴ脂質の化学構造にほぼ依存しないことから,体内で生じる共通の代謝物が効果を発揮する可能性も推定される.その詳細なメカニズムについて,科学的な根拠を見いだすことが今後の大きな課題ともいえる.
スフィンゴ脂質の食品機能性として,Merrillらの研究グループからスフィンゴミエリンやグルコシルセラミドの経口摂取による大腸がん(または大腸腺腫)抑制作用が動物実験で報告されている50, 51).その作用機序の一部として,下部消化管に到達したスフィンゴ脂質が腸内細菌によって加水分解され,生じたセラミドやスフィンゴイド塩基ががん細胞にアポトーシスを誘導することが推測される.植物や海産物由来のスフィンゴイド塩基でもがん細胞に対して,強いアポトーシス誘導作用を示すことが確認されている5, 52, 53).さらに動物実験や培養細胞を用いた検討から,スフィンゴミエリンやグルコシルセラミドは下部消化管での炎症反応を抑制することも見いだされており,LPSやTNFαによって誘発される炎症反応において,それぞれの受容体に対してスフィンゴ脂質が直接作用する可能性も推測されている54, 55).
スフィンゴ脂質は消化や吸収を受けにくく,下部消化管に効率的に到達することから,腸内細菌叢に影響を与える可能性も考えられる.高脂肪食マウスを用いた研究では,食事由来スフィンゴミエリンは,糞中のBifidobacterium属を増加させ,グラム陰性菌を減少させることによって,腸内細菌叢を変化させることが報告されている56).また,スフィンゴシンが腸管病原性のさまざまなグラム陰性および陽性菌に対して殺菌作用を示すことも報告されている57).複合スフィンゴ脂質の一つであるガングリオシドの乳児用調製粉乳への添加は,大腸菌を減らし,Bifidobacterium属を増やすことで糞便の細菌叢を改変する58).このようなガングリオシドの効果は,プレバイオティクスとして働くオリゴ糖部分によるものと考えられるが,特異的なセラミド部分と細菌の相互作用もまた関連するのかもしれない59).たとえばHelicobacter pyloriとスフィンゴミエリンのように,腸管毒性のある細菌やそれらの毒素とスフィンゴ脂質が結合することもある60, 61).したがって,食事由来スフィンゴ脂質は病原性細菌の過剰な増殖を制御し,それらと付着することで排泄の促進に寄与できるのかもしれない.
食事由来スフィンゴ脂質のなかでも特にスフィンゴミエリンは,コレステロールやトリアシルグリセロールの腸管吸収を濃度依存的に抑制することが動物実験で示されている62–66).このとき,スフィンゴミエリンの脂肪酸組成の違いが,脂質吸収阻害作用に影響するようである.牛乳由来スフィンゴミエリンは,ラットにおけるトリアシルグリセロールとコレステロールの腸管吸収の阻害について,卵由来のものよりも強力な作用を示し,その理由として,共存する脂質成分との強力な疎水性相互作用による可能性が考えられている65).スフィンゴミエリンやグルコシルセラミドの消化の過程で生じるセラミドやスフィンゴシンもまた,コレステロールや脂肪酸の吸収を阻害する66–68).
食事由来スフィンゴ脂質は,血清脂質を低下させることも報告されている.高脂血症となるマウス(APOE*3Leiden)に西洋食とさまざまなタイプのスフィンゴ脂質(スフィンゴミエリン,セラミド,スフィンゴシン,フィトスフィンゴシン)を摂取させた試験では,0.2~0.4%の食事由来スフィンゴ脂質の摂取は,血漿コレステロールとトリアシルグリセロールを20%以上低下させた.このとき,スフィンゴイド塩基は複合スフィンゴ脂質と同等の効果を示している66).また,遺伝的な肥満ラット(Zucker fattyラット)において,ニワトリ由来スフィンゴミエリンや植物由来グルコシルセラミドの4週間摂取により,血漿コレステロールの有意な低下が報告されている69).さらに筆者らも,ナマコ由来グルコシルセラミドの摂取がコレステロール負荷マウスの血清コレステロールを低下させることを確認している70).
