長いDNAをわずか直径10マイクロメートルに満たない核の中に収納するために必須なクロマチンの構造は,DNAを介したさまざまな細胞現象を制御する.たとえば遺伝子の転写を例にとっても,クロマチンがオープンな状態にある遺伝子は転写が可能であるが,凝集していると転写が抑制される.そのため,クロマチンを構成する分子は遺伝子の転写を制御する重要な因子であると同時に,DNAの複製や修復などを介して,広範な生命現象を制御する.
HMGA(high-mobility group AT-hook)タンパク質は多数のクロマチン構造分子の一つで,ヒトやマウスではHMGA1, HMGA2という二つの遺伝子にコードされている1).HMGA1遺伝子からは選択的スプライシングによってHMGA1aとHMGA1bという2種類のタンパク質が産生される.もともとは,クロマチンに結合する小さい核内分子として生化学的に分離され2),その後AT-hookドメインを介してAT配列に富んだDNAの副溝(マイナーグルーヴ)に結合することが明らかになった3, 4).また,HMGA2の全身ノックアウトマウスは体が小さくなるピグミーマウスであることが知られていた5).さらに最近の解析から,HMGA1とHMGA2ダブルノックアウトマウスは体重が5グラム程度(通常の1/4),体長が5センチメートル程度(通常の1/3)しかないスーパーピグミーマウスになることがわかった6).驚いたことにこのような小さいマウスでもメンデルの法則の約半分の割合で産まれてきて,少なくとも12か月齢までは生存する.つまり,体の大きさの制御には重要なものの,多くの組織において,少なくとも個体の生存のための機能には必須でないということである.遺伝子欠損によりこのような非常に興味深い表現型を示すHMGAタンパク質だが,どのような分子機能を有しているのだろうか.
HMGAタンパク質の生化学的な機能については,1973年に発見されて以来,2000年代初めにかけて精力的な研究が行われ,DNAとの結合様式やクロマチン構造への影響,そして遺伝子の転写に与える影響が主に試験管内の実験により明らかになっていた1).一方HMGAタンパク質の個体にとっての役割は,1995年のHMGA2ノックアウトマウスの作製以降,ノックアウトやノックダウンなど遺伝子操作技術の発展に伴い,さまざまな細胞現象や発生,疾患に関わることが明らかとなってきた7, 8).しかし,個体で果たすマクロな役割に,どういったHMGAタンパク質のミクロな生化学的機能が寄与しているのかはいまだ不明である.本稿では,2000年代初めまでに主に試験管内で明らかになっていたHMGAタンパク質の分子機能についての報告を紹介したのちに,それが細胞や個体レベルのどういった現象に関与しているかを,実際のHMGAタンパク質の生理機能に関連づけながら議論したい.
なお,HMGAの表記について本来はヒトではHMGA,マウスではHmgaと記載するのが適当であるが,紹介するほとんどの解析でヒトHMGAを用いているためすべてHMGAで統一する.
2. HMGAタンパク質の構造とクロマチンとの相互作用
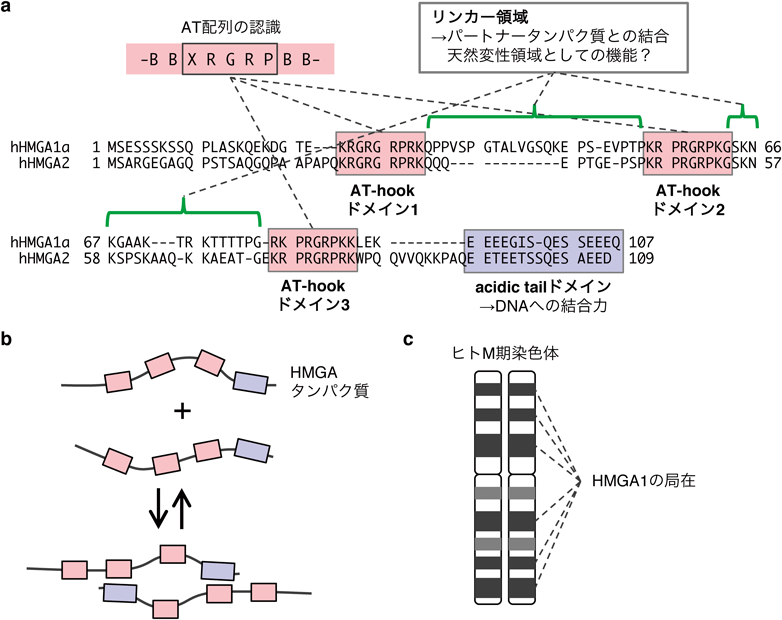
HMGA1a, HMGA1b, HMGA2の3種類のHMGAタンパク質は,わずか100アミノ酸程度の小さいタンパク質で,その中に名前の由来にもなっているBBXRGRPBB(BはKあるいはR, XはGあるいはP)という配列で構成されるAT-hookドメインを三つ持つ4)(図1a).このドメインは塩基性アミノ酸残基が連なり,ATに富んだDNAの副溝に結合するHMGAタンパク質のDNA結合領域であることが,フットプリントアッセイを用いた当初の研究により明らかになった4).また,HMGA1のAT-hookドメインのどこがAT配列の認識に重要なのかをNMRにより調べたところ,XRGRP配列だけでAT配列の認識に十分であることがわかった9).さらに,この二つのアルギニン残基をリシン残基に置換するとAT配列認識能が低下したため,この二つのアルギニン残基がAT-hookのATの認識に重要であることがわかった9).またその後のX線結晶構造解析からも,RGR配列が副溝に強く結合し,特に二つ目のRがチミジン塩基と結合していたこともわかり,アルギニン残基の重要性が支持された10).
HMGAタンパク質で重要なドメインとして,C末端には酸性アミノ酸残基が連なったacidic tailがある.興味深いことに,このC末端を除いたHMGA2変異体を用いてゲルシフトアッセイを行ったところ,ATを含むDNAへの結合力が低下した11).すなわち,HMGAタンパク質のATへの特異性はAT-hookが決めている一方で,acidic tailはHMGAタンパク質のDNA結合力を上げていると考えられる.一方で,負電荷を帯びた酸性領域が,同じく酸性のDNAへの結合力を上げるのは少し不思議である.ではacidic tailはHMGAタンパク質のどういった性質に寄与しているのだろうか?
最近の報告でHMGA2を用いて試験管内でクロスリンク実験やFRET[蛍光共鳴エネルギー移動(fluorescence resonance energy transfer)]実験を組み合わせることで,HMGA2がホモ二量体を形成することが明らかとなった12).また,acidic tailを欠損するとこの二量体化活性が喪失したことからacidic tailはホモ二量体形成に必須なことがわかった.また,詳細な解析から酸性のacidic tailと塩基性のAT-hookによる静電的相互作用がホモ二量体化のメカニズムであると考えられた(図1b).このacidic tailの機能を考慮すると,もしかしたらホモ二量体を形成することが,HMGAタンパク質が強くDNAに結合することに重要なのかもしれない.しかし,HMGAタンパク質二量体形成の生理的機能はいまだ明らかではなく,どういう場面でホモ二量体形成が重要な役割を果たすのか興味深い.
