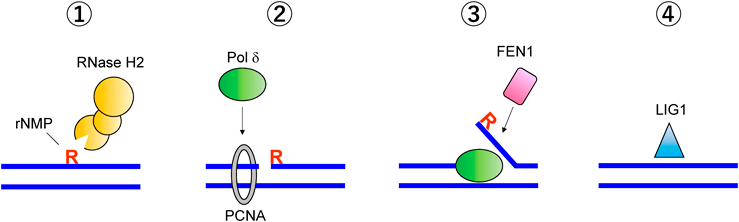
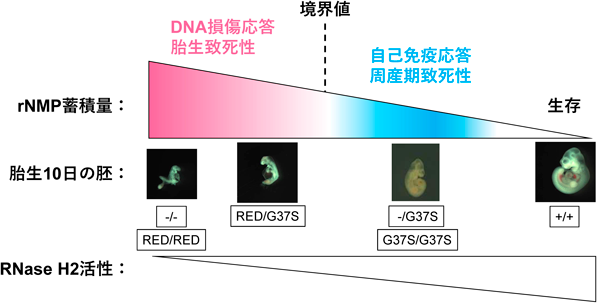
染色体DNAにおけるリボヌクレオチドの許容限界A threshold of ribonucleotide tolerance in chromosomal DNA
愛知県がんセンター研究所腫瘍制御学分野Division of Cancer Cell Regulation, Aichi Cancer Center Research Institute ◇ 愛知県名古屋市千種区鹿子殿1番1号 ◇ 1–1 Kanokoden, Chikusa-ku, Nagoya
発行日:2020年10月25日Published: October 25, 2020