ヒトにおいても,食事由来スフィンゴ脂質による脂質異常症改善の可能性が示されている.げっ歯類の胆汁と対照的に,ヒト胆汁はアルカリ性スフィンゴミエリナーゼを含むため,ヒトにおけるスフィンゴ脂質の消化効率は高いようである71).メタボリックシンドロームの成人男性に対して,フィトスフィンゴシンの4週間投与(1 g/日)は,有意に血漿LDLコレステロールを下げることが報告されている72).また,ホイップクリームとして40 g/日の乳脂摂取が,体重超過の成人におけるLDLコレステロールを有意に低下させることが示されており73),ホイップクリームに含まれる乳脂肪球皮膜のスフィンゴミエリンによる効果の可能性が考えられる.一方,健常成人における2週間の精製スフィンゴミエリンの摂取(1 g/日)は,non-HDLコレステロール濃度やコレステロール吸収には影響を与えないが,HDLコレステロールの有意な増加が認められている74).
本稿では食品としてのスフィンゴ脂質の存在や摂取量,消化吸収の機構,そして食品機能性について解説した.一方で,動脈硬化や糖尿病のリスクとセラミドの関係が近年注目されている75–77).血中セラミド濃度とインスリン抵抗性の関係78)や筋肉中のセラミドがインスリン抵抗性を惹起することなども報告されており79),スフィンゴ脂質摂取の危険性も危惧される.しかしながら,食事性スフィンゴ脂質は,前述したように生活習慣病に対してはむしろポジティブな効果も示されている.筆者らの動物実験による検討では,スフィンゴ脂質の2か月程度の摂取によっても筋肉中のセラミド濃度に大きな変動は確認されていない(未発表).セラミド合成を阻害することでインスリン抵抗性が改善することから79),内因性と外因性の作用の違いや,脂肪酸鎖長などの分子種の違いによって,その作用や役割が異なることも考えられる.
スフィンゴ脂質は,食品中の含有量やその摂取量も低いため,長い間食品成分や栄養素としては注目されていなかった.しかしながら本稿で示したように,食品機能成分としての認識が高まりつつあり,その作用機構も示されてきている.今後,さらなる研究成果によって,「スフィンゴ脂質栄養学」あるいは「スフィンゴ脂質食品機能学」が確立されることを大いに期待しながら,研究を推進していきたいと考える.
本稿で紹介した著者の研究成果の一部は,JSPS科研費16H04923および20H02931の助成を受けたものである.
1) 上沢 愛,中山雅浩,大西正男,新井基晴,菅原達也,伊藤精亮(1997)機能性素材として市販されている「植物セラミド」に含まれるスフィンゴ脂質の含有量と構成分の分析,帯広畜産大学学術研究報告,20, 199–206.
2) Sperling, P. & Heinz, E. (2003) Plant sphingolipids: structural diversity, biosynthesis, first genes and functions. Biochim. Biophys. Acta, 1632, 1–15.
3) Takakuwa, N., Kinoshita, M., Oda, Y., & Ohnishi, M. (2002) Existence of cerebroside in saccharomyces kluyveri and its related species. FEMS Yeast Res., 2, 533–538.
4) Ohashi, Y., Tanaka, T., Akashi, S., Morimoto, S., Kishimoto, Y., & Nagai, Y. (2000) Squid nerve sphingomyelin containing an unusual sphingoid base. J. Lipid Res., 41, 1118–1124.
5) Sugawara, T., Zaima, N., Yamamoto, A., Sakai, S., Noguchi, R., & Hirata, T. (2006) Isolation of sphingoid bases of sea cucumber cerebrosides and their cytotoxicity against human colon cancer cells. Biosci. Biotechnol. Biochem., 70, 2906–2912.
6) Mukhamedova, K.S. & Glushenkova, A.I. (2000) Structural analysis of ceramide-amino-ethyl-phosphonate in edible mediterranean cephalopods. Chem. Nat. Compd., 36, 329–341.
7) Vesper, H., Schmelz, E.M., Nikolova-Karakashian, M.N., Dillehay, D.L., Lynch, D.V., & Merrill, A.H. Jr. (1999) Sphingolipids in food and the emerging importance of sphingolipids to nutrition. J. Nutr., 129, 1239–1250.