AT-hookとacidic tailに挟まれたリンカー領域はさまざまなパートナータンパク質と結合する領域として知られている13).またこの領域は,特別な構造をとらないと考えられており,HMGAタンパク質に天然変性タンパク質としての性質を与えている可能性もある.近年さまざまな天然変性タンパク質が液-液相分離を誘導して細胞内に機能ドメインを形成することが知られており,リンカー領域を介してHMGAタンパク質も液-液相分離を誘導していたら興味深い.
ではHMGAタンパク質は,DNAではなくクロマチンとはどのように相互作用しているのだろうか? 当初からクロマチン相互作用因子として同定されたことからも想像できるとおり,HMGA1はヌクレオソームと直接結合することがわかっているが,特にAT配列が外側を向いているヌクレオソームでより結合が強くなる14, 15).また,クロスリンクの実験から,ヌクレオソームと結合するときにはコアヒストンの中でもヒストンH2A,ヒストンH2B,ヒストンH3と近接することが明らかとなっており,コアヒストンに直接結合している可能性がある14).興味深いことに,HMGA1が結合するとヒストンへのDNAの結合様式が変化することがわかっており15),HMGAタンパク質が結合することでクロマチン構造が変化することが示唆される.
HMGAタンパク質の核内でのグローバルな局在については,古くはトリプシン/ライト染色によるGバンディングにより調べられている16).その結果,M期染色体においてHMGA1はヘテロクロマチン領域と考えられるダークバンド上に共局在することが示唆された(図1c).また,その後の詳細な解析からM期染色体上でHMGA1が結合するのは,SAR(scaffold attachment region)と呼ばれる領域であることが示された17).AT-richな配列で構成されるSARは,M期染色体のバックボーンとして働く一方で,遺伝子を含むDNAのループ構造の基盤を形成し,機能性DNAエレメントとしても機能することが知られている18).このように,過去に免疫染色で大雑把に示されていた,HMGAタンパク質がヘテロクロマチン領域に局在するという事実は,最近のES細胞におけるHMGAタンパク質のシークエンス解析によって検証された19).この報告では,ビオチン化したHMGA1とHMGA2をプルダウンし,結合配列をシークエンス解析し,その局在とヒストン修飾などのさまざまなクロマチンの特徴の相関を調べている.その結果,HMGAタンパク質が局在するゲノム領域は調べたヒストン修飾のうちヘテロクロマチン領域であるH3K9me2(ヒストンH3リシン9のジメチル化)とのみ相関し,H3K4meやH3K36me3, RNAポリメラーゼII, DNase感受性領域などのユークロマチン領域とは相関しなかった.これらの結果から,HMGAタンパク質はゲノムワイドにはヘテロクロマチン領域に局在することがわかった.
3. HMGAタンパク質による遺伝子転写の制御機構
上記のようにゲノムワイドにはヘテロクロマチン領域に局在する一方で,HMGAタンパク質は個々の標的遺伝子座に結合して転写を正に制御することが,主に試験管内転写のシステムを用いた研究でわかっている1).よく研究されている制御メカニズムは主に三つある.
1)エンハンソソーム形成とそのための場の提供
IL-2受容体(IL-2Rα)やインターフェロンβ(IFN-β)をコードする遺伝子は上流の刺激に応答してすばやく転写されるが,その応答性を実現するために転写開始点上流に特別なDNAエレメントが存在することが知られている.IL-2Rα遺伝子座では,PRRI(positive regulatory region I),PRRII, PRRIIIが存在し,これらのエレメント上に転写に重要な転写因子であるNF-κB, SRF, GAT A, Stat5, Elf-1に加えてHMGA1が集合し,エンハンソソーム(enhanceosome)という複合体を形成することで転写が亢進する20, 21).三つのPRRの中で,PRRIIにはElf-1というT細胞特異的な転写因子が結合する.PRRII配列を含む再構成ヌクレオソームを用いてフットプリントアッセイを行ったところ,まず通常の状態ではElf-1結合領域はヌクレオソーム内にあり,Elf-1は結合できないことがわかった22).一方で,HMGA1はPRRIIに結合することができ,その後の刺激に応答してヌクレオソームを移動させ,Elf-1を呼び込み,エンハンソソームを形成する場を提供していることが示唆されている.またHMGA1は,ヌクレオソームのリモデリングによるエンハンソソーム形成の場を提供する機能に加えて,エンハンソソームの形成にも寄与していると考えられている1).それぞれのステップにおいてHMGA1のどういった性質が寄与しているのかは不明であるが,ヌクレオソームのリモデリングにはHMGAタンパク質が結合することによるヌクレオソーム構造の変化が15),エンハンソソームの形成にはリンカー領域を介した相互作用タンパク質との結合が重要な役割を果たしている可能性がある13).
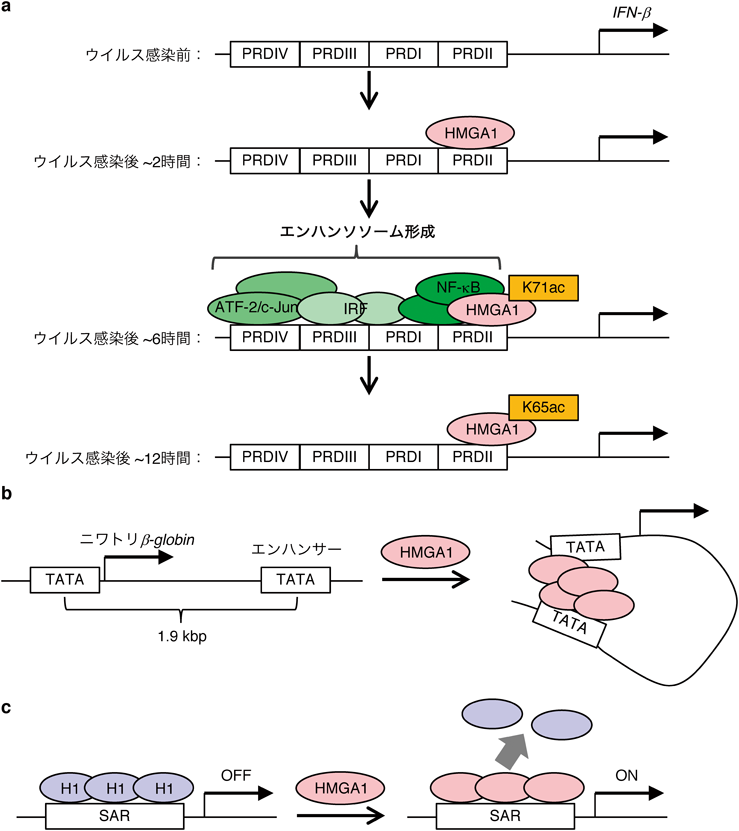
IFN-β遺伝子座にあるPRDI(positive regulatory domain II),PRDII, PRDIII, PRDIVと呼ばれるエレメントでもHMGA1によるNF-κB, ATF-2/c-Jun, IRFを含むエンハンソソームの形成が重要である23–25).IL-2α遺伝子座と同様,HMGA1がPRDIIに結合してヌクレオソームのリモデリングとエンハンソソームを構成するタンパク質の集合の両方に重要だと考えられているが,こちらの遺伝子座ではHMGA1タンパク質のアセチル化によるダイナミックな制御が知られている.ウイルス感染の下流で起きるIFN-βの転写にはヒストンアセチル化酵素であるPCAFやCBPが重要な役割を果たすが,PCAFはHMGA1タンパク質のK71を,CBPはK65をアセチル化することがわかった26).クロマチン免疫沈降(chromatin immunoprecipitation:ChIP)実験によりこれらのアセチル化されたHMGA1がIFN-β遺伝子座に結合するタイミングを調べると,HMGA1タンパク質自体は感染後2時間後から24時間後まで常に強く結合していた.それに対して,HMGA1上のK71のアセチル化はウイルス感染後6時間くらいをピークに,一方K65のアセチル化は12時間くらいをピークに一過的に増加することがわかった27)(図2a).