動物では,cAMPやcGMPといった環状ヌクレオチドがさまざまな生命機能に関与することが知られている.細菌でもこれらの因子が機能しているが,最近bis(3′,5′)-cyclic diguanylic acid(環状ジグアニル酸,以下c-di-GMP)は,細菌が環境に適応する際のさまざまな反応を制御するアロステリックなアクチベーターであり,セカンドメッセンジャーとしても働くことがわかってきた.c-di-GMPは,細菌界で広く作られ,また細菌界に特有のものである.c-di-GMPはセルロースを合成することで知られる細菌,Gluconacetobacter xylinusにおいてセルロース合成酵素を正に調節する因子として見つかり,膜結合型のセルロース合成系のアロステリック活性因子に結合することが明らかにされた1–3).その後,細胞外成分であるべん毛あるいは線毛の合成と機能調節,あるいは,菌体外多糖合成を制御することが明らかにされた4).c-di-GMPは,いろいろな結合タンパク質を介して,多様な機能を発現する.その結合ドメインとして,PilZドメインが知られている.PilZタンパク質ファミリーは,緑膿菌で初めて同定されたIV型線毛(ピリ)制御タンパク質PilZにちなんで命名された.PilZは,ピリ産生オペロンの一部として発現する13 kDaの小さな細胞質タンパク質であり,ピリの生合成に必須である.ゲノム解析からPilZドメインがc-di-GMPと結合する主な部位なのではないかと予測された5).精製されたPilZドメインを持つタンパク質を用いて実際にその結合が証明され6),c-di-GMPを結合したPilZドメインの結晶構造も報告された7).
細菌におけるセカンドメッセンジャーとしては,cAMP, cGMP, ppGpp,そしてc-di-GMP, c-di-AMPなどが知られている8, 9).この中では,cAMPが最も早くに発見され研究された.大腸菌では糖代謝の転写制御に働いていることが明らかにされたが,動物ではGタンパク質共役受容体と共役して非常に多様な制御に関与していることがわかっている10).細菌においてはc-di-GMPは,非常に多様な環境応答に対応している.
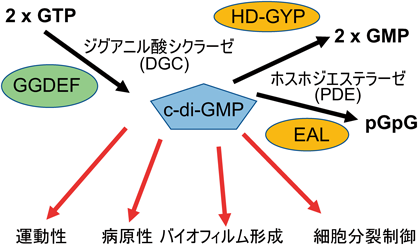
c-di-GMPは,ジグアニル酸シクラーゼ(diguanylate cyclase:DGC)によってGTP 2分子から合成され,ホスホジエステラーゼ(phosphodiesterase:PDE)によってGMP 2分子あるいは直鎖diGMP(pGpG)に分解される.シクラーゼ活性は,GGDEFタンパク質ドメイン(DUF1としても知られている)にコードされている11, 12).DGCのGGDEFドメインは,Caulobacter crescentusにおいてべん毛の形成を制御するPleDが持つ5個のアミノ酸残基GGDEFから命名された13).その後,このモチーフがDGCの活性中心を形成していることが明らかにされた14).また,ホスホジエステラーゼ活性は,EAL(DUF2)ドメイン15–17)およびHD-GYPドメイン18)にコードされている(図1).
多様な環境応答に対応するため,c-di-GMPの合成と分解を担うと考えられるGGDEF, EAL, HD-GYPドメインを含むタンパク質が細菌のゲノム上に非常に多くコードされていることが,ゲノム解析からわかっている19).Vibrio choleraeではゲノム配列から推定されるc-di-GMPの合成と分解に関わるタンパク質として,GGDEFが31, GGDEFとEALを両方持つものが10, EALが12, HD-GYPが9,合わせて62種類もの因子が存在している.表1に示すように,Vibrio属菌の持つ数は大腸菌に比べて多い.一方,古細菌ではこれらのホモログは存在していない.なぜこのように多くのc-di-GMP合成分解酵素が必要なのかはよくわかっていないが,厳しい環境に対応するために進化したことは想像にかたくない.
表1 γプロテオバクテリアのc-di-GMP合成分解タンパク質とエフェクタータンパク質の数| 菌種 | タンパク質 | GGDEF | GGDEF+EAL | EAL | HD-GYP | PilZ | MshEN | その他 |
|---|
| 1:E. coli | 4132 | 12 | 7 | 10 | — | 2 | 1 | BcsE |
| 2:P. aeru | 5567 | 17 | 16 | 5 | 3 | 8 | 2 | FleQ |
| 3:V. chol | 3835 | 31 | 10 | 12 | 9 | 5 | 1 | VpsT, FleQ |
| 4:V. harv | 6055 | 32 | 12 | 14 | 5 | 5 | 1 | VpsT, FleQ |
| 5:V. para | 4832 | 28 | 16 | 13 | 5 | 5 | 1 | VpsT, FleQ |
| 6:V. anti | 4518 | 29 | 16 | 11 | 4 | 5 | 1 | VpsT, FleQ |
| この表は文献19で発表されたWeb補足データより引用した(https://www.ncbi.nlm.nih.gov/Complete_Genomes/c-di-GMP.html).1:Escherichia coli(大腸菌),2:Pseudomonas aeruginosa(緑膿菌),3:Vibrio cholerae(コレラ菌),4:Vibrio harveyi,5:Vibrio parahaemolyticus(腸炎ビブリオ),6:Vibrio antiquarius. |
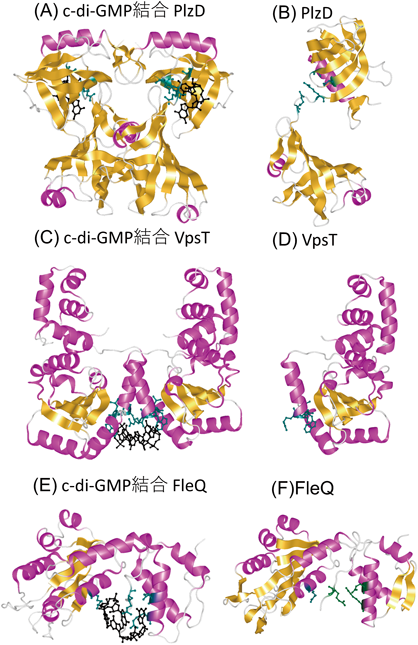
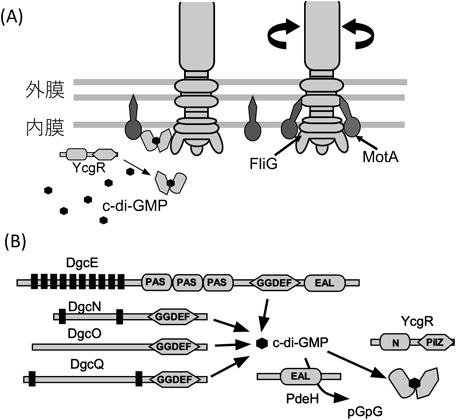
最初に決められたc-di-GMP結合タンパク質は,Vibrio choleraeのPlzDである7).PlzD(PilZ domain-containing protein D)はもともとVCA0042タンパク質として,V. choleraeゲノム配列から同定された五つのPilZドメインタンパク質の一つである.このタンパク質のN末端側は,大腸菌においてべん毛遺伝子制御に関わるYcgRと相同なドメイン(YcgR-N)を持つ.YcgRはそのC末端側にPilZドメインを持ち,後にc-di-GMPを結合してべん毛モーターに作用することが明らかにされたが,当初はH-NSと呼ばれる核様体タンパク質の欠損株によるべん毛形成を抑圧する遺伝子として同定されていた20).PlzDには,C末端側のPilZドメイン部分にRxxxRとD/NxSxxGのc-di-GMP結合コンセンサス配列が存在する.PlzDについてc-di-GMPのないアポ構造だけでなく,c-di-GMPをPilZドメインに1分子結合した複合体構造も解かれ,c-di-GMP結合型PlzDは二量体を形成していることが明らかにされた7).重要なことに,c-di-GMP結合によってPlzDの大きな構造変化が生じていた(図2A, B).大腸菌YcgRにおいては,C末端のPilZドメインがべん毛モーターの固定子タンパク質MotAと相互作用し,N末端ドメインが他のモータータンパク質(回転子タンパク質のFliGを想定)と相互作用して,運動を阻害すると現状では考えられている21).最近,ClpXPプロテアーゼによる転写因子の分解がc-di-GMP依存的に起こり,べん毛遺伝子制御系に影響することがわかってきた22).YcgRが直接にべん毛遺伝子制御に関わっているかはわかっていない.