8) Sugawara, T. & Miyazawa, T. (1999) Separation and determination of glycolipids from edible plant sources by high-performance liquid chromatography and evaporative light-scattering detection. Lipids, 34, 1231–1237.
9) Yunoki, K., Ogawa, T., Ono, J., Miyashita, R., Aida, K., Oda, Y., & Ohnishi, M. (2008) Analysis of sphingolipid classes and their contents in meals. Biosci. Biotechnol. Biochem., 72, 222–225.
10) Pham, P.H., Duffy, T.L., Dmytrash, A.L., Lien, V.W., Thomson, A.B., & Clandinin, M.T. (2011) Estimate of dietary ganglioside intake in a group of healthy Edmontonians based on selected foods. J. Food Compos. Anal., 24, 1032–1037.
11) Duan, R.D., Bergman, T., Xu, N., Wu, J., Cheng, Y., Duan, J., Nelander, S., Palmberg, C., & Nilsson, Å. (2003) Identification of human intestinal alkaline sphingomyelinase as a novel ecto-enzyme related to the nucleotide phosphodiesterase family. J. Biol. Chem., 278, 38528–38536.
12) Wu, J., Cheng, Y., Palmberg, C., Bergman, T., Nilsson, Å., & Duan, R.D. (2005) Cloning of alkaline sphingomyelinase from rat intestinal mucosa and adjusting of the hypothetical protein XP_221184 in GenBank. Biochim. Biophys. Acta, 1687, 94–102.
13) Wu, J., Liu, F., Nilsson, Å., & Duan, R.D. (2004) Pancreatic trypsin cleaves intestinal alkaline sphingomyelinase from mucosa and enhances the sphingomyelinase activity. Am. J. Physiol., 287, G967–G973.
14) Tomonaga, N., Manabe, Y., & Sugawara, T. (2017) Digestion of ceramide 2-aminoethylphosphonate, a sphingolipid from the jumbo flying squid Dosidicus gigas, in mice. Lipids, 52, 353–362.
15) Leese, H.J. & Semenza, G. (1973) On the identity between the small intestinal enzymes phlorizin hydrolase and glycosylceramidase. J. Biol. Chem., 248, 8170–8173.
16) Kobayashi, T. & Suzuki, K. (1981) The glycosylceramidase in the murine intestine. Purification and substrate specificity. J. Biol. Chem., 256, 7768–7773.
17) Sugawara, T., Kinoshita, M., Ohnishi, M., Nagata, J., & Saito, M. (2003) Digestion of maize sphingolipids in rats and uptake of sphingadienine by Caco-2 cells. J. Nutr., 133, 2777–2782.
18) Duan, J., Ishida, M., Aida, K., Tsuduki, T., Zhang, J., Manabe, Y., Hirata, T., & Sugawara, T. (2016) Dietary cerebroside from sea cucumber (Stichopus japonicus): absorption and effects on skin barrier and cecal short-chain fatty acids. J. Agric. Food Chem., 64, 7014–7021.
19) Kono, M., Dreier, J.L., Ellis, J.M., Allende, M.L., Kalkofen, D.N., Sanders, K.M., Bielawski, J., Bielawska, A., Hannun, Y.A., & Proia, R.L. (2006) Neutral ceramidase encoded by the Asah2 gene is essential for the intestinal degradation of sphingolipids. J. Biol. Chem., 281, 7324–7331.
20) Olsson, M., Duan, R.D., Ohlsson, L., & Nilsson, Å. (2004) Rat intestinal ceramidase: purification, properties, and physiological relevance. Am. J. Physiol., 287, G929–G937.
21) Duan, R.D., Cheng, Y., Yang, L., Ohlsson, L., & Nilsson, Å. (2001) Evidence for specific ceramidase present in the intestinal contents of rats and humans. Lipids, 36, 807–812.
22) Nilsson, Å. (1968) Metabolism of sphingomyelin in the intestinal tract of the rat. Biochim. Biophys. Acta, 164, 575–584.