エンハンソソームを形成するNF-κB, c-Jun, ATF-2, IRF-1はウイルス感染後6時間をピークに結合していたことから,K71のアセチル化が遺伝子発現と相関していると考えられた.そこで,エンハンソソーム形成におけるK65とK71のアセチル化の役割をフットプリントアッセイで調べたところ,K71のアセチル化はエンハンソソーム形成を促進した一方で,K65のアセチル化はエンハンソソーム形成を阻害した26, 27).以上のことから,ウイルス感染後,始めに施されるK71のアセチル化は転写を亢進し,その後のK65のアセチル化によってエンハンソソームが崩壊,転写が終息する,という制御が示唆された.では,K65とK71のアセチル化はどのようにしてエンハンソソームの形成を制御しているのだろうか? K65とK71は二つ目と三つ目のAT-hookドメインの間に位置するため,そのアセチル化はDNAとの結合より他のタンパク質との相互作用に影響している可能性が考えられる.そこで実際に,K65あるいはK71がアセチル化されたHMGA1とエンハンソソームの構成因子であるNF-κB, ATF-2との結合を調べてみた.その結果,K71にアセチル化があると相互作用が強く,逆にK65にアセチル化があると弱くなることがわかった27).すなわち,K71あるいはK65のアセチル化はエンハンソソームの形成を,それぞれ正あるいは負に制御することで,IFN-β遺伝子の転写をダイナミックに制御していることが示唆された.このようにHMGAタンパク質が翻訳後修飾でその機能を変化させ,刻々と変化する遺伝子の転写状態を制御しているのはとても興味深い.
2)エンハンサー・プロモーター相互作用の形成
遺伝子の転写を亢進するDNAエレメントであるエンハンサーは,現在は制御する遺伝子のプロモーター領域に物理的に相互作用して機能することがわかっている28).HMGAタンパク質は,このエンハンサー・プロモーター相互作用を媒介する因子として当初から研究されている.ニワトリのβ-globin遺伝子座には,TAT Aボックスモチーフがプロモーター領域とプロモーターの1.9 kbp下流のエンハンサー領域に存在し,赤血球系特異的転写因子であるGAT A1が転写を活性化する29).試験管内の実験において,β-globin遺伝子座を含むプラスミドDNAと,Hela細胞から精製したGAT A1を含むRNAポリメラーゼII,そしてHMGA1を加えて転写活性を調べた30).すると,エンハンサーのないプラスミドDNA上ではHMGA1は転写を阻害した.一方で,エンハンサーがあるプラスミドDNA上では転写を著しく亢進した.重要なことに電子顕微鏡にてβ-globin遺伝子座を含むDNAの構造を調べると,HMGA1を加えるだけでエンハンサーとプロモーターを物理的に結合させているような像が得られた.これらの結果から,HMGA1はプロモーターとエンハンサーのTAT Aボックスモチーフを結合させており,それがエンハンサー活性を媒介している,ということが考えられる(図2b).上述したように,HMGAタンパク質はホモ二量体を形成することがわかったため,AT-hookドメインでTAT Aモチーフに結合したHMGAタンパク質がacidic tailを介してホモ二量体を形成することが,このエンハンサーとプロモーターの結合に重要なのかもしれない12).
3)クロマチン上でのリンカーヒストンH1との競合作用
リンカーヒストンH1は,ヌクレオソームの高次構造形成を介して遺伝子の転写を抑制するクロマチン分子である31).HMGAタンパク質による遺伝子転写制御機構の一つとして,ヒストンH1との競合により遺伝子の転写を促進する可能性を示唆する報告もある32).上述のように機能性DNAエレメントとして働くSAR配列にT7プロモーターを結合させた人工遺伝子を試験管内で転写させるとき,ヒストンH1を添加するとその転写が抑制された.一方で,その反応系にHMGA1を加えると,ヒストンH1による転写抑制が脱抑制することがわかった(図2c).このとき,SAR配列に結合していたヒストンH1がHMGA1により剥がれたことから,HMGA1はヒストンH1とDNAの結合を競合することでSARからの転写を促進していたことが示唆された.同様のことが細胞内でも起きることは,その後のFRAP[光褪色後蛍光回復法(fluorescence recovery after photobleaching)]実験を用いた解析でも示唆されている33).この報告では,ヒストンH1にGFPを融合させたH1-GFPを発現させた細胞において,FRAP実験でH1-GFPとクロマチンの結合へのHMGA1の影響を調べた.すると,コントロールに比べて,HMGA1を過剰発現させた細胞ではH1-GFPの交換が速い,すなわちH1-GFPとクロマチンの結合が弱まっていることが示唆された.これまでに,HMGAタンパク質がH1と競合して遺伝子の転写を促進している内在遺伝子の例は知られていないが,このような制御がいずれかの遺伝子座で起きている可能性がある.
以上の三つの例のようにHMGAタンパク質は他の転写因子やクロマチン因子,あるいは機能性エレメントと相互作用あるいは競合することで遺伝子の転写を亢進することがよく研究されている.一方で,HMGAタンパク質は遺伝子の転写を抑制することも知られている.たとえば,DNAへの結合を競合する相手が転写を活性化する因子であれば,結果的にHMGAタンパク質はその転写を抑制する.実際に,HMGA1はIL-4プロモーターにおけるNFATの結合や,Hoxd9のホメオボックスドメインのDNA結合と競合して,転写を阻害することも知られている34, 35).
また,上記のβ-globinの転写においてはエンハンサー非存在条件で,SARにおけるH1との競合を示した報告ではH1が存在しない条件で,それぞれHMGA1の添加は試験管内転写を抑制することがわかっている30, 32).いずれの報告もヒストンが存在しないDNAに対する影響であるが,HMGAタンパク質がヘテロクロマチンによく結合していることなどを考えると,HMGAタンパク質がDNAに結合することそのものが遺伝子の転写に負に働くことはそれほど不思議ではない.HMGAタンパク質自体は転写を負に制御するが,結合する遺伝子座によって転写促進因子を誘導する,エンハンサーとの相互作用を引き起こす,あるいは競合する相手がヒストンH1のようにHMGAタンパク質よりもより転写抑制機能が強い場合は転写を亢進する,というのがHMGAタンパク質の遺伝子転写に対する作用の本質なのかもしれない.