Vibrio属菌の中で,c-di-GMP結合タンパク質として構造が解けているものにVpsTがある23).このタンパク質は,5節で述べるようにLuxRやCsgDファミリーに属し,リン酸を受ける二成分制御系のエフェクタータンパク質である.推定リン酸化部位は保存されているものの,他のリン酸転移に関与する残基は保存されていない.N末端がレシーバー(REC)ドメインで,C末端がヘリックス・ターン・ヘリックス(helix-turn-helix:HTH)モチーフを持つ転写制御部位である.4残基(W[F/L/M][T/S]R)のc-di-GMP結合保存配列を持つ.c-di-GMPとVpsTの結合モル比は1:1で,2分子のc-di-GMPが二量体のVpsTに結合している.c-di-GMP 2分子を挟むようにVpsTが配置している.ただし,VpsTの二量体化にはc-di-GMPは必要ないらしい.c-di-GMPが結合することで,DNA結合部位のHTHドメインの構造が変わることでDNAとの相互作用が変わって,転写活性に影響するということである(図2C, D).
最後にもう一つc-di-GMP結合タンパク質の構造を紹介したい.Vibrio属菌のものではないのだが,Pseudomonas aeruginosaのFleQタンパク質である.7節でも述べるように,FleQはV. choleraeのべん毛遺伝子マスターレギュレーターとして知られるFlrAのオルソログで,P. aeruginosaでべん毛遺伝子のマスターレギュレーターとして機能する.FleQはN末端にRECレシーバードメイン,中央に細菌AAA+ATPase/σ54相互作用ドメイン,C末端にHTH DNA結合ドメインを持ち,細菌のエンハンサー結合タンパク質(bEBP)であるNtrCサブファミリーと高い相同性を持つ.しかし,制御機構はNtrCとかなり違っていると推測される.FleQはc-di-GMPが結合していない状況では,安定な六量体リング構造を作って,転写を抑制していると考えられる.c-di-GMP濃度が上昇して結合すると,六量体構造が壊れて構造を変えることによって転写がONになると考えられる.c-di-GMPが結合する制御領域は,RECとAAA+ドメインのインターフェースに存在するLFR144Sモチーフ,post-WalkerA領域のR185とN186残基,そしてExxxR334配列であることが構造データから示されている(図2E, F)24).
4. Vibrio属菌でのc-di-GMPによる病原因子制御
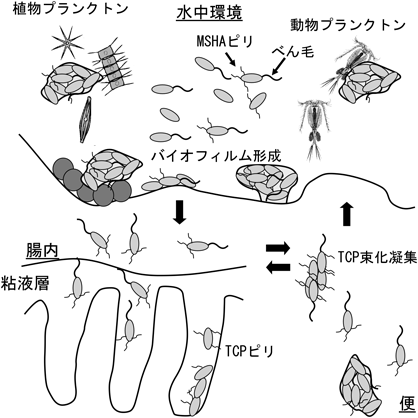
コレラの病原細菌であるV. choleraeのライフサイクルは,free-swimming phaseとvirulent phaseの二つに大別される.前者は,主に宿主外に存在しているときの状態であり,病原因子を発現することはなく,活発に動き回る.実際の環境では,菌はべん毛を使って水中を活発に運動する遊泳形態か,魚やエビあるいは,プランクトンや堆積物などの表面にバイオフィルムを形成して存在する固着形態をとって生育する(図3)25).水中環境において付着してバイオフィルムを形成するときには,IV型線毛であるマンノース感受性ヘマグルチニン(mannose-sensitive hemagglutinin:MSHA)線毛を形成する.宿主内に入り,感染する腸の表面に到達すると後者の状態に移り,運動性がなくなると同時に,電解質の分泌(漏出)を引き起こすコレラ毒素(cholera toxin:CT)に代表される病原因子と小腸のコロニー化に必要とされる毒素共調節線毛(toxin co-regulated pilus:TCP)を発現する(図4).これが一連のコレラの症状を引き起こす.
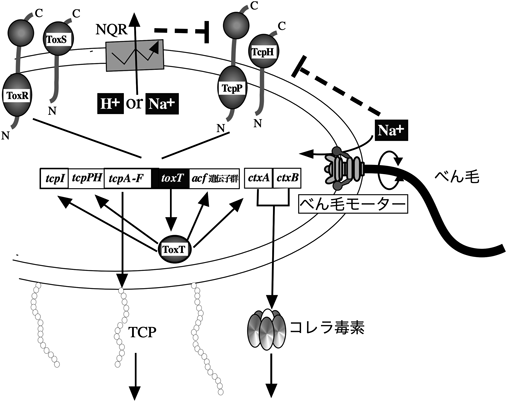
V. choleraeの病原性には,いくつかのべん毛遺伝子や,走化性関連遺伝子が病原性に関与することが知られている.AphA/B, TcpP/H, ToxR/SおよびToxTを含む転写因子のカスケードは,CTおよびTCPの発現を調節することが知られていた26).V. choleraeの病原因子の発現に大きく関わっているのがToxTである.転写アクチベーターであるToxTが,CTをコードしているctxABのプロモーターを活性化することで毒素産生が起こり,病原性が発揮される27, 28).toxT遺伝子は,ToxRやTcpP(ともに内膜タンパク質,細胞質領域にDNA結合ドメインを持つ)によりその発現が活性化される29).これら遺伝子の発現誘導は,温度,pH,ナトリウムイオン濃度などの外部環境の影響を受けることが知られている.ToxTはCTの他に,toxT自身やTCPの主成分をコードするtcpAをはじめとするtcp(toxin co-regulated pilus)遺伝子群や,acf(accessory colonizing factor)遺伝子群の転写も活性化する30, 31).CTやTCPなどの多くの病原性因子の発現は,転写制御因子のToxRとその下流にあるToxTによって協調的に制御されている(図4).運動性を失ったV. chorelaeについてはCT, TCP,ヘモリシン,バイオフィルムなどの病原因子の発現が上昇し,逆に運動性が上昇した変異株では病原因子の発現が低下することが見いだされた32).