23) Nilsson, Å. (1969) The presence of spingomyelin- and ceramide-cleaving enzymes in the small intestinal tract. Biochim. Biophys. Acta, 176, 339–347.
24) Buehrer, B.M. & Bell, R.M. (1993) Sphingosine kinase: properties and cellular functions. Adv. Lipid Res., 26, 59–67.
25) Van Veldhoven, P.P. & Mannaerts, G.P. (1993) Sphingosine kinase: properties and cellular functions. Adv. Lipid Res., 26, 69–98.
26) Gijiber, S., Van der Hoeven, G., & Van Veldhoven, P.P. (2001) Subcellular study of sphingoid base phosphorylation in rat tissues: evidence for multiple sphingosine kinases. Biochim. Biophys. Acta, 1532, 37–50.
27) Nakahara, K., Ohkuni, A., Kitamura, T., Abe, K., Naganuma, T., Ohno, Y., Zoeller, R.A., & Kihara, A. (2012) The Sjögren-Larsson syndrome gene encodes a hexadecenal dehydrogenase of the sphingosine 1-phosphate degradation pathway. Mol. Cell, 46, 461–471.
28) Kondo, N., Ohno, Y., Yamagata, M., Obara, T., Seki, N., Kitamura, T., Naganuma, T., & Kihara, A. (2014) Identification of the phytosphingosine metabolic pathway leading to odd-numbered fatty acids. Nat. Commun., 5, 5338.
29) Schmelz, E.M., Crall, K.J., Larocque, R., Dillehay, D.L., & Merrill, A.H. Jr. (1994) Uptake and metabolism of sphingolipids in isolated intestinal loops of mice. J. Nutr., 124, 702–712.
30) Nyberg, L., Nilsson, Å., Lundgren, P., & Duan, R.D. (1997) Localization and capacity of sphingomyelin digestion in the rat intestinal tract. J. Nutr. Biochem., 8, 112–118.
31) Larson, G., Falk, P., & Hoskins, L.C. (1988) Degradation of human intestinal glycosphingolipids by extracellular glycosidases from mucin-degrading bacteria of the human fecal flora. J. Biol. Chem., 263, 10790–10798.
32) Sugawara, T., Tsuduki, T., Yano, S., Hirose, M., Duan, J., Aida, K., Ikeda, I., & Hirata, T. (2010) Intestinal absorption of dietary maize glucosylceramide in lymphatic duct cannulated rats. J. Lipid Res., 51, 1761–1769.
33) Sugawara, T., Kinoshita, M., Ohnishi, M., Tsuzuki, T., Miyazawa, T., Nagata, J., Hirata, T., & Saito, M. (2004) Efflux of sphingoid bases by P-glycoprotein in human intestinal Caco-2 cells. Biosci. Biotechnol. Biochem., 68, 2541–2546.
34) Fujii, A., Manabe, Y., Aida, K., Tsuduki, T., Hirata, T., & Sugawara, T. (2017) Selective absorption of dietary sphingoid bases from the intestine via efflux by P-glycoprotein in rats. J. Nutr. Sci. Vitaminol. (Tokyo), 63, 44–50.
35) Morifuji, M., Higashi, S., Oba, C., Ichikawa, S., Kawahata, K., Yamaji, T., Itoh, H., Manabe, Y., & Sugawara, T. (2015) Milk phospholipids enhance lymphatic absorption of dietary sphingomyelin in lymph-cannulated rats. Lipids, 50, 987–996.
36) Breiden, B. & Sandhoff, K. (2014) The role of sphingolipid metabolism in cutaneous permeability barrier formation. Biochim. Biophys. Acta, 1841, 441–452.
37) van Smeden, J., Janssens, M., Gooris, G.S., & Bouwstra, J.A. (2014) The important role of stratum corneum lipids for the cutaneous barrier function. Biochim. Biophys. Acta, 1841, 295–313.
38) Akimoto, K., Yoshikawa, N., Higaki, Y., Kawashima, M., & Imokawa, G. (1993) Quantitative analysis of stratum corneum lipids in xerosis and asteatotic eczema. J. Dermatol., 20, 1–6.