4. HMGAタンパク質の細胞・個体での機能とその基盤となる生化学的性質
上述のとおり,ノックアウトやノックダウンなど遺伝子操作技術が発展してきた2000年前後から,HMGAタンパク質の研究はそれまでの試験管内での生化学的性質の解明から細胞や個体での機能解析に移ってきた7, 8).その結果,HMGAタンパク質は個体サイズの制御に加えて,細胞老化や上皮間葉転換,幹細胞の維持といった細胞現象や,神経系・造血系・筋肉などの発生,がん化や糖尿病などの疾患など,多くの生理機能を制御していることが明らかとなってきている.ここではHMGAタンパク質が制御する数ある生理機能の中から,特にクロマチン構造の制御と関連が研究されている細胞老化と神経発生での役割に焦点を当て,これらの現象におけるHMGAタンパク質の分子機能について議論したい.
1)細胞老化におけるHMGAタンパク質の役割
細胞周期が不可逆的に停止する細胞老化は,細胞分裂を限界まで繰り返すことや,DNA損傷,がん遺伝子の異所的発現などによって引き起こされる36).興味深いことに,がん遺伝子の発現によって引き起こされる細胞老化はDNA染色のレベルで可視化できる大規模なクロマチン構造変化を伴うことがわかっている37).すなわち,ヒト線維芽細胞に恒常活性化型のRas変異体を過剰発現する,あるいは長期間培養することで細胞老化を誘導し,DAPIにてDNA染色を行うとコントロールでは観察できないような明暗のはっきりしたDAPIの集合体が観察された.この集合体は細胞老化マーカーと相関し,細胞老化の実行因子であるp16の過剰発現でも観察されること,またヘテロクロマチンマーカーであるH3K9meやHP1と共局在したことから,SAHF(senescence-associated heterochromatic foci)と名づけられた.
その後の報告でこのSAHF形成にHMGAタンパク質が重要であることが明らかとなった38).Ras変異体の過剰発現により細胞老化が誘導されるとき,HMGA1とHMGA2の発現が誘導され,どちらのタンパク質もSAHFに共局在した.このとき,HMGA1あるいはHMGA2をノックダウンすると,Rasによって誘導されるSAHF陽性細胞が減少したため,HMGAタンパク質がSAHF形成に重要であることがわかった.興味深いことに,HMGA1とp16をノックダウンすると協調的に細胞周期が回復したことから,両者は協調的に細胞老化を誘導している,すなわちSAHF形成は細胞老化の単なる副産物ではなく,細胞老化における細胞周期の停止に重要な役割を果たすことが示唆された.SAHF形成は細胞老化という細胞周期を停止した「不活性な」状態を維持するための機構だと考えられており,現在では細胞老化の一つの判断基準としても捉えられている.
ではHMGAタンパク質はどのようにしてこのSAHFの形成を誘導しているのだろうか? そのためにはまず,SAHFがどう構築されているのかを知ることが必要である.最近,RasによってSAHFが誘導された細胞において網羅的なクロマチン構造解析が行われた39–41).この報告ではまずHi-Cによりゲノム間相互作用を解析した結果,ヘテロクロマチン領域であるBコンパートメントどうしの結合が強くなること,特にゲノム一次配列上で近接していないBコンパートメントどうしの結合が強くなることが明らかとなった41).また,ヘテロクロマチン領域は核膜に結合していることが知られているが,核膜に結合しているクロマチンドメイン(lamin-associated domain:LAD)の解析を行った結果,細胞老化の誘導に伴い,核内膜構成因子であるLamin B1の発現が低下し,SAHFを形成するヘテロクロマチンドメインが核膜周囲から離れる様子が観察された40, 41).エピジェネティクスの変化については,SAHF上のヒストン修飾を免疫染色によって調べると,SAHF中心にH3K9me3が,その周りにリング上にH3K27me3が集積していた39).重要なことに,ゲノム上でのH3K9me3, H3K27me3の分布をChIP-seqにより調べると,細胞老化誘導前後でこれらの分布はほとんど変化していないことがわかった.これらの結果を合わせると,Ras変異体の過剰発現によるSAHFは,すでに存在するH3K9me3やH3K27me3でマークされるヘテロクロマチン領域が,核膜周囲から離れて核質で集合することで形成されていると考えられる.重要なことに,Lamin B1のノックダウンとHMGA1あるいはHMGA2の過剰発現を組み合わせることによって,SAHFの形成が誘導された40).このことから,HMGAタンパク質は核膜から剥がれたヘテロクロマチン領域を集合させることでSAHF形成を誘導している可能性がある.細胞老化を誘導した細胞においてHMGA1をノックダウンしてもH3K9me3やH3K27me3のゲノム上の分布は変わらなかったことからも,HMGAタンパク質はヘテロクロマチンの形成そのものではなく,その局在か集合の制御に寄与していることが示唆される.HMGAタンパク質は非常に小さい分子であるが,これまでの報告のとおりAT-richなヘテロクロマチン領域に結合し19),acidic tailを介してホモ二量体を形成することで12),ヘテロクロマチンの集合,ひいてはSAHFのような巨大な構造体を誘導している可能性がある.
もう一つのSAHF形成におけるHMGAタンパク質の役割としては,ヒストンH1の制御が考えられる.Ras変異体の過剰発現による細胞老化の過程では,ヒストンH1が顕著に減少することが観察されており,ヒストンH1の存在が細胞老化やSAHF形成に阻害的に働いていることを示唆している42).このヒストンH1の減少は翻訳後の分解によって起きていることが示唆されており,通常の過剰発現実験ではヒストンH1の減少の細胞老化やSAHF形成における役割は明らかにできず,いまだその意義は検証されていない.ただ興味深いことに,ヒストンH1のN末端にGFPを融合したヒストンH1(GFP-H1)を過剰発現すると細胞周期の低下が観察され,そこにさらにHMGA2を過剰発現するとSAHF形成が起きることがわかった.C末端にGFPを融合したヒストンH1(H1-GFP)ではこのような減少は起きなかった.人工的なシステムで解釈は難しいが,GFP-H1は何かしら内在のヒストンH1の機能を阻害することで細胞周期の低下,SAHF形成に寄与している可能性がある.そして,HMGAタンパク質はヒストンH1をクロマチン上から剥がす機能があることを考えると32),細胞老化の過程でHMGAタンパク質はヒストンH1をクロマチン上から剥がし,結果的にヒストンH1の分解を誘導することで細胞老化やSAHF形成に寄与している可能性がある.ヒストンH1とHMGAタンパク質によって形成されるクロマチンが,それぞれどのような構造なのか,核膜との相互作用にどのような違いがあるのかなどは不明だが,それが明らかになれば,HMGAタンパク質がSAHF形成において果たす役割の全体像が明らかになるだろう.