これらのことから,べん毛発現と病原因子発現を共通にコントロールする因子の存在が想像された.そこで,細菌において,生理条件の変化に対応して,種々の遺伝子の転写制御を行う代替σ因子が考えられた.通常の生育に使われているσ因子は,その分子量からσ70と呼ばれる.代替σ因子の中でσ54と呼ばれるものは,窒素同化遺伝子,べん毛遺伝子,線毛遺伝子などの発現を制御していることがわかっていた33).V. chorelaeでσ54欠損株が作製され,病原性因子の発現が調べられた34).それによるとCTやTCPへの影響はなく,べん毛形成に欠損が起こり,グルタミン酸合成酵素の発現に影響した.さらに,幼若マウスのモデル系において,V. chorelaeのコロニー形成がσ54欠損株では起こらなくなった.このコロニー形成欠損は,べん毛欠損やグルタミン酸合成酵素量による間接的な影響ではないことが示唆された.しかし,この状況でもσ因子がどのようにして環境からのシグナルによって制御されているのかは不明であった.
以上の病原因子の発現を結びつける因子として,c-di-GMPが登場することになる.c-di-GMPが直接関与しているという事実はなかったのだが,Vibrio属菌で,GGDEFとEALドメインを持っているタンパク質がバイオフィルム形成に関与しているということがわかっていた35–37).このような背景の中で,Vibrio属菌の二成分制御系のコレラ毒素産生に関与しているVieAが,バイオフィルム形成に必要な菌体外多糖合成遺伝子の発現を負に制御していることが明らかにされた38).そして,VieAがEALドメインを持ち,その分解活性の制御によりc-di-GMPの濃度が調節されて,遺伝子発現の制御が起こっていることが明らかになった.
5. Vibrio属菌でのバイオフィルム形成とc-di-GMP
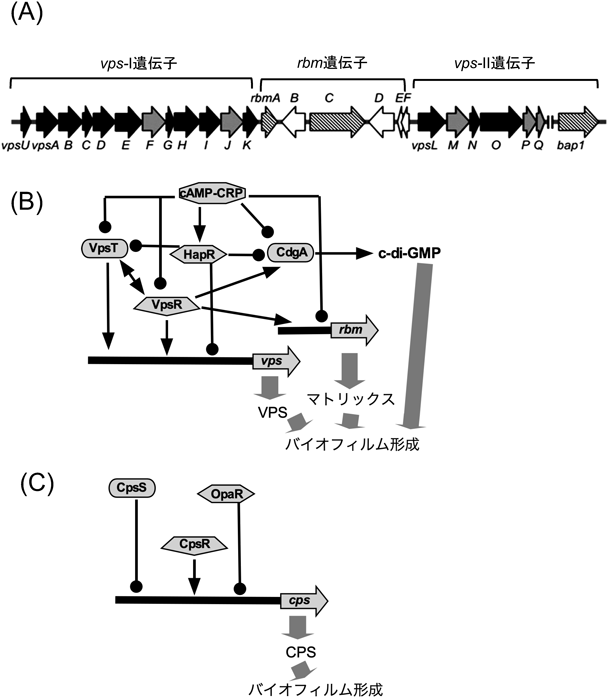
V. choleraeは,べん毛を使って泳ぐ遊泳状態から,ピリを使って固着状態になるとバイオフィルム形成を行う.バイオフィルムの主要構成因子は菌体外分泌多糖(Vibrio polysaccharide:VPS)であり,vps-I(vpsU, vpsA~K, VC0916~27)およびvps-II(vpsL~Q, VC0934~39)領域にVPS産生に必要な遺伝子がコードされている.これに加えて,マトリックスタンパク質(RbmA, RbmC,およびBap1)が加わってバイオフィルムとなる.マトリックスタンパク質合成に必要は遺伝子は,vps-Iとvps-II領域の間にクラスターとなって存在している(図5A)39).vps遺伝子発現は,VpsR, VpsT, HapRによって制御されている40).VpsRは,AAA+ATPaseドメインとDNA結合ドメインを含むσ54依存の転写調節促進因子である.VpsTはLuxRやCsgDファミリーに属する二成分制御系のリン酸化を受けるエフェクタータンパク質である.HapRはDNAに結合して転写を抑制するTetRファミリーに属する.VpsTはN末端レシーバー(REC)とDNA結合を媒介するC末端HTHドメインから構成されるタンパク質である.VpsTにc-di-GMPが結合することによって二量体化することで,上流のキナーゼによるリン酸化により,強い影響を転写に及ぼす23).VpsRがc-di-GMP依存的に発現制御を受けるTfoYという別の転写因子の発現制御を制御しているという複雑な制御機構もわかっている41).in vitro系を用いて,VpsRがσ70 RNAポリメラーゼとc-di-GMP依存的にvps遺伝子のプロモーターに作用して発現を制御していることが示されている42).マトリックスタンパク質の産生もVpsRによって制御を受けているのだが,さらにこれらが菌体外に分泌されるために必要なII型の分泌装置の遺伝子(eps遺伝子)発現をc-di-GMP依存的に制御している43).このeps遺伝子のプロモーター領域には,σEとσ70依存の二つのプロモーターが存在することが示されている44).さらに,c-di-GMPの濃度によってcAMPもバイオフィルム形成に関与し,非常に複雑な制御を受けている(図5B)40).腸炎ビブリオVibrio parahaemolyticusにおいても,バイオフィルムの主要構成因子はVPSであるが,合成に必要な遺伝子は1か所にまとまって存在し,cps(capsular polysaccharide)と名づけられている.その遺伝子組成もV. choleraeとは少し異なり,V. choleraeのVPSの組成が,グルコース(52.6%),ガラクトース(37.0%),N-アセチルグルコサミン(5.1%),マンノース(3.8%),キシロース(1.5%)に対して45),V. parahaemolyticusではグルコース(34.6%),ガラクトース(27.8%),フコース(21.3%),N-アセチルグルコサミン(13.9%),アラビノース(1.0%),マンノース(0.7%),N-アセチルガラクトサミン(0.6%)となっている46).遺伝子制御に関しても,V. choleraeとV. parahaemolyticusでは類似の転写制御因子(OpaR, CpsR, CpsS)が関与しているが,少し制御機構は異なっているらしい(図5C)47).
V. parahaemolyticusとその近縁種である海洋性ビブリオ菌Vibrio alginolyticusは,基本的には1本の極べん毛と多数の側べん毛という機能的に異なる2種類のべん毛を一つの菌体が持つ.V. choleraeでは,基本的には1本の極べん毛を,Vibrio fischeriは複数本の極べん毛を持つ.極べん毛と側べん毛のべん毛運動のエネルギー源を調べると,極べん毛はNa+駆動力,側べん毛はH+駆動力によって運動することがわかった48, 49).極べん毛は恒常的に発現しているのに対して,側べん毛は粘性の高い条件下で発現することから,Vibrio属菌の菌体は粘性の上昇を何らかの方法で感じていると考えられ,そのセンサーは極べん毛であるというダイナモメーター仮説が提案された50).そして,極べん毛のNa+駆動型モーターの阻害剤であるフェナミル(ナトリウムチャネル阻害剤:アミロライドの誘導体)を用いた実験から,極べん毛の回転の低下と側べん毛遺伝子の発現の間に相関関係があることが示された.このことから極べん毛は力学的センサーとして働き,側べん毛の発現を制御すると考えられている51).しかしながら,30年以上も前に発見された現象であるが,いまだにどのようにして極べん毛が粘性を感知しているかわかっていない.V. choleraeにおいて,極べん毛運動がバイオフィルム形成制御に影響することが報告されている.べん毛繊維タンパク質を欠損すると細胞のc-di-GMP濃度が上昇し,極べん毛のモーター機能に必須なMotYの欠損でその上昇が抑圧される52).極べん毛遺伝子がどのようにc-di-GMP濃度を制御しているかについては謎のままである.V. parahaemolyticusではScrABCが側べん毛の発現とバイオフィルム形成を制御していることが報告されている.ScrCは膜タンパク質であり,細胞質ドメインにGGDEF-EALドメインを持つためc-di-GMPの分解と合成活性を持ち,シグナルを受けてc-di-GMPの濃度を制御する53).極べん毛がScrCのc-di-GMP分解合成に関与するのかについてはわかっていない.