39) Imokawa, G., Abe, A., Jin, K., Higaki, Y., Kawashima, M., & Hidano, A. (1991) Recharacterization of the nonlesional dry skin in atopic dermatitis through disrupted barrier function. J. Invest. Dermatol., 96, 523–526.
40) Macheleidt, O., Kaiser, H.W., & Sandhoff, K. (2002) Deficiency of epidermal protein-bound ω-hydroxyceramides in atopic dermatitis. J. Invest. Dermatol., 119, 166–173.
41) Imokawa, G., Yada, Y., Higuchi, K., Okuda, M., Ohashi, Y., & Kawamata, A. (1994) Pseudo-acylceramide with linoleic acid produces selective recovery of diminished cutaneous barrier function in essential fatty acid-deficient rats and has an inhibitory effect on epidermal hyperplasia. J. Clin. Invest., 94, 89–96.
42) Tsuji, K., Mitsutake, S., Ishikawa, J., Takagi, Y., Akiyama, M., Shimizu, H., Tomiyama, T., & Igarashi, Y. (2006) Dietary glucosylceramide improves skin barrier function in hairless mice. J. Dermatol. Sci., 44, 101–107.
43) Duan, J., Sugawara, T., Aida, K., Hirose, M., Sakai, S., Fujii, A., & Hirata, T. (2012) Dietary sphingolipids improve skin barrier functions via the upregulation of ceramide synthases in the epidermis. Exp. Dermatol., 21, 448–452.
44) Kuwata, T., Hashimoto, T., Ohto, N., Kuwahara, H., Lee, J.W., Bamba, T., & Mizuno, M. (2017) A metabolite of dietary glucosylceramide from pineapples improves the skin barrier function in hairless mice. J. Funct. Foods, 30, 228–236.
45) Kawano, K. & Umemura, K. (2013) Oral intake of beet extract provides protection against skin barrier impairment in hairless mice. Phytother. Res., 27, 775–783.
46) Haruta-Ono, Y., Setoguchi, S., Ueno, H.M., Higurashi, S., Ueda, N., Kato, K., Saito, T., Matsunaga, K., & Takata, J. (2012) Orally administered sphingomyelin in bovine milk is incorporated into skin sphingolipids and is involved in the water-holding capacity of hairless mice. J. Dermatol. Sci., 68, 56–62.
47) Morifuji, M., Oba, C., Ichikawa, S., Ito, K., Kawahata, K., Asami, Y., Ikegami, S., Itoh, H., & Sugawara, T. (2015) A novel mechanism for improvement of dry skin by dietary milk phospholipids: Effect on epidermal covalently bound ceramides and skin inflammation in hairless mice. J. Dermatol. Sci., 78, 224–231.
48) 平河聡,佐藤綾,服部祐子,松本 剛,横山浩治,金井 周(2013)米胚芽エキス配合粉末顆粒の摂取による全身の皮膚バリア機能に対する改善効果,薬理と治療,41, 1051–1059.
49) 野嶋潤,村上敏之,桑原浩誠,竹田竜嗣,渡邉泰雄(2014)パイナップル由来セラミド経口摂取による日本人女性の皮膚機能改善効果,応用薬理,87, 81–85.
50) Dillehay, D.L., Webb, S.K., Schmelz, E.M., & Merrill, A.H. Jr. (1994) Dietary sphingomyelin inhibits 1, 2-dimethylhydrazine-induced colon cancer in CF1 mice. J. Nutr., 124, 615–620.
51) Schmelz, E.M., Sullards, M.C., Dillehay, D.L., & Merrill, A.H. Jr. (2000) Colonic cell proliferation and aberrant crypt foci formation are inhibited by dairy glycosphingolipids in 1, 2-dimethylhydrazine-treated CF1 mice. J. Nutr., 130, 522–527.
52) Sugawara, T., Kinoshita, M., Ohnishi, M., & Miyazawa, T. (2002) Apoptosis induction by wheat-flour sphingoid bases in DLD-1 human colon cancer cells. Biosci. Biotechnol. Biochem., 66, 2228–2231.
