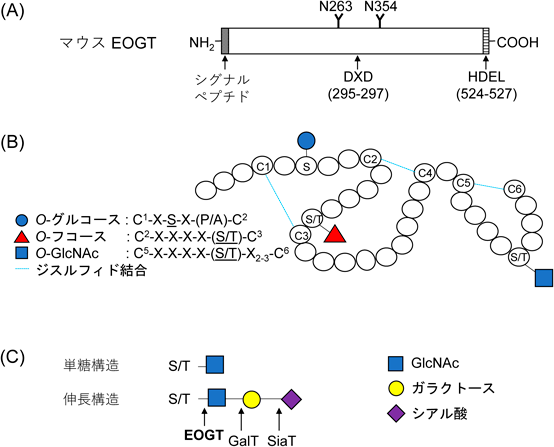
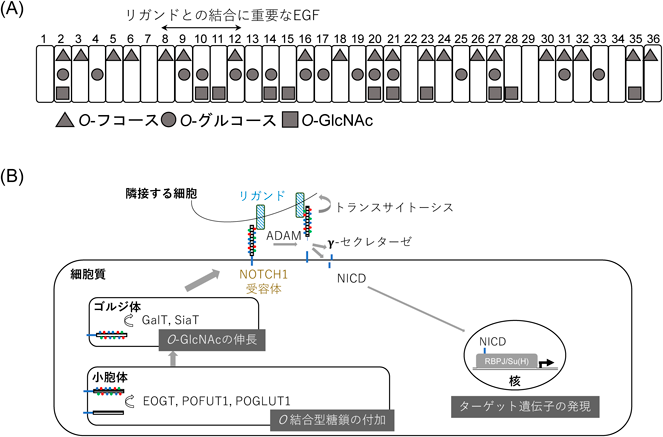
細胞外O-GlcNAcの構造と機能Recent topics on structure and function of extracellular O-GlcNAc
名古屋大学大学院医学系研究科Nagoya University Graduate School of Medicine ◇ 〒466–8550 名古屋市昭和区鶴舞町65 ◇ 65 Tsuruma-cho, Showa-ku, Nagoya, 466–8550 Japan
発行日:2020年12月25日Published: December 25, 2020