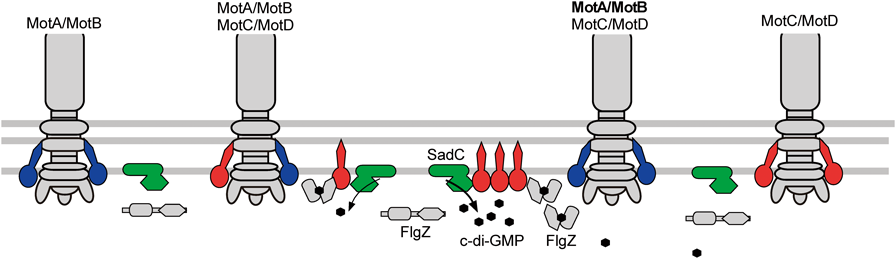
最近,べん毛系のc-di-GMP制御機構についてのヒントとなるような結果が緑膿菌で報告された54).Pseudomonas属菌は,べん毛の固定子タンパク質を2種類(MotA/MotBとMotC/MotD)持ち,MotC/MotD固定子により粘性の高い条件において,表面を這うように動く遊走運動をすることができる.この遊走運動に,PliZドメインを持ち大腸菌YcgRと相同性のあるFlgZが,MotCと相互作用することで影響することが示されていた55).MotC/MotDが欠損するとc-di-GMP濃度が低下し,MotC/MotDを過剰に発現するとc-di-GMP濃度が上昇することによりバイオフィルム形成の違いが起こることが発見された.また,C末端細胞質領域にGGDEFモチーフを持つ膜タンパク質SadCの膜貫通領域にMotCが作用することが見いだされ,MotC/MotD固定子複合体がDGCであるSadCに相互作用することでc-di-GMP合成活性が上昇するというモデルが発表された(図6)54).Vibrio属菌のPomA/PomB固定子複合体が,c-di-GMP分解合成活性のある膜タンパク質と相互作用して,c-di-GMP濃度が変化するということはありそうなことである.
べん毛による運動は,栄養豊富な培養条件では,菌の生育にまったく必要ない.研究室環境で,べん毛欠損株は野生株とまったく同様に生育する.しかし,自然環境の中では,生き残りに必要な形質である.そして,べん毛の組み立てコストは非常に大きなものである.赤痢菌などでは,べん毛遺伝子が不要になって失われたと考えられている.べん毛は20個以上の構造タンパク質から組み立てられ,それらのべん毛構造タンパク質をコードする遺伝子や発現を調節する遺伝子が50個以上存在する.それだけ大きなコストをかけても,細菌の生き残りには必要ということなのだ.コストを最小に抑えるため,必要のないときには,べん毛遺伝子発現は厳密に制御されている56, 57).
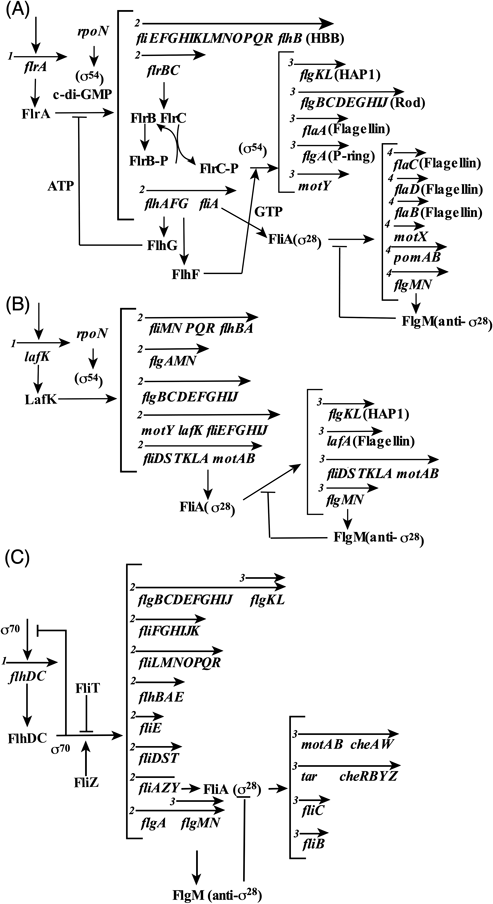
V. parahaemolyticusとV. choleraeの極べん毛遺伝子の発現制御が推測されている58–60).V. choleraeでは,オペロンの発現の順番により四つのクラスに分けられて,オペロンの転写制御のモデルが出されている(図7A)61, 62).pomAB(motAB)やmotXといったモーター遺伝子やフラジェリンをコードする遺伝子flaB, flaC, flaD, flaE,そして,推定アンチσ因子をコードするflgMを含むオペロンは,σ28型のプロモーター配列を持ち,fliA遺伝子がコードするσ28に依存して発現することが確認されている.これに対して,コアフラジェリンをコードするflaAやモータータンパク質をコードするmotY,フックやべん毛基部体の構造遺伝子とフック付随タンパク質(HAPs)などは,FlrCとσ54に依存した発現を示す.最初に転写されるのはflrAであり,その産物であるFlrAとσ54がクラスIIオペロンの転写を起こす.FlrBのリン酸基がFlrCに転移され,活性化される.先の節で述べたようにバイオフィルムを形成して定着するときには運動は不要になるのだから,このべん毛遺伝子発現は低下することになる.c-di-GMPがFlrAに結合することで,flrBCオペロンへのFlrAの結合が阻害されて,転写が起こらなくなりべん毛を形成しなくなることが報告されている63).このc-di-GMPによるべん毛遺伝子発現制御は緑膿菌でもみられており,FlrAのホモログであるFleQの機能メカニズムがよく研究されている.FleQはFlrAと約50%の相同性を持った緑膿菌の極べん毛遺伝子マスターレギュレーターで,AAA+タイプに分類されるATPaseである.FleQは六量体を形成し,c-di-GMPの結合によって,そのATPase活性が制御される64).FleQについては結晶構造も解かれており(図2E, F),c-di-GMPの結合によって六量体構造の再構築が起こることが示唆されている24).
V. parahaemolyticusの極べん毛遺伝子もV. choleraeと同様な(遺伝子名は異なるが)転写制御によって調節されることがわかっている65, 66).ビブリオ属の極べん毛制御は,同じ極べん毛を持つシュードモナス属のべん毛遺伝子の制御機構と類似している67).V. parahaemolyticusの側べん毛遺伝子の転写制御も解析され,階層的に制御されているていることがわかっている(図7B).面白いことに,周毛性のV. parahaemolyticusの側べん毛遺伝子のマスター制御遺伝子はσ54タイプであるが,下流は大腸菌のもつσ28型に近い制御因子が働いている(図7C)68).