53) Aida, K., Kinoshita, M., Sugawara, T., Ono, J., Miyazawa, T., & Ohnishi, M. (2004) Apoptosis inducement by plant and fungus sphingoid bases in human colon cancer cells. J. Oleo Sci., 53, 503–510.
54) Yamashita, S., Sakurai, R., Hishiki, K., Aida, K., & Kinoshita, M. (2017) Effects of dietary plant-origin glucosylceramide on colon cytokine contents in DMH-treated mice. J. Oleo Sci., 66, 157–160.
55) Yamashita, S., Seino, T., Aida, K., & Kinoshita, M. (2017) Effects of plant sphingolipids on inflammatory stress in differentiated Caco-2 cells. J. Oleo Sci., 66, 1337–1342.
56) Norris, G.H., Jiang, C., Ryan, J., Porter, C.M., & Blesso, C.N. (2016) Milk sphingomyelin improves lipid metabolism and alters gut microbiota in high fat diet-fed mice. J. Nutr. Biochem., 30, 93–101.
57) Sprong, R.C., Hulstein, M.F., & Van der Meer, R. (2001) Bactericidal activities of milk lipids. Antimicrob. Agents Chemother., 45, 1298–1301.
58) Rueda, R., Sabatel, J.L., Maldonado, J., Molina-Font, J.A., & Gil, A. (1998) Addition of gangliosides to an adapted milk formula modifies levels of fecal Escherichia coli in preterm newborn infants. J. Pediatr., 133, 90–94.
59) Martin, M.J., Martin-Sosa, S., Alonso, J.M., & Hueso, P. (2003) Enterotoxigenic Escherichia coli strains bind bovine milk gangliosides in a ceramide-dependent process. Lipids, 38, 761–768.
60) Lingwood, C.A., Boyd, B., & Nutikka, A. (2000) Analysis of interactions between glycosphingolipids and microbial toxins. Methods Enzymol., 312, 459–473.
61) Gupta, V.R., Patel, H.K., Kostolansky, S.S., Ballivian, R.A., Eichberg, J., & Blanke, S.R. (2008) Sphingomyelin functions as a novel receptor for Helicobacter pylori VacA. PLoS Pathog., 4, e1000073.
62) Nyberg, L., Duan, R.D., & Nilsson, A. (2000) A mutual inhibitory effect on absorption of sphingomyelin and cholesterol. J. Nutr. Biochem., 11, 244–249.
63) Eckhardt, E.R., Wang, D.Q., Donovan, J.M., & Carey, M.C. (2002) Dietary sphingomyelin suppresses intestinal cholesterol absorption by decreasing thermodynamic activity of cholesterol monomers. Gastroenterology, 122, 948–956.
64) Noh, S.K. & Koo, S.I. (2003) Egg sphingomyelin lowers the lymphatic absorption of cholesterol and α-tocopherol in rats. J. Nutr., 133, 3571–3576.
65) Noh, S.K. & Koo, S.I. (2004) Milk sphingomyelin is more potent inhibitor than egg sphingomyelin of intestinal absorption of cholesterol and α-tocopherol in rats. J. Nutr., 134, 2611–2616.
66) Duivenvoorden, I., Voshol, P.J., Rensen, P.C., van Duyvenvoorde, W., Romijn, J.A., Emeis, J.J., Havekes, L.M., & Nieuwenhuizen, W.F. (2006) Dietary sphingolipids lower plasma cholesterol and triacylglycerol and prevent liver steatosis in APOE*3Leiden mice. Am. J. Clin. Nutr., 84, 312–321.
67) Garmy, N., Taieb, N., Yahi, N., & Fantini, J. (2005) Interaction of cholesterol with sphingosine physicochemical characterization and impact on intestinal absorption. J. Lipid Res., 46, 36–45.
68) Feng, D., Ohlsson, L., Ling, W., Nilsson, A., & Duan, R.D. (2010) Generating ceramide from sphingomyelin by alkaline sphingomyelinase in the gut enhances sphingomyelin-induced inhibition of cholesterol uptake in Caco-2 cells. Dig. Dis. Sci., 55, 3377–3383.