2)神経発生におけるHMGAタンパク質の役割
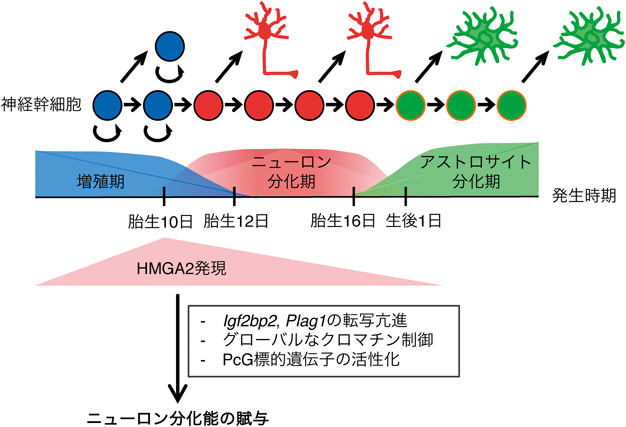
我々の大脳を構成するニューロンやグリア細胞は,そのほとんどが発生期に共通の神経幹細胞から産生される43–46).興味深いことに,神経幹細胞は常にニューロンやグリア細胞を産生しているわけではなく,時期依存的にその分化運命を転換させることがわかっている.たとえばマウス大脳新皮質においては,胎生10日目くらいまでは一つの神経幹細胞が二つの神経幹細胞に分裂する非対称分裂を行って神経幹細胞の数を増やす(増殖期)(図3).胎生10日目以降は,ニューロン分化能を獲得して一つの神経幹細胞が一つの神経幹細胞と一つのニューロン前駆細胞を産み出す非対称分裂を行ってニューロンを産生する(ニューロン分化期).そして,出産前後になるとニューロン分化能を失い,グリア細胞の一種であるアストロサイトを産み出す(アストロサイト分化期).この分化運命の転換タイミングが異常になると,正常な数のニューロンやグリア細胞が産生されなくなってしまう.そのため,我々の大脳が正しく機能するためには神経幹細胞の分化運命転換タイミングを正しく制御する必要がある.
これまでの研究から,大脳新皮質の神経幹細胞の分化運命転換にはさまざまなクロマチン因子が関わることがわかっている44, 47–49).その中で,HMGAタンパク質が重要な働きを果たすことが明らかとなっている.ニューロン分化期からアストロサイト分化期の大脳新皮質において,HMGA1とHMGA2は,徐々に発現が低下することがわかっている50–52).そこで,HMGAタンパク質の役割を調べるために初代培養のニューロン分化期神経幹細胞でHMGA1とHMGA2をノックダウンしたところ,ニューロンに分化できる神経幹細胞が減少し,アストロサイトに分化する神経幹細胞が増加した52).逆に,HMGA1あるいはHMGA2をアストロサイト分化期神経幹細胞に過剰発現したところ,ニューロンに分化できる神経幹細胞が増加した.このことは,初代培養系の神経幹細胞だけでなく,生体内の神経幹細胞に子宮内電気穿孔法で遺伝子導入を行っても観察され,HMGA1とHMGA2のノックダウンによりニューロン分化が減少し,過剰発現によりニューロン分化が増加した.興味深いことに,生後の本来アストロサイトを主として産生する神経幹細胞にHMGA1とHMGA2を遺伝子導入すると,ニューロンに分化できるようになった.このことは,HMGAタンパク質が神経幹細胞の分化運命を若返らせることができる「リプログラミング因子」であることを示唆している.
また,ニューロン分化期からアストロサイト分化期にかけて神経幹細胞はその増殖速度が徐々に低下していくが,HMGAタンパク質はこの増殖低下にも重要な役割を果たす.すなわち,HMGA2をノックアウトしたり,HMGA1とHMGA2をノックダウンしたりすると,ニューロスフェアと呼ばれる一つの神経幹細胞が構築するコロニーの形成能が減少した51, 52).逆に,HMGAタンパク質の過剰発現によりニューロスフェア形成能が増加したことから,HMGAタンパク質は神経幹細胞の増殖にも寄与することがわかった.以上の結果から,HMGAタンパク質はニューロン分化期からアストロサイト分化期において転換する神経幹細胞の性質の多くを制御していることがわかった.
一方最近の研究で,HMGAタンパク質は増殖期からニューロン分化期に移行する際のニューロン分化能の獲得にも重要な役割を果たすことが明らかになった53).増殖期からニューロン分化期にかけて神経幹細胞は,ニューロン分化能の獲得に加えて対称分裂から非対称分裂への分裂様式の変化,放射状突起の伸展など,さまざまな変化を遂げる.しかし,胎仔がきわめて小さく,初代培養が難しく,大脳への遺伝子導入法が確立していないなどの理由により,この分化運命転換の分子メカニズムはほとんど不明であった.この報告では,増殖期神経幹細胞への新規遺伝子導入法を開発し,それを用いてHMGA2の機能解析を行った.というのも,胎生8日目から胎生12日目まで大脳あるいは大脳新皮質の神経幹細胞を採取しRNA-seqを行ったところ,HMGA2の発現が増殖期からニューロン分化期への転換点である胎生10日目で一過的に増加したからである.新規遺伝子導入法を用いて胎生8日目にHMGA2のノックダウンウイルスを導入したところ,胎生11日目において早期ニューロンマーカーTbr2陽性細胞が減少することがわかった.また,胎生10日目に遺伝子導入された細胞を回収しRNA-seq解析を行ったところ,遺伝子発現パターンが増殖期のそれに近くなることがわかった.これらの結果から,HMGA2は大脳新皮質神経幹細胞がニューロン分化を開始するために重要な遺伝子であることがわかった.
これらの報告からHMGAタンパク質は大脳新皮質神経幹細胞のニューロン分化期の始まりと終わりを決定する因子であることがわかった.ではHMGAタンパク質はどのようにクロマチン構造を制御して神経幹細胞にニューロン分化能を賦与しているのだろうか? これまでの研究からHMGAタンパク質は以下の三つのメカニズムで神経幹細胞のニューロン分化能を制御していると考えられる.
まず一つは,HMGAタンパク質がIFN-βやIL-2Rα, β-globinのように特定の下流遺伝子の発現を制御することで,神経幹細胞にニューロン分化能を賦与している可能性である.実際に,神経幹細胞においてHMGA2がIgf2bp2やPlag1という遺伝子の転写を亢進すること,またIgf2bp2やPlag1は神経幹細胞のニューロン分化能を制御する重要な分子であることが明らかとなっている54, 55).両遺伝子座にはHMGA2が直接結合することも,神経幹細胞を用いたChIP解析により明らかになっている(岸ら,未発表)55).しかし,HMGA2が神経幹細胞にてこれらの遺伝子の転写をどのように亢進しているかは不明である.過去の報告から,HMGA2は造血系や筋肉,がん細胞などさまざまな細胞でIgf2bp2を制御していることがわかっているが,ある報告ではHMGA2はNF-κBと協調的にIgf2bp2遺伝子座の活性を亢進することが明らかとなっている56–58).NF-κBはIFN-βやIL-2α遺伝子座においてHMGA1とエンハンソソームを形成するために重要な因子であったことから,Igf2bp2遺伝子座においてもHMGA2はエンハンソソームのような複合体形成を介して転写を亢進している可能性がある.