8. c-di-GMPとべん毛運動のアクセルとブレーキ
細菌の生育環境が突然変化すると,菌体にかかる負荷も大きく変化し,遊泳・遊走の速度に直接影響することになる.そのため,こうした外部環境変化に応答するためには,菌体の運動をつかさどるべん毛モーターの回転速度を調節する能力が非常に重要となってくる.車でたとえるならば,ブレーキとアクセルの調節に対応し,たとえば高負荷では,モーターはローターを駆動するためにより大きなトルクを生成する(アクセルを踏む)必要がある.実際,最近の研究では,モーターの回転子(ローター)を囲む固定子(ステーター)ユニットの数を増やすことで,細胞が負荷のアップシフトに応答することが明らかになっており69–72),高負荷では高トルクを生成するために約10個の固定子ユニットが組み込まれており,低負荷では回転子の周りにモーターを回転させるために組み立てられる固定子ユニットは減少する.そのため,固定子ユニット自体が荷重センサーとして機能し,外部の負荷変動に応じて回転数を調整しているようにみえる.環境変化に応じてモーターの回転速度を調整する別の例は,バイオフィルムを形成する際に遊泳菌体が表面に付着し,非運動性固着状態へ細胞の生理状態が変化する際にみられる73).この移行の間,モーターの回転が抑制されることにより,運動性がブロックされ,バイオフィルムの形成が誘発される.c-di-GMPは,このプロセスにおいて重要な役割を担っている.高レベルの細胞内c-di-GMPは,表面付着とバイオフィルム形成を促進する74, 75).
大腸菌においてPDEをコードする遺伝子yhjHを欠損させ,細胞内c-di-GMPの濃度を高めると菌体の運動速度は低下し,逆にDGCをコードする遺伝子を欠損させ,c-di-GMPの濃度を低くすると運動速度が高まるということがわかった20).c-di-GMPによる運動阻害は,PilZドメインを持ちc-di-GMPを結合するタンパク質YcgRの変異体で部分的に抑圧されることから,YcgRはc-di-GMPの濃度に応じて大腸菌べん毛モーターの回転速度を抑えていると考えられていた.そして,YcgRのモーター中の作用部位が報告され,c-di-GMPが結合して活性化したYcgRは,回転子であるFliMとFliGに特異的に結合する76, 77)という報告と,固定子MotAに結合するという報告78)がなされている(図8).YcgRの回転子への結合により反時計回りのモーター回転が増え,同時にブレーキがかかるようにトルク発生が阻害されて,それまで活発だった細菌の運動が低下すると考察されている.YcgRとMotAの細胞質領域の相互作用については,生化学的なアプローチで確かめられている21).YcgRのC末端ドメインがMotAと,N末端ドメインがFliGと相互作用すると推測している.また,細菌は飢餓状態で細胞どうしが細胞表面でくっつき,多細胞で集団を形成することが知られているが,c-di-GMPが高濃度の条件下では表面接着が強くなり,バイオフィルムの形成に有利な状態になることがわかった.さらに,ycgRを欠損させると細胞接着が野生型のレベルにまで低下することから,べん毛の運動を抑制する遺伝子ycgRは,バイオフィルムの形成を促進する働きも担っていると考えられる77).大腸菌はH+駆動型モーターで回転するが,Na+駆動型モーターの場合にも,V. choleraeにおいてYcgRホモログによるモーター回転への影響が報告されている.V. choleraeはPilZドメインを持つタンパク質を複数個持つが,そのうちPlzDがc-di-GMPを結合すること,最少培地で野生株で大量発現させると,運動能が低下することがわかっている79).V. alginolyticusにおいても貧栄養の状態でPlzDを発現させると運動性が大きく阻害されることが報告されている80).V. choleraeにおいて,c-di-GMPが高濃度のときにのみバイオフィルムの形成を促進するPlzCと,c-di-GMPの濃度に関わらずバイオフィルムの形成を促進するPlzBが見つかった.これらの二つのタンパク質はいずれもPilZドメインを持っていることはわかっているが,その運動能やバイオフィルム形成への影響がどのようにして起こるのか,特に上述のH+駆動型モーターと同じような仕組みで回転阻害が起こるのかは明らかではない.
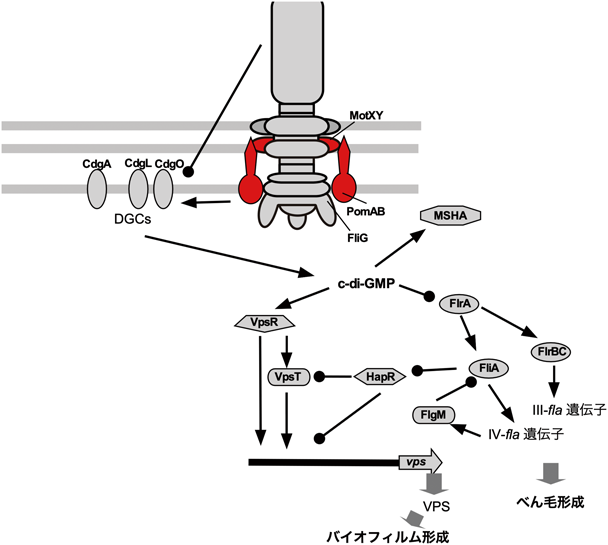
べん毛遺伝子欠損とバイオフィルム形成の関係はこれまでにいろいろ調べられていた.コレラ菌のO139株とエルトール株ではバイオフィルム形成の応答が異なっているが,O139株ではべん毛遺伝子が欠損すると,vps遺伝子の発現に依存したrugoseコロニー(コロニーの表面がしわになる)を作ることが報告されていた81).また,べん毛遺伝子制御系がDGCであるCdgDの発現を制御していることも報告されている58).つまりCdgDを介して,べん毛遺伝子制御系がコレラ菌の病原因子の発現を制御すると考えられる.O139株の主要フラジェリン遺伝子であるflaA遺伝子の欠損によりrugoseコロニーが形成されるが,べん毛固定子関連遺伝子であるpomAやmotX遺伝子欠損を導入することで,smoothコロニーに戻ることが報告されている82).そして,この現象は菌体外多糖合成のvpsR遺伝子発現がpomAやmotX遺伝子欠損で変化した結果であると説明されている.最近,この現象を網羅的に調べて,べん毛遺伝子とバイオフィルム形成の関係が明らかにされた.28個あるDGCに変異を導入して,CdgA, CdgL, CdgOがこの現象に関与していることが明らかにされた52).2回膜貫通タンパク質と予想されているCdgAは単独でコロニー形態に影響を与える強い効果があり,rugoseコロニー形成に関与することはすでに知られていた83).flaA遺伝子が欠損すると細胞内c-di-GMPの濃度が上昇するのだが,このc-di-GMP濃度上昇を担っているのが,これら三つのDGCということになる.c-di-GMPがべん毛遺伝子やvps遺伝子発現を制御しているのだから,非常に複雑な応答となっている(図9).
c-di-GMPによる制御は,役者が多くて理解するのが困難である.なぜ,べん毛形成の変異でc-di-GMPの濃度変化が起こり,バイオフィルム形成が影響を受けるのか,さっぱりわからなかった.しかし,最近の研究で,べん毛遺伝子がc-di-GMPの合成や分解に関与する遺伝子と相互作用して,その活性を制御するという事実が明らかになっている.そして,べん毛の固定子膜タンパク質が膜結合型のc-di-GMP合成分解タンパク質の活性を制御していることも明らかになった.さらに,今年,長年の構造解析が待たれていたエネルギー変換複合体であるべん毛モーター固定子の構造解析も発表され,大きなブレークスルーが期待されている.我々の研究室でも長年の課題であった,ダイナモメーターとして粘性やべん毛回転がどのように遺伝子発現に関与するのかが,明らかになる日も近い気がする.