69) Yunoki, K., Renaguli, M., Kinoshita, M., Matsuyama, H., Mawatari, S., Fujino, T., Kodama, Y., Sugiyama, M., & Ohnishi, M. (2010) Dietary sphingolipids ameliorate disorders of lipid metabolism in Zucker fatty rats. J. Agric. Food Chem., 58, 7030–7035.
70) Hossain, Z., Sugawara, T., Aida, K., & Hirata, T. (2011) Effect of dietary glucosylceramide from sea cucumber on plasma and liver lipids in cholesterol-fed mice. Fish. Sci., 77, 1081–1085.
71) Ohlsson, L., Hertervig, E., Jonsson, B.A., Duan, R.D., Nyberg, L., Svernlöv, R., & Nilsson, A. (2010) Sphingolipids in human ileostomy content after meals containing milk sphingomyelin. Am. J. Clin. Nutr., 91, 672–678.
72) Snel, M., Sleddering, M.A., Pijl, H., Nieuwenhuizen, W.F., Frölich, M., Havekes, L.M., Romijn, J.A., & Jazet, I.M. (2010) The effect of dietary phytosphingosine on cholesterol levels and insulin sensitivity in subjects with the metabolic syndrome. Eur. J. Clin. Nutr., 64, 419–423.
73) Rosqvist, F., Smedman, A., Lindmark-Mansson, H., Paulsson, M., Petrus, P., Straniero, S., Rudling, M., Dahlman, I., & Risérus, U. (2015) Potential role of milk fat globule membrane in modulating plasma lipoproteins, gene expression, and cholesterol metabolism in humans: a randomized study. Am. J. Clin. Nutr., 102, 20–30.
74) Ramprasath, V.R., Jones, P.J., Buckley, D.D., Woollett, L.A., & Heubi, J.E. (2013) Effect of dietary sphingomyelin on absorption and fractional synthetic rate of cholesterol and serum lipid profile in humans. Lipids Health Dis., 12, 125.
75) Meikle, P.J. & Summers, S.A. (2017) Sphingolipids and phospholipids in insulin resistance and related metabolic disorders. Nat. Rev. Endocrinol., 13, 79–91.
76) Hannun, Y.A. & Obeid, L.M. (2017) Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol., 19, 175–191.
77) Summers, S.A., Chaurasia, B., & Holland, W.L. (2019) Metabolic messengers: ceramides. Nat. Metab., 1, 1051–1058.
78) Lemaitre, R.N., Yu, C., Hoofnagle, A., Hari, N., Jensen, P.N., Fretts, A.M., Umans, J.G., Howard, B.V., Sitlani, C.M., Siscovick, D.S., et al. (2018) Circulating sphingolipids, insulin, HOMA-IR, and HOMA-B: the strong heart family study. Diabetes, 67, 1663–1672.
79) Turpin-Nolan, S.M., Hammerschmidt, P., Chen, W., Jais, A., Timper, K., Awazawa, M., Brodesser, S., & Brüning, J.C. (2019) CerS1-derived C18: 0 ceramide in skeletal muscle promotes obesity-induced insulin resistance. Cell Rep., 26, 1–10.

京都大学大学院農学研究科教授.博士(農学).
1968年東京都に生る.91年東北大学農学部卒業.93年同大学院農学研究科博士前期課程修了.同年日本油脂株式会社勤務.2000年東北大学大学院農学研究科博士後期課程修了.同年生研機構派遣研究員(食品総合研究所勤務).02年科学技術特別研究員(国立健康・栄養研究所勤務).04年京都大学大学院農学研究科助教授(07年より准教授).13年より現職.05年日本栄養・食糧学会奨励賞.
研究テーマと抱負海洋生物に含まれる機能性物質の探索と作用機序の解明.スフィンゴ脂質やカロテノイドなどの機能性脂質をキーワードとする.とくに消化吸収機構を含めた食品機能性の解明を目指す.
ウェブサイトhttp://www.bioproducts.marine.kais.kyoto-u.ac.jp/
趣味テニス,美術鑑賞.

This page was created on 2020-08-31T15:46:01.73+09:00
This page was last modified on 2020-09-28T08:19:54.000+09:00
このサイトは(株)国際文献社によって運用されています。