二つ目のメカニズムは,HMGAタンパク質が神経幹細胞のクロマチン構造をグローバルにオープンにしている可能性である.多能性を持つマウスES細胞は,クロマチンの構造が核全体でオープンになっており,そのことがその後分化する可能性のあるさまざまな系譜の遺伝子を転写可能な状態にし,多能性を保持していると考えられている59–61).神経幹細胞でも,DNase感受性試験やFRAP実験によりニューロン分化期ではアストロサイト分化期よりもクロマチン構造がグローバルにオープンであることが示されている52).そして,HMGAタンパク質のノックダウン,過剰発現によりこのクロマチン構造が変化することから,HMGAタンパク質がこのグローバルなクロマチン構造の制御を行っていると考えられる.では,HMGAタンパク質はクロマチン構造をどのようにしてオープンにしているのだろうか? HMGAタンパク質はヒストンH1と競合する,というこれまでの報告を考えるならば,HMGAタンパク質はヒストンH1をクロマチン上から剥がすことで,クロマチンをオープンにしている可能性がある32).しかし,最近のES細胞でのHMGA2の分布を調べた報告によると,ゲノムワイドにはHMGA2はヘテロクロマチンドメインに局在することから,単純にヒストンH1を剥がしてヘテロクロマチンをユークロマチン化しているわけではないようにも考えられる19).HMGAタンパク質がどのようにグローバルなクロマチン構造変換に関わっているかについては,試験管内の生化学実験と生体内神経幹細胞による実験を組み合わせた今後の解析が必要になってくるだろう.
最後に,HMGAタンパク質はポリコーム群タンパク質(Polycomb group proteins:PcG)の活性を制御している可能性が考えられる.PcGは,H3K27me3を触媒するポリコーム抑制複合体2(Polycomb repressive complex 2:PRC2)複合体と,それを認識してH2AK119ubを触媒するPRC1により構成される抑制性エピジェネティック因子である62, 63).これまでの研究により,神経幹細胞においてPcGはニューロン分化に関わる遺伝子を抑制することで,ニューロン分化期からアストロサイト分化期への移行を促進していることがわかっている64–68).これらの報告で同定されたPcGの標的遺伝子は,増殖期にHMGA2をノックダウンすると転写が活性化していることが,RNA-seqの結果から明らかとなった53).では,どのようにしてHMGA2はPcGの活性を抑制しているのだろうか? 現段階では,HMGA2が直接クロマチン上でPcGの活性を抑制している可能性も,何らかの遺伝子の発現調節を介して間接的にPcGの活性を抑制している可能性もいずれもありうる.ES細胞や神経幹細胞にてHMGA2の分布を調べた実験においてHMGA2とH3K27me3の分布が排反になることは,HMGA2がPcGの活性をクロマチン上で制御している可能性を示唆するが,その詳細について今後の解析が期待される19, 53).
引用文献References
1) Reeves, R. (2001) Molecular biology of HMGA proteins: hubs of nuclear function. Gene, 277, 63–81.
2) Goodwin, G.H., Sanders, C., & Johns, E.W. (1973) A new group of chromatin-associated proteins with a high content of acidic and basic amino acids. Eur. J. Biochem., 38, 14–19.
3) Solomon, M.J., Strauss, F., & Varshavsky, A. (1986) A mammalian high mobility group protein recognizes any stretch of six A.T base pairs in duplex DNA. Proc. Natl. Acad. Sci. USA, 83, 1276–1280.
4) Reeves, R. & Nissen, M.S. (1990) The A.T-DNA-binding domain of mammalian high mobility group I chromosomal proteins. A novel peptide motif for recognizing DNA structure. J. Biol. Chem., 265, 8573–8582.
5) Zhou, X., Benson, K.F., Ashar, H.R., & Chada, K. (1995) Mutation responsible for the mouse pygmy phenotype in the developmentally regulated factor HMGI-C. Nature, 376, 771–774.
6) Federico, A., Forzati, F., Esposito, F., Arra, C., Palma, G., Barbieri, A., Palmieri, D., Fedele, M., Pierantoni, G.M., De Martino, I., et al. (2014) Hmga1/Hmga2 double knock-out mice display a “superpygmy” phenotype. Biol. Open, 3, 372–378.
7) Fusco, A. & Fedele, M. (2007) Roles of HMGA proteins in cancer. Nat. Rev. Cancer, 7, 899–910.
8) Vignali, R. & Marracci, S. (2020) HMGA Genes and Proteins in Development and Evolution. Ijms, 21, 654–39.
9) Geierstanger, B.H., Volkman, B.F., Kremer, W., & Wemmer, D.E. (1994) Short peptide fragments derived from HMG-I/Y proteins bind specifically to the minor groove of DNA. Biochemistry, 33, 5347–5355.
10) Fonfría-Subirós, E., Acosta-Reyes, F., Saperas, N., Pous, J., Subirana, J.A., & Campos, J.L. (2012) Crystal structure of a complex of DNA with one AT-hook of HMGA1. PLoS One, 7, e37120.
11) Noro, B., Licheri, B., Sgarra, R., Rustighi, A., Tessari, M.A., Chau, K.-Y., Ono, S.J., Giancotti, V., & Manfioletti, G. (2003) Molecular dissection of the architectural transcription factor HMGA2. Biochemistry, 42, 4569–4577.
12) Frost, L., Baez, M.A.M., Harrilal, C., Garabedian, A., Fernandez-Lima, F., & Leng, F. (2015) The Dimerization State of the Mammalian High Mobility Group Protein AT-Hook 2 (HMGA2). PLoS One, 10, e0130478.
13) Sgarra, R., Zammitti, S., Lo Sardo, A., Maurizio, E., Arnoldo, L., Pegoraro, S., Giancotti, V., & Manfioletti, G. (2010) HMGA molecular network: From transcriptional regulation to chromatin remodeling. Biochim. Biophys. Acta, 1799, 37–47.
14) Reeves, R. & Nissen, M.S. (1993) Interaction of high mobility group-I (Y) nonhistone proteins with nucleosome core particles. J. Biol. Chem., 268, 21137–21146.
15) Reeves, R. & Wolffe, A.P. (1996) Substrate structure influences binding of the non-histone protein HMG-I(Y) to free nucleosomal DNA. Biochemistry, 35, 5063–5074.
16) Disney, J.E., Johnson, K.R., Magnuson, N.S., Sylvester, S.R., & Reeves, R. (1989) High-mobility group protein HMG-I localizes to G/Q- and C-bands of human and mouse chromosomes. J. Cell Biol., 109, 1975–1982.
17) Saitoh, Y. & Laemmli, U.K. (1994) Metaphase chromosome structure: bands arise from a differential folding path of the highly AT-rich scaffold. Cell, 76, 609–622.
18) Laemmli, U.K., Käs, E., Poljak, L., & Adachi, Y. (1992) Scaffold-associated regions: cis-acting determinants of chromatin structural loops and functional domains. Curr. Opin. Genet. Dev., 2, 275–285.