謝辞Acknowledgments
名古屋大学大学院理学研究科生命理学専攻生体膜機能グループにおいて,多くの共同研究者や学生が細菌べん毛研究に携わってくれた.c-di-GMPに関連した研究は,私のメインであるべん毛モーター研究のサイドビジネスとして行ってきた.c-di-GMPに関連したべん毛の総説を書くことを勧め,執筆を手伝ってくださいました本間道夫教授に感謝申し上げます.
引用文献References
1) Ross, P., Aloni, Y., Weinhouse, C., Michaeli, D., Weinberger-Ohana, P., Meyer, R., & Benziman, M. (1985) An unusual guanyl oligonucleotide regulates cellulose synthesis in Acetobacter xylinum. FEBS Lett., 186, 191–196.
2) Ross, P., Weinhouse, H., Aloni, Y., Michaeli, D., Weinberger-Ohana, P., Mayer, R., Braun, S., de Vroom, E., van der Marel, G.A., van Boom, J.H., et al. (1987) Regulation of cellulose synthesis in Acetobacter xylinum by cyclic diguanylic acid. Nature, 325, 279–281.
3) Weinhouse, H., Sapir, S., Amikam, D., Shilo, Y., Volman, G., Ohana, P., & Benziman, M. (1997) c-di-GMP-binding protein, a new factor regulating cellulose synthesis in Acetobacter xylinum. FEBS Lett., 416, 207–211.
4) Romling, U. & Amikam, D. (2006) Cyclic di-GMP as a second messenger. Curr. Opin. Microbiol., 9, 218–228.
5) Amikam, D. & Galperin, M.Y. (2006) PilZ domain is part of the bacterial c-di-GMP binding protein. Bioinformatics, 22, 3–6.
6) Ryjenkov, D.A., Simm, R., Romling, U., & Gomelsky, M. (2006) The PilZ domain is a receptor for the second messenger c-di-GMP: The PilZ domain protein YcgR controls motility in enterobacteria. J. Biol. Chem., 281, 30310–30314.
7) Benach, J., Swaminathan, S.S., Tamayo, R., Handelman, S.K., Folta-Stogniew, E., Ramos, J.E., Forouhar, F., Neely, H., Seetharaman, J., Camilli, A., et al. (2007) The structural basis of cyclic diguanylate signal transduction by PilZ domains. EMBO J., 26, 5153–5166.
8) Opoku-Temeng, C., Zhou, J., Zheng, Y., Su, J., & Sintim, H.O. (2016) Cyclic dinucleotide (c-di-GMP, c-di-AMP, and cGAMP) signalings have come of age to be inhibited by small molecules. Chem. Commun. (Camb.), 52, 9327–9342.
9) Hengge, R. (2020) Linking bacterial growth, survival, and multicellularity—small signaling molecules as triggers and drivers. Curr. Opin. Microbiol., 55, 57–66.
10) Gancedo, J.M. (2013) Biological roles of cAMP: variations on a theme in the different kingdoms of life. Biol. Rev. Camb. Philos. Soc., 88, 645–668.
11) Ryjenkov, D.A., Tarutina, M., Moskvin, O.V., & Gomelsky, M. (2005) Cyclic diguanylate is a ubiquitous signaling molecule in bacteria: Insights into biochemistry of the GGDEF protein domain. J. Bacteriol., 187, 1792–1798.
12) Paul, R., Weiser, S., Amiot, N.C., Chan, C., Schirmer, T., Giese, B., & Jenal, U. (2004) Cell cycle-dependent dynamic localization of a bacterial response regulator with a novel di-guanylate cyclase output domain. Genes Dev., 18, 715–727.
13) Hecht, G.B. & Newton, A. (1995) Identification of a novel response regulator required for the swarmer-to-stalked-cell transition in Caulobacter crescentus. J. Bacteriol., 177, 6223–6229.
14) Schirmer, T. & Jenal, U. (2009) Structural and mechanistic determinants of c-di-GMP signalling. Nat. Rev. Microbiol., 7, 724–735.
15) Schmidt, A.J., Ryjenkov, D.A., & Gomelsky, M. (2005) The ubiquitous protein domain EAL is a cyclic diguanylate-specific phosphodiesterase: Enzymatically active and inactive EAL domains. J. Bacteriol., 187, 4774–4781.
16) Christen, M., Christen, B., Folcher, M., Schauerte, A., & Jenal, U. (2005) Identification and characterization of a cyclic di-GMP-specific phosphodiesterase and its allosteric control by GTP. J. Biol. Chem., 280, 30829–30837.
17) Tamayo, R., Tischler, A.D., & Camilli, A. (2005) The EAL domain protein VieA is a cyclic diguanylate phosphodiesterase. J. Biol. Chem., 280, 33324–33330.
18) Ryan, R.P., Fouhy, Y., Lucey, J.F., Crossman, L.C., Spiro, S., He, Y.W., Zhang, L.H., Heeb, S., Camara, M., Williams, P., et al. (2006) Cell-cell signaling in Xanthomonas campestris involves an HD-GYP domain protein that functions in cyclic di-GMP turnover. Proc. Natl. Acad. Sci. USA, 103, 6712–6717.
19) Romling, U., Galperin, M.Y., & Gomelsky, M. (2013) Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev., 77, 1–52.
20) Ko, M. & Park, C. (2000) Two novel flagellar components and H-NS are involved in the motor function of Escherichia coli. J. Mol. Biol., 303, 371–382.
21) Hou, Y.J., Yang, W.S., Hong, Y., Zhang, Y., Wang, D.C., & Li, D.F. (2020) Structural insights into the mechanism of c-di-GMP-bound YcgR regulating flagellar motility in Escherichia coli. J. Biol. Chem., 295, 808–821.
22) Nieto, V., Partridge, J.D., Severin, G.B., Lai, R.Z., Waters, C.M., Parkinson, J.S., & Harshey, R.M. (2020) Under elevated c-di-GMP in Escherichia coli, YcgR alters flagellar motor bias and speed sequentially, with additional negative control of the flagellar regulon via the adaptor protein RssB. J. Bacteriol., 202, e00578–e00519.
23) Krasteva, P.V., Fong, J.C., Shikuma, N.J., Beyhan, S., Navarro, M.V., Yildiz, F.H., & Sondermann, H. (2010) Vibrio cholerae VpsT regulates matrix production and motility by directly sensing cyclic di-GMP. Science, 327, 866–868.
24) Matsuyama, B.Y., Krasteva, P.V., Baraquet, C., Harwood, C.S., Sondermann, H., & Navarro, M.V. (2016) Mechanistic insights into c-di-GMP-dependent control of the biofilm regulator FleQ from Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA, 113, E209–E218.
25) Teschler, J.K., Zamorano-Sanchez, D., Utada, A.S., Warner, C.J., Wong, G.C., Linington, R.G., & Yildiz, F.H. (2015) Living in the matrix: assembly and control of Vibrio cholerae biofilms. Nat. Rev. Microbiol., 13, 255–268.
26) Reidl, J. & Klose, K.E. (2002) Vibrio cholerae and cholera: Out of the water and into the host. FEMS Microbiol. Rev., 26, 125–139.
27) Skorupski, K. & Taylor, R.K. (1997) Control of the ToxR virulence regulon in Vibrio cholerae by environmental stimuli. Mol. Microbiol., 25, 1003–1009.
28) Champion, G.A., Neely, M.N., Brennan, M.A., & DiRita, V.J. (1997) A branch in the ToxR regulatory cascade of Vibrio cholerae revealed by characterization of toxT mutant strains. Mol. Microbiol., 23, 323–331.
29) Häse, C.C. & Mekalanos, J.J. (1998) TcpP protein is a positive regulator of virulence gene expression in Vibrio cholerae. Proc. Natl. Acad. Sci. USA, 95, 730–734.
30) Brown, R.C. & Taylor, R.K. (1995) Organization of tcp, acf, and toxT genes within a ToxT-dependent operon. Mol. Microbiol., 16, 425–439.
31) Faruque, S.M., Albert, M.J., & Mekalanos, J.J. (1998) Epidemiology, genetics, and ecology of toxigenic Vibrio cholerae. Microbiol. Mol. Biol. Rev., 62, 1301–1314.
32) Gardel, C.L. & Mekalanos, J.J. (1996) Alterations in Vibrio cholerae motility phenotypes correlate with changes in virulence factor expression. Infect. Immun., 64, 2246–2255.
33) Kustu, S., Santero, E., Keener, J., Popham, D., & Weiss, D. (1989) Expression of sigma54 (ntrA)-dependent genes is probably united by a common mechanism. Microbiol. Rev., 53, 367–376.
34) Klose, K.E. & Mekalanos, J.J. (1998) Distinct roles of an alternative s factor during both free-swimming and colonizing phases of the Vibrio cholerae pathogenic cycle. Mol. Microbiol., 28, 501–520.
35) Bomchil, N., Watnick, P., & Kolter, R. (2003) Identification and characterization of a Vibrio cholerae gene, mbaA, involved in maintenance of biofilm architecture. J. Bacteriol., 185, 1384–1390.
36) Rashid, M.H., Rajanna, C., Ali, A., & Karaolis, D.K. (2003) Identification of genes involved in the switch between the smooth and rugose phenotypes of Vibrio cholerae. FEMS Microbiol. Lett., 227, 113–119.
37) Boles, B.R. & McCarter, L.L. (2002) Vibrio parahaemolyticus scrABC, a novel operon affecting swarming and capsular polysaccharide regulation. J. Bacteriol., 184, 5946–5954.
38) Tischler, A.D. & Camilli, A. (2004) Cyclic diguanylate (c-di-GMP) regulates Vibrio cholerae biofilm formation. Mol. Microbiol., 53, 857–869.
39) Fong, J.C.N., Syed, K.A., Klose, K.E., & Yildiz, F.H. (2010) Role of Vibrio polysaccharide (vps) genes in VPS production, biofilm formation and Vibrio cholerae pathogenesis. Microbiology, 156, 2757–2769.
40) Fong, J.C. & Yildiz, F.H. (2008) Interplay between cyclic AMP-cyclic AMP receptor protein and cyclic di-GMP signaling in Vibrio cholerae biofilm formation. J. Bacteriol., 190, 6646–6659.
41) Pursley, B.R., Maiden, M.M., Hsieh, M.L., Fernandez, N.L., Severin, G.B., & Waters, C.M. (2018) Cyclic di-GMP regulates TfoY in Vibrio cholerae to control motility by both transcriptional and posttranscriptional mechanisms. J. Bacteriol., 200, e00578–e17.
42) Hsieh, M.L., Hinton, D.M., & Waters, C.M. (2018) VpsR & cyclic di-GMP together drive transcription initiation to activate biofilm formation in Vibrio cholerae. Nucleic Acids Res., 46, 8876–8887.
43) Sloup, R.E., Konal, A.E., Severin, G.B., Korir, M.L., Bagdasarian, M.M., Bagdasarian, M., & Waters, C.M. (2017) Cyclic di-GMP and VpsR induce the expression of Type II secretion in Vibrio cholerae. J. Bacteriol., 199, e00106–e00117.
44) Zielke, R.A., Simmons, R.S., Park, B.R., Nonogaki, M., Emerson, S., & Sikora, A.E. (2014) The type II secretion pathway in Vibrio cholerae is characterized by growth phase-dependent expression of exoprotein genes and is positively regulated by sigmaE. Infect. Immun., 82, 2788–2801.
45) Yildiz, F.H. & Schoolnik, G.K. (1999) Vibrio cholerae O1 El Tor: identification of a gene cluster required for the rugose colony type, exopolysaccharide production, chlorine resistance, and biofilm formation. Proc. Natl. Acad. Sci. USA, 96, 4028–4033.
46) Enos-Berlage, J.L. & McCarter, L.L. (2000) Relation of capsular polysaccharide production and colonial cell organization to colony morphology in Vibrio parahaemolyticus. J. Bacteriol., 182, 5513–5520.
47) Yildiz, F.H. & Visick, K.L. (2009) Vibrio biofilms: So much the same yet so different. Trends Microbiol., 17, 109–118.
48) Kawagishi, I., Maekawa, Y., Atsumi, T., Homma, M., & Imae, Y. (1995) Isolation of the polar and lateral flagellum-defective mutants in Vibrio alginolyticus and identification of their flagellar driving energy sources. J. Bacteriol., 177, 5158–5160.
49) Atsumi, T., McCarter, L., & Imae, Y. (1992) Polar and lateral flagellar motors of marine Vibrio are driven by different ion-motive forces. Nature, 355, 182–184.
50) McCarter, L., Hilmen, M., & Silverman, M. (1988) Flagellar dynamometer controls swarmer cell differentiation of V. parahaemolyticus. Cell, 54, 345–351.
51) Kawagishi, I., Imagawa, M., Imae, Y., McCarter, L., & Homma, M. (1996) The sodium-driven polar flagellar motor of marine Vibrio as the mechanosensor that regulates lateral flagellar expression. Mol. Microbiol., 20, 693–699.
52) Wu, D.C., Zamorano-Sanchez, D., Pagliai, F.A., Park, J.H., Floyd, K.A., Lee, C.K., Kitts, G., Rose, C.B., Bilotta, E.M., Wong, G.C.L., et al. (2020) Reciprocal c-di-GMP signaling: Incomplete flagellum biogenesis triggers c-di-GMP signaling pathways that promote biofilm formation. PLoS Genet., 16, e1008703.
53) Trimble, M.J. & McCarter, L.L. (2011) Bis-(3′-5′)-cyclic dimeric GMP-linked quorum sensing controls swarming in Vibrio parahaemolyticus. Proc. Natl. Acad. Sci. USA, 108, 18079–18084.
54) Baker, A.E., Webster, S.S., Diepold, A., Kuchma, S.L., Bordeleau, E., Armitage, J.P., & O’Toole, G.A. (2019) Flagellar stators stimulate c-di-GMP production by Pseudomonas aeruginosa. J. Bacteriol., 201, e00741–e18.
55) Baker, A.E., Diepold, A., Kuchma, S.L., Scott, J.E., Ha, D.G., Orazi, G., Armitage, J.P., & O’Toole, G.A. (2016) PilZ domain protein FlgZ mediates cyclic di-GMP-dependent swarming motility control in Pseudomonas aeruginosa. J. Bacteriol., 198, 1837–1846.