19) Colombo, D.F., Burger, L., Baubec, T., & Schübeler, D. (2017) Binding of high mobility group A proteins to the mammalian genome occurs as a function of AT-content. PLoS Genet., 13, e1007102.
20) John, S., Reeves, R.B., Lin, J.X., Child, R., Leiden, J.M., Thompson, C.B., & Leonard, W.J. (1995) Regulation of cell-type-specific interleukin-2 receptor alpha-chain gene expression: potential role of physical interactions between Elf-1, HMG-I(Y), and NF-kappa B family proteins. Mol. Cell. Biol., 15, 1786–1796.
21) John, S., Robbins, C.M., & Leonard, W.J. (1996) An IL-2 response element in the human IL-2 receptor alpha chain promoter is a composite element that binds Stat5, Elf-1, HMG-I(Y) and a GATA family protein. EMBO J., 15, 5627–5635.
22) Reeves, R., Leonard, W.J., & Nissen, M.S. (2000) Binding of HMG-I(Y) imparts architectural specificity to a positioned nucleosome on the promoter of the human interleukin-2 receptor alpha gene. Mol. Cell. Biol., 20, 4666–4679.
23) Thanos, D. & Maniatis, T. (1992) The high mobility group protein HMG I(Y) is required for NF-kappa B-dependent virus induction of the human IFN-beta gene. Cell, 71, 777–789.
24) Thanos, D. & Maniatis, T. (1995) Virus induction of human IFN beta gene expression requires the assembly of an enhanceosome. Cell, 83, 1091–1100.
25) Yie, J., Liang, S., Merika, M., & Thanos, D. (1997) Intra- and intermolecular cooperative binding of high-mobility-group protein I(Y) to the beta-interferon promoter. Mol. Cell. Biol., 17, 3649–3662.
26) Munshi, N., Merika, M., Yie, J., Senger, K., Chen, G., & Thanos, D. (1998) Acetylation of HMG I(Y) by CBP turns off IFN beta expression by disrupting the enhanceosome. Mol. Cell, 2, 457–467.
27) Munshi, N., Agalioti, T., Lomvardas, S., Merika, M., Chen, G., & Thanos, D. (2001) Coordination of a transcriptional switch by HMGI(Y) acetylation. Science, 293, 1133–1136.
28) Vernimmen, D. & Bickmore, W.A. (2015) The Hierarchy of Transcriptional Activation: From Enhancer to Promoter. Trends Genet., 31, 696–708.
29) Fong, T.C. & Emerson, B.M. (1992) The erythroid-specific protein cGATA-1 mediates distal enhancer activity through a specialized beta-globin TATA box. Genes Dev., 6, 521–532.
30) Bagga, R., Michalowski, S., Sabnis, R., Griffith, J.D., & Emerson, B.M. (2000) HMG I/Y regulates long-range enhancer-dependent transcription on DNA and chromatin by changes in DNA topology. Nucleic Acids Res., 28, 2541–2550.
31) Happel, N. & Doenecke, D. (2009) Histone H1 and its isoforms: contribution to chromatin structure and function. Gene, 431, 1–12.
32) Zhao, K., Käs, E., Gonzalez, E., & Laemmli, U.K. (1993) SAR-dependent mobilization of histone H1 by HMG-I/Y in vitro: HMG-I/Y is enriched in H1-depleted chromatin. EMBO J., 12, 3237–3247.
33) Catez, F., Yang, H., Tracey, K.J., Reeves, R., Misteli, T., & Bustin, M. (2004) Network of dynamic interactions between histone H1 and high-mobility-group proteins in chromatin. Mol. Cell. Biol., 24, 4321–4328.
34) Klein-Hessling, S., Schneider, G., Heinfling, A., Chuvpilo, S., & Serfling, E. (1996) HMG I(Y) interferes with the DNA binding of NF-AT factors and the induction of the interleukin 4 promoter in T cells. Proc. Natl. Acad. Sci. USA, 93, 15311–15316.
35) Arlotta, P., Rustighi, A., Mantovani, F., Manfioletti, G., Giancotti, V., Tell, G., & Damante, G. (1997) High mobility group I proteins interfere with the homeodomains binding to DNA. J. Biol. Chem., 272, 29904–29910.
36) Campisi, J. (2013) Aging, cellular senescence, and cancer. Annu. Rev. Physiol., 75, 685–705.
37) Narita, M., Nuñez, S., Heard, E., Narita, M., Lin, A.W., Hearn, S.A., Spector, D.L., Hannon, G.J., & Lowe, S.W. (2003) Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell, 113, 703–716.
38) Narita, M., Narita, M., Krizhanovsky, V., Nuñez, S., Chicas, A., Hearn, S.A., Myers, M.P., & Lowe, S.W. (2006) A novel role for high-mobility group a proteins in cellular senescence and heterochromatin formation. Cell, 126, 503–514.
39) Chandra, T., Kirschner, K., Thuret, J.-Y., Pope, B.D., Ryba, T., Newman, S., Ahmed, K., Samarajiwa, S.A., Salama, R., Carroll, T., et al. (2012) Independence of Repressive Histone Marks and Chromatin Compaction during Senescent Heterochromatic Layer Formation. Mol. Cell, 47, 203–214.
40) Sadaie, M., Salama, R., Carroll, T., Tomimatsu, K., Chandra, T., Young, A.R.J., Narita, M., Pérez-Mancera, P.A., Bennett, D.C., Chong, H., et al. (2013) Redistribution of the Lamin B1 genomic binding profile affects rearrangement of heterochromatic domains and SAHF formation during senescence. Genes Dev., 27, 1800–1808.
41) Sati, S., Bonev, B., Szabo, Q., Jost, D., Bensadoun, P., Serra, F., Loubiere, V., Papadopoulos, G.L., Rivera-Mulia, J.C., Fritsch, L., et al. (2020) 4D Genome Rewiring during Oncogene-Induced and Replicative Senescence. Mol. Cell, 78, 522–538.e9.
42) Funayama, R., Saito, M., Tanobe, H., & Ishikawa, F. (2006) Loss of linker histone H1 in cellular senescence. J. Cell Biol., 175, 869–880.
43) Otz, M.G. & Huttner, W.B. (2005) The cell biology of neurogenesis. Nat. Rev. Mol. Cell Biol., 6, 777–788.
44) Hirabayashi, Y. & Gotoh, Y. (2010) Epigenetic control of neural precursor cell fate during development. Nat. Rev. Neurosci., 11, 377–388.
45) Kriegstein, A. & Alvarez-Buylla, A. (2009) The Glial Nature of Embryonic and Adult Neural Stem Cells. Annu. Rev. Neurosci., 32, 149–184.
46) Mukhtar, T. & Taylor, V. (2018) Untangling Cortical Complexity During Development. J. Exp. Neurosci., 12, 1179069518759332.
47) Tsuboi, M., Hirabayashi, Y., & Gotoh, Y. (2019) Diverse gene regulatory mechanisms mediated by Polycomb group proteins during neural development. Curr. Opin. Neurobiol., 59, 164–173.
48) Kishi, Y. & Gotoh, Y. (2018) Regulation of Chromatin Structure During Neural Development. Front. Neurosci., 12, 874.
49) Nakagawa, T., Wada, Y., Katada, S., & Kishi, Y. (2020) Epigenetic regulation for acquiring glial identity by neural stem cells during cortical development. Glia, 14, 67–14.
50) Sanosaka, T., Namihira, M., Asano, H., Kohyama, J., Aisaki, K., Igarashi, K., Kanno, J., & Nakashima, K. (2008) Identification of genes that restrict astrocyte differentiation of midgestational neural precursor cells. Neuroscience, 155, 780–788.
51) Nishino, J., Kim, I., Chada, K., & Morrison, S.J. (2008) Hmga2 promotes neural stem cell self-renewal in young but not old mice by reducing p16Ink4a and p19Arf Expression. Cell, 135, 227–239.
52) Kishi, Y., Fujii, Y., Hirabayashi, Y., & Gotoh, Y. (2012) HMGA regulates the global chromatin state and neurogenic potential in neocortical precursor cells. Nat. Neurosci., 15, 1127–1133.
53) Kuwayama, N., Kishi, Y., Maeda, Y., Nishiumi, Y., Suzuki, Y., Koseki, H., Hirabayashi, Y., & Gotoh, Y. (2020) In utero gene transfer system for embryos before neural tube closure reveals a role for Hmga2 in the onset of neurogenesis. bioRxiv., 125, 2020.05.14.086330.
54) Fujii, Y., Kishi, Y., & Gotoh, Y. (2013) IMP2 regulates differentiation potentials of mouse neocortical neural precursor cells. Genes Cells, 18, 79–89.
55) Sakai, H., Fujii, Y., Kuwayama, N., Kawaji, K., Gotoh, Y., & Kishi, Y. (2019) Plag1 regulates neuronal gene expression and neuronal differentiation of neocortical neural progenitor cells. Genes Cells, 24, 650–666.
56) Cleynen, I., Brants, J.R., Peeters, K., Deckers, R., Debiec-Rychter, M., Sciot, R., Van de Ven, W.J., & Petit, M.M. (2007) HMGA2 regulates transcription of the Imp2 gene via an intronic regulatory element in cooperation with nuclear factor-kappaB. Mol. Cancer Res., 5, 363–372.
57) Li, Z., Gilbert, J.A., Zhang, Y., Zhang, M., Qiu, Q., Ramanujan, K., Shavlakadze, T., Eash, J.K., Scaramozza, A., Goddeeris, M.M., et al. (2012) An HMGA2-IGF2BP2 axis regulates myoblast proliferation and myogenesis. Dev. Cell, 23, 1176–1188.
58) Copley, M.R., Babovic, S., Benz, C., Knapp, D.J.H.F., Beer, P.A., Kent, D.G., Wohrer, S., Treloar, D.Q., Day, C., Rowe, K., et al. (2013) The Lin28b-let-7-Hmga2 axis determines the higher self-renewal potential of fetal haematopoietic stem cells. Nat. Cell Biol., 15, 916–925.
59) Meshorer, E., Yellajoshula, D., George, E., Scambler, P.J., Brown, D.T., & Misteli, T. (2006) Hyperdynamic plasticity of chromatin proteins in pluripotent embryonic stem cells. Dev. Cell, 10, 105–116.
60) Meshorer, E. & Misteli, T. (2006) Chromatin in pluripotent embryonic stem cells and differentiation. Nat. Rev. Mol. Cell Biol., 7, 540–546.
61) Gaspar-Maia, A., Alajem, A., Polesso, F., Sridharan, R., Mason, M.J., Heidersbach, A., Ramalho-Santos, J., McManus, M.T., Plath, K., Meshorer, E., et al. (2009) Chd1 regulates open chromatin and pluripotency of embryonic stem cells. Nature, 460, 863–868.
62) Margueron, R. & Reinberg, D. (2011) The Polycomb complex PRC2 and its mark in life. Nature, 469, 343–349.
63) Di Croce, L. & Helin, K. (2013) Transcriptional regulation by Polycomb group proteins. Nat. Struct. Mol. Biol., 20, 1147–1155.
64) Hirabayashi, Y., Suzki, N., Tsuboi, M., Endo, T.A., Toyoda, T., Shinga, J., Koseki, H., Vidal, M., & Gotoh, Y. (2009) Polycomb limits the neurogenic competence of neural precursor cells to promote astrogenic fate transition. Neuron, 63, 600–613.
65) Pereira, J.D., Sansom, S.N., Smith, J., Dobenecker, M.-W., Tarakhovsky, A., & Livesey, F.J. (2010) Ezh2, the histone methyltransferase of PRC2, regulates the balance between self-renewal and differentiation in the cerebral cortex. Proc. Natl. Acad. Sci. USA, 107, 15957–15962.
66) Morimoto-Suzki, N., Hirabayashi, Y., Tyssowski, K., Shinga, J., Vidal, M., Koseki, H., & Gotoh, Y. (2014) The polycomb component Ring1B regulates the timed termination of subcerebral projection neuron production during mouse neocortical development. Development, 141, 4343–4353.
67) Albert, M., Kalebic, N., Florio, M., Lakshmanaperumal, N., Haffner, C., Brandl, H., Henry, I., & Huttner, W.B. (2017) Epigenome profiling and editing of neocortical progenitor cells during development. EMBO J., 36, 2642–2658.
68) Tsuboi, M., Kishi, Y., Yokozeki, W., Koseki, H., Hirabayashi, Y., & Gotoh, Y. (2018) Ubiquitination-Independent Repression of PRC1 Targets during Neuronal Fate Restriction in the Developing Mouse Neocortex. Dev. Cell, 47, 758–772.e5.
69) Lieberman-Aiden, E., van Berkum, N.L., Williams, L., Imakaev, M., Ragoczy, T., Telling, A., Amit, I., Lajoie, B.R., Sabo, P.J., Dorschner, M.O., et al. (2009) Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science, 326, 289–293.
70) Rao, S.S.P., Huntley, M.H., Durand, N.C., Stamenova, E.K., Bochkov, I.D., Robinson, J.T., Sanborn, A.L., Machol, I., Omer, A.D., Lander, E.S., et al. (2014) A 3D Map of the Human Genome at Kilobase Resolution Reveals Principles of Chromatin Looping. Cell, 159, 1665–1680.
71) Buenrostro, J.D., Wu, B., Litzenburger, U.M., Ruff, D., Gonzales, M.L., Snyder, M.P., Chang, H.Y., & Greenleaf, W.J. (2015) Single-cell chromatin accessibility reveals principles of regulatory variation. Nature, 523, 486–490.
72) Hsieh, T.-H.S., Fudenberg, G., Goloborodko, A., & Rando, O.J. (2016) Micro-C XL: assaying chromosome conformation from the nucleosome to the entire genome. Nat. Methods, 13, 1009–1011.
73) Maeshima, K., Tamura, S., Hansen, J.C., & Itoh, Y. (2020) Fluid-like chromatin: Toward understanding the real chromatin organization present in the cell. Curr. Opin. Cell Biol., 64, 77–89.