56) Chevance, F.F. & Hughes, K.T. (2008) Coordinating assembly of a bacterial macromolecular machine. Nat. Rev. Microbiol., 6, 455–465.
57) Terashima, H., Kojima, S., & Homma, M. (2008) Flagellar motility in bacteria structure and function of flagellar motor. Int. Rev. Cell Mol. Biol., 270, 39–85.
58) Syed, K.A., Beyhan, S., Correa, N., Queen, J., Liu, J., Peng, F., Satchell, K.J., Yildiz, F., & Klose, K.E. (2009) The Vibrio cholerae flagellar regulatory hierarchy controls expression of virulence factors. J. Bacteriol., 191, 6555–6570.
59) McCarter, L.L. (2004) Dual flagellar systems enable motility under different circumstances. J. Mol. Microbiol. Biotechnol., 7, 18–29.
60) Echazarreta, M.A. & Klose, K.E. (2019) Vibrio flagellar synthesis. Front. Cell. Infect. Microbiol., 9, 131.
61) Prouty, M.G., Correa, N.E., & Klose, K.E. (2001) The novel σ54- and σ28-dependent flagellar gene transcription hierarchy of Vibrio cholerae. Mol. Microbiol., 39, 1595–1609.
62) Correa, N.E., Lauriano, C.M., McGee, R., & Klose, K.E. (2000) Phosphorylation of the flagellar regulatory protein FlrC is necessary for Vibrio cholerae motility and enhanced colonization. Mol. Microbiol., 35, 743–755.
63) Srivastava, D., Hsieh, M.L., Khataokar, A., Neiditch, M.B., & Waters, C.M. (2013) Cyclic di-GMP inhibits Vibrio cholerae motility by repressing induction of transcription and inducing extracellular polysaccharide production. Mol. Microbiol., 90, 1262–1276.
64) Baraquet, C. & Harwood, C.S. (2013) Cyclic diguanosine monophosphate represses bacterial flagella synthesis by interacting with the Walker A motif of the enhancer-binding protein FleQ. Proc. Natl. Acad. Sci. USA, 110, 18478–18483.
65) Kim, Y.K. & McCarter, L.L. (2000) Analysis of the polar flagellar gene system of Vibrio parahaemolyticus. J. Bacteriol., 182, 3693–3704.
66) McCarter, L.L. (2001) Polar flagellar motility of the Vibrionaceae. Microbiol. Mol. Biol. Rev., 65, 445–462.
67) Soutourina, O.A. & Bertin, P.N. (2003) Regulation cascade of flagellar expression in Gram-negative bacteria. FEMS Microbiol. Rev., 27, 505–523.
68) Stewart, B.J. & McCarter, L.L. (2003) Lateral flagellar gene system of Vibrio parahaemolyticus. J. Bacteriol., 185, 4508–4518.
69) Lele, P.P., Hosu, B.G., & Berg, H.C. (2013) Dynamics of mechanosensing in the bacterial flagellar motor. Proc. Natl. Acad. Sci. USA, 110, 11839–11844.
70) Tipping, M.J., Delalez, N.J., Lim, R., Berry, R.M., & Armitage, J.P. (2013) Load-dependent assembly of the bacterial flagellar motor. MBio, 4, e00551–e13.
71) Terahara, N., Noguchi, Y., Nakamura, S., Kami-Ike, N., Ito, M., Namba, K., & Minamino, T. (2017) Load- and polysaccharide-dependent activation of the Na+-type MotPS stator in the Bacillus subtilis flagellar motor. Sci. Rep., 7, 46081.
72) Pourjaberi, S.N.S., Terahara, N., Namba, K., & Minamino, T. (2017) The role of a cytoplasmic loop of MotA in load-dependent assembly and disassembly dynamics of the MotA/B stator complex in the bacterial flagellar motor. Mol. Microbiol., 106, 646–658.
73) Baker, A.E. & O’Toole, G.A. (2017) Bacteria, rev your engines: Stator dynamics regulate flagellar motility. J. Bacteriol., 199, e00088–e17.
74) Jenal, U., Reinders, A., & Lori, C. (2017) Cyclic di-GMP: Second messenger extraordinaire. Nat. Rev. Microbiol., 15, 271–284.
75) Hengge, R. (2009) Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol., 7, 263–273.
76) Paul, K., Nieto, V., Carlquist, W.C., Blair, D.F., & Harshey, R.M. (2010) The c-di-GMP binding protein YcgR controls flagellar motor direction and speed to affect chemotaxis by a “backstop brake” mechanism. Mol. Cell, 38, 128–139.
77) Fang, X. & Gomelsky, M. (2010) A post-translational, c-di-GMP-dependent mechanism regulating flagellar motility. Mol. Microbiol., 76, 1295–1305.
78) Boehm, A., Kaiser, M., Li, H., Spangler, C., Kasper, C.A., Ackermann, M., Kaever, V., Sourjik, V., Roth, V., & Jenal, U. (2010) Second messenger-mediated adjustment of bacterial swimming velocity. Cell, 141, 107–116.
79) Pratt, J.T., Tamayo, R., Tischler, A.D., & Camilli, A. (2007) PilZ domain proteins bind cyclic diguanylate and regulate diverse processes in Vibrio cholerae. J. Biol. Chem., 282, 12860–12870.
80) Kojima, S., Yoneda, T., Morimoto, W., & Homma, M. (2019) Effect of PlzD, a YcgR homologue of c-di-GMP-binding protein, on polar flagellar motility in Vibrio alginolyticus. J. Biochem., 166, 77–88.
81) Watnick, P.I., Lauriano, C.M., Klose, K.E., Croal, L., & Kolter, R. (2001) The absence of a flagellum leads to altered colony morphology, biofilm development and virulence in Vibrio cholerae O139. Mol. Microbiol., 39, 223–235.
82) Lauriano, C.M., Ghosh, C., Correa, N.E., & Klose, K.E. (2004) The sodium-driven flagellar motor controls exopolysaccharide expression in Vibrio cholerae. J. Bacteriol., 186, 4864–4874.
83) Beyhan, S., Bilecen, K., Salama, S.R., Casper-Lindley, C., & Yildiz, F.H. (2007) Regulation of rugosity and biofilm formation in Vibrio cholerae: comparison of VpsT and VpsR regulons and epistasis analysis of vpsT, vpsR, and hapR. J. Bacteriol., 189, 388–402.
著者紹介Author Profile
 小嶋 誠司(こじま せいじ)
小嶋 誠司(こじま せいじ)名古屋大学大学院理学研究科 生命理学専攻教授(国際プログラム担当).博士(理学).
略歴1970年愛知県に生る.94年名古屋大学理学部卒業.99年同大学院理学研究科博士後期課程修了.同年4月米国ユタ大学生物学科博士研究員.2004年3月科学技術振興機構ICORP博士研究員.05年10月名古屋大学理学研究科助手.同研究科助教,講師,准教授を経て20年10月より現職.
研究テーマと抱負細菌べん毛モーターのエネルギー変換および回転制御の仕組みと,べん毛の形成位置・本数制御機構の解明を目指しています.太古の昔から存在し,進化の中で研ぎ澄まされた回転モーターの全貌を明らかにし,生命の基本機能である細胞運動の理解につなげたいと考えています.
ウェブサイトhttp://bunshi4.bio.nagoya-u.ac.jp/~bunshi4/profile/kojima.html
趣味読書・映画と音楽の鑑賞.学生時代はバレーボールをしていましたが最近は運動